Exogenous L-fucose attenuates depression induced by chronic unpredictable stress: Implicating core fucosylation has an antidepressant potential
Dan Wang, Tomohiko Fukuda, Tiangui Wu, Xing Xu, Tomoya Isaji, Jianguo Gu

TL;DR
L-fucose reduces depressive behaviors and neuroinflammation in mice, suggesting core fucosylation may have antidepressant potential.
Contribution
This study reveals that exogenous L-fucose has antidepressant effects by modulating neuroinflammation and synaptic defects.
Findings
L-fucose reduced depressive-like behaviors and pro-inflammatory cytokines in mice with CUS-induced depression.
L-fucose treatment increased dendritic spine density and PSD-95 expression suppressed by CUS.
Antidepressant effects of L-fucose were observed in both Fut8+/− and Fut8+/+ mice.
Abstract
Core fucosylation is one of the most essential modifications of the N-glycans, catalyzed by α1,6-fucosyltransferase (Fut8), which transfers fucose from guanosine 5′-diphosphate (GDP)-fucose to the innermost N-acetylglucosamine residue of N-glycans in an α1-6 linkage. Our previous studies demonstrated that lipopolysaccharide (LPS) can induce a more robust neuroinflammatory response in Fut8 homozygous knockout (KO) (Fut8−/−) and heterozygous KO (Fut8+/−) mice contrasted to the wild-type (Fut8+/+) mice. Exogenous administration of L-fucose suppressed LPS-induced neuroinflammation. Numerous studies indicate that neuroinflammation plays a vital role in the development of depression. Here, we investigated whether core fucosylation regulates depression induced by chronic unpredictable stress (CUS), a well-established model for depression. Our results showed that Fut8+/− mice exhibited…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
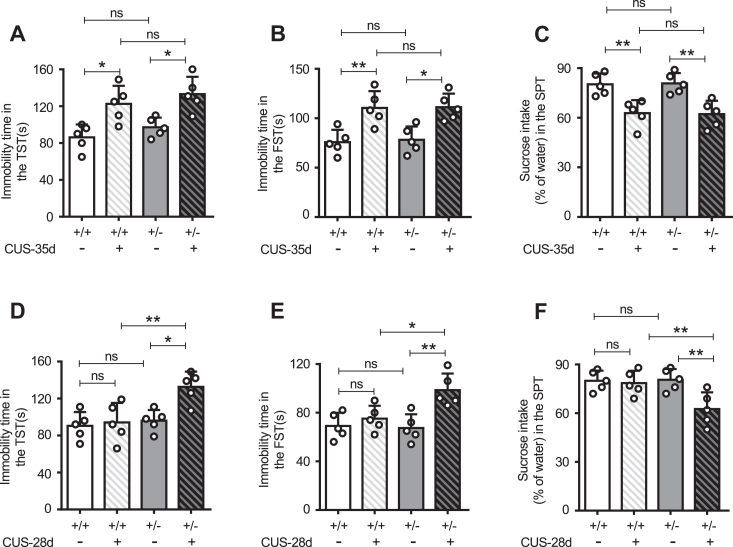 Figure 1
Figure 1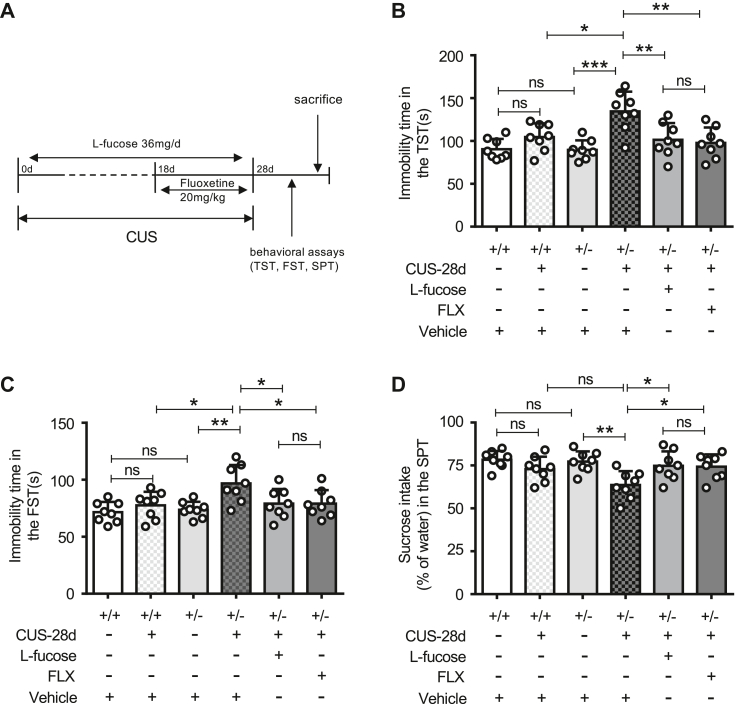 Figure 2
Figure 2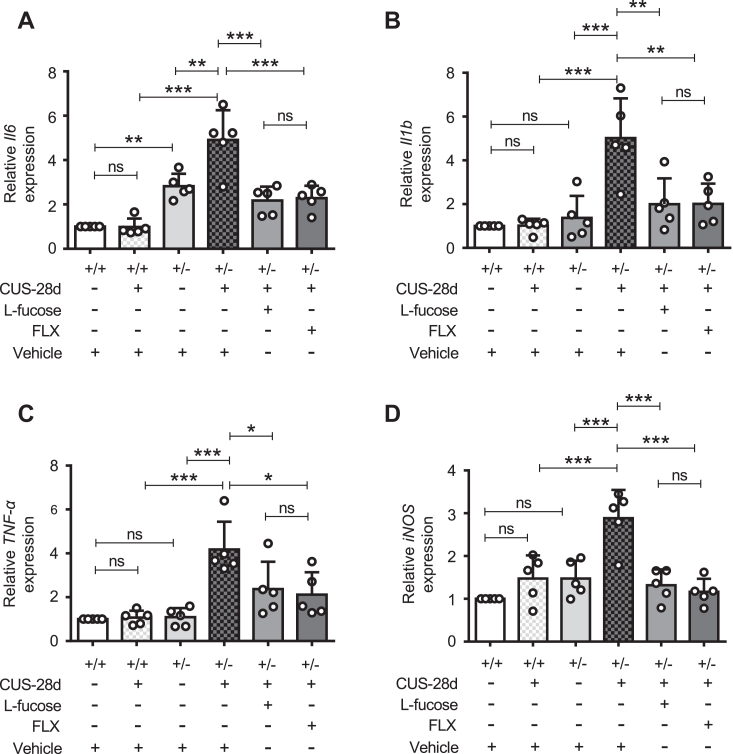 Figure 3
Figure 3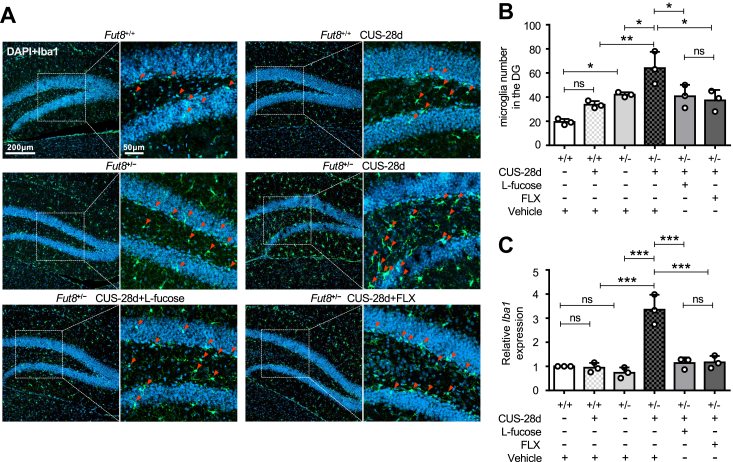 Figure 4
Figure 4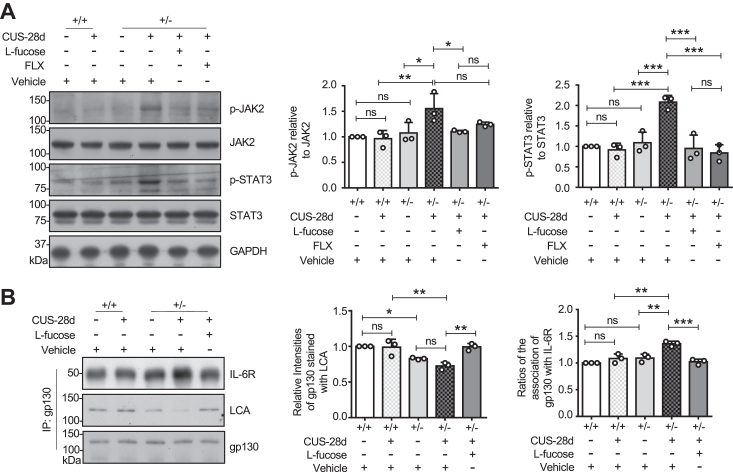 Figure 5
Figure 5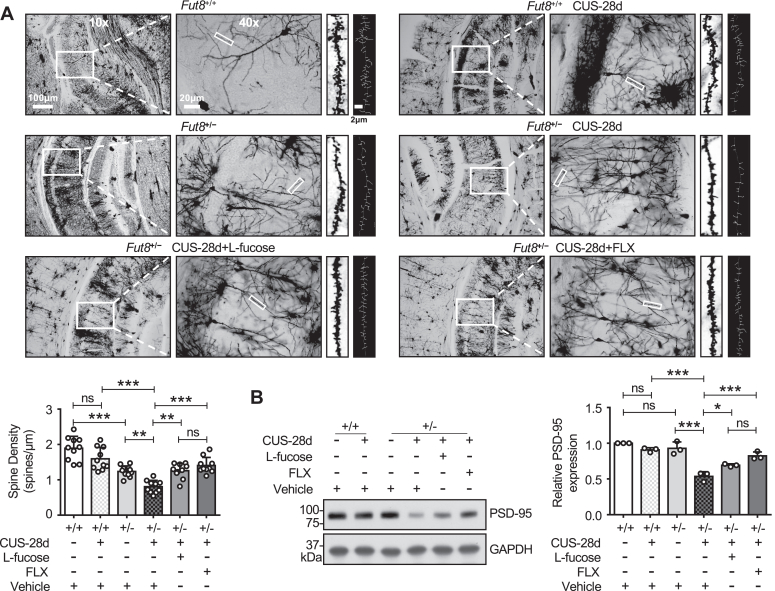 Figure 6
Figure 6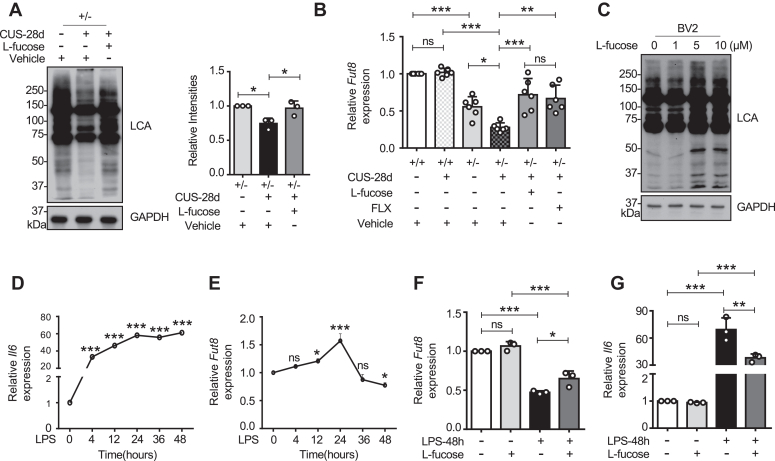 Figure 7
Figure 7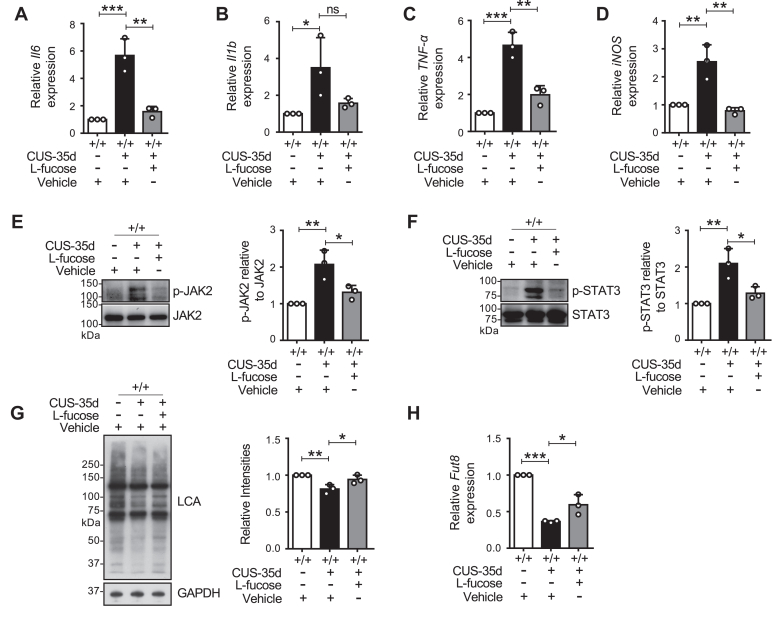 Figure 8
Figure 8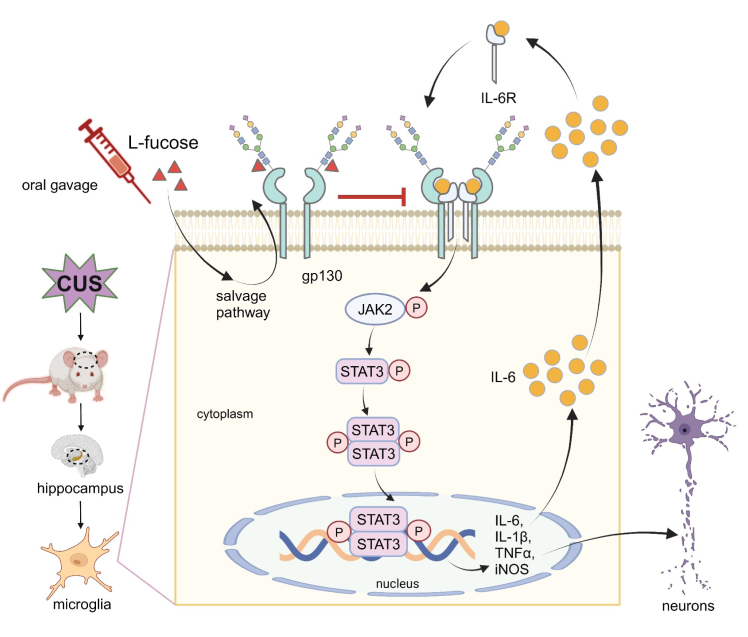 Figure 9
Figure 9Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsTryptophan and brain disorders · Stress Responses and Cortisol · Neuroinflammation and Neurodegeneration Mechanisms
Major depressive disorder (MDD) is a common mental disorder characterized by persistent symptoms of depressed mood and anhedonia (1, 2). MDD heavily impacts patients' sleep quality, appetite, cognitive flexibility, and mobility (3, 4, 5). In severe cases, individuals with depression may exhibit suicidal tendencies (6). Approximately 300 million people worldwide suffer from MDD, and its prevalence continues to rise (7). The World Health Organization listed MDD as the third largest disease burden globally in 2018, with projections indicating it will rank first by 2030 (8).
Numerous studies have shown that MDD is closely related to elevated levels of neuroinflammatory biomarkers, such as interleukin (IL)-6 (9, 10, 11), IL-1β (12, 13), and tumor necrosis factor-alpha (TNF-α) (14, 15, 16). Inflammation has even been proposed as a biomarker in the treatment of depression (17). Rodent models of chronic stress also exhibit increased levels of pro-inflammatory cytokines (18, 19, 20). Depression is increasingly recognized as a microglial disease (21). As immune effector cells in the central nervous system (CNS), microglia play a crucial role in neuroprotection. In depression and its animal models, microglia become overactivated, accelerating the neuroinflammatory response by secreting large amounts of pro-inflammatory (22, 23, 24) cytokines. Excessive inflammation can lead to neuronal apoptosis and cell death (25, 26).
L-fucose is a dietary deoxyhexose sugar that is taken up and processed in cells, converted into guanosine 5′-diphosphate (GDP)-fucose, which is utilized in the endoplasmic reticulum or Golgi by 13 fucosyltransferases to synthesize fucosylated glycans (27, 28, 29). GDP-fucose can be synthesized in two ways: the de novo pathway and the salvage pathway, and the de novo pathway is dominant (30, 31). Dietary intake or lysosomal breakdown of glycoproteins can provide L-fucose in the salvage pathway. When the de novo pathway is compromised, the salvage pathway supplements GDP-fucose to maintain normal physiological functions (32). L-fucose has shown anti-tumor and anti-inflammatory effects in various diseases. For example, in melanoma models, dietary L-fucose enhanced immunotherapy response; it has also been shown to reduce gut inflammation and mitigate high-salt diet-induced inflammation (33, 34, 35). Ample evidence suggests that L-fucose, the predominant bioactive component of fucoidan, possesses neuroprotective and neurogenic effects in the CNS (36, 37, 38). However, the underlying molecular mechanisms for these effects remain unclear.
Core fucosylation is a process catalyzed by α1,6-fucosyltransferase (Fut8), which transfers a fucose residue from GDP-fucose to the innermost N-acetylglucosamine residue of N-linked glycans in complex and hybrid glycoproteins (39, 40). Patients with FUT8-congenital disorders of glycosylation, characterized by impaired core fucosylation, often present with epilepsy, short stature, microcephaly, and intellectual disabilities (41). Our previous studies demonstrated that Fut8 homozygous knockout (KO) (Fut8^−/−^) mice exhibit memory impairments and abnormal behaviors consistent with schizophrenia-like phenotypes (42), along with decreased hippocampal long-term potentiation (LTP) compared to the wild-type (Fut8^+/+^) mice (43). Additionally, Fut8 deficiency via shRNA has been shown to impair memory, learning, and neurogenesis in adult mice (44). These findings suggest that a lack of core fucosylation may contribute to developing CNS disorders. Both Fut8^−/−^ mice and Fut8^+/−^ mice display heightened microglial activation and a more severe inflammatory response to lipopolysaccharide (LPS) stimulation than Fut8^+/+^ mice (45, 46), indicating that neuroinflammation may be negatively regulated by core fucosylation. Lack of the Fut8 gene significantly enhances the complex formation between the IL-6 receptor (IL-6R) and glycoprotein 130 (gp130), resulting in stronger neuroinflammation. Conversely, L-fucose administration increases core fucosylation of gp130, thereby reducing the complex formation and exerting anti-neuroinflammation effects (45). Given the close relationship between neuroinflammation and CNS disorders, we hypothesize that core fucosylation may also influence the onset and progression of depression. In this study, we investigated the effects of core fucosylation on depression induced by chronic unpredictable stress (CUS), one of the most established models for depression (47). Our findings indicate that Fut8^+/−^ mice develop depressive-like behaviors earlier than Fut8^+/+^ mice, which can be rescued by L-fucose administration. Furthermore, L-fucose ameliorated neuroinflammation and synaptic defects in CUS-induced depression mice. Therefore, L-fucose may hold potential as a therapeutic agent for CNS diseases.
Results
Fut8+/− mice exhibited depression-like behavior earlier than Fut8+/+ mice
Our previous studies revealed that core fucosylation negatively regulates the neuroinflammation induced by LPS; specifically, lower core fucosylation can induce severe neuroinflammation (45, 46). Given the established link between neuroinflammation and depression (12, 15), we explored whether core fucosylation affects the onset and progression of depression. Stress is a significant risk factor for several psychiatric illnesses, including MDD, and is often a trigger for depressive episodes (48, 49, 50). The CUS model of depression is a valid, reliable, and widely used model (47), and the method for simulating the stress variations is quite similar to what humans are exposed to in their daily lives (51, 52). Following 7, 14, 21, 28, and 35 days of exposure to the CUS model, we tested the depressive behavior of Fut8^+/+^ and Fut8^+/−^ mice. At 35 days, compared to each control mice without CUS, both Fut8^+/+^ and Fut8^+/−^ CUS mice exhibited increased immobility times in the tail suspension test (TST) (Fig. 1A) and forced swimming test (FST) (Fig. 1B), as well as decreased sucrose intake in the sucrose preference test (SPT) (Fig. 1C), indicating the success of the CUS model. Notably, in the CUS-28-days group, Fut8^+/−^ mice showed significantly higher immobility time in the TST (Fig. 1D) and FST (Fig. 1E) than controls without CUS. In addition, Fut8^+/−^ mice exhibited significant attenuation in sucrose preference at 28 days of CUS compared to controls in the SPT (Fig. 1F). However, these differences were not observed in Fut8^+/+^ mice at 28 days. All mice did not display depressive-like behaviors after 7, 14, and 21 days of CUS (Fig. S1, A–I). These findings demonstrate that the mice with lower core fucosylation are more susceptible to exhibiting depressive-like behaviors.Figure 1Comparison of depression-like behaviors upon CUS treatment between Fut8^+/−^and Fut8^+/+^mice, Chronic unpredictable stress (CUS) models were established in Fut8^+/+^and Fut8^+/−^mice, grouped at 7, 14, 21, 28, and 35 days post-CUS. Behavioral assessments were conducted using the tail suspension test (TST) (A), forced swimming test (FST) (B), and sucrose preference test (SPT) (C) after 35 days of CUS. Data for TST (D), FST (E), and SPT (F) at 28 days of CUS are also presented. Data were analyzed by one-way ANOVA with Tukey's multiple comparison tests and shown as mean ± SD. n = 5 mice per group. n.s. p > 0.05; ∗p < 0.05; ∗∗p < 0.01.
L-fucose administration abolished increased depressive-like behaviors induced by CUS in Fut8+/− mice
Previous studies found that L-fucose has an inhibitory effect on LPS-induced neuroinflammation (45). Thus, we investigated whether exogenous L-fucose could affect the depressive-like behavior of Fut8^+/−^ mice subjected to CUS. The role of serotonin in CNS immunity is still unclear, but serotonin is indeed linked to inflammation (53, 54). The treatment of MDD patients with selective serotonin reuptake inhibitors (SSRIs) reduces IL-6 and TNF-α levels, and their antidepressant therapeutic effects may be partly attributed to their anti-inflammatory properties (55). The classic antidepressant FLX is an SSRI medication and is known to exert significant anti-neuroinflammatory effects (56), which is achieved by inhibiting microglia activation and reducing the release of inflammatory factors (57, 58, 59). FLX is often served as a positive control in studies of depression models (60, 61). Here, we also used FLX as a positive control to evaluate the degree of antidepressant effect of L-fucose since FLX is one of the most widely used drugs in clinical practice and has significant anti-inflammatory effects. As depicted in the schematic (Fig. 2A), L-fucose was administered intragastrically at a dose of 36 mg/day throughout the CUS model (45), while FLX was administered intraperitoneally at 20 mg/kg (62) during the last 10 days. Behavioral tests revealed that L-fucose eliminated the depressive-like behaviors in Fut8^+/−^ mice following CUS, exhibiting similar effects to FLX (Fig. 2). Exogenous L-fucose significantly decreased the immobility time in TST (Fig. 2B) and FST (Fig. 2C) and increased sucrose intake in the SPT (Fig. 2D). These results suggest that L-fucose may possess antidepressant effects similar to FLX.Figure 2L-fucose administration abolished depressive-like behaviors induced by CUS in Fut8^+/−^**mice.**A, schedule of CUS modeling, with Fut8^+/−^ mice administered 36 mg/day of L-fucose or 20 mg/kg of fluoxetine (FLX) during the last 10 days. Behavioral tests were performed, and brain tissues were extracted afterward. B, the immobility time of mice in the TST. C, the immobility time of mice in the FST. D, the sucrose consumption of mice in the SPT. Data was analyzed by one-way ANOVA with Tukey's multiple comparison tests and shown as mean ± SD. n = 8 mice per group. n.s. p > 0.05; ∗p < 0.05; ∗∗p < 0.01; ∗∗∗p < 0.001.
L-fucose suppression increased neuroinflammation induced by CUS in Fut8+/− mice
Neuroinflammation is implicated in MDD through various neurobiological mechanisms (7). Evidence indicates that CUS stress induces the release of pro-inflammatory cytokines, such as IL-6, IL-1β, TNF-α, and inducible NO synthase (iNOS) (18, 19, 20). The structure and function of the hippocampus have been linked to the pathophysiology and progression of MDD (63, 64, 65). Thus, we focused on the hippocampus to evaluate the effect of L-fucose on CUS-induced neuroinflammation. After 28 days of CUS, real-time PCR analysis revealed that the mRNA levels of the Il6 (Fig. 3A), Il1b (Fig. 3B), TNF-α (Fig. 3C), and iNOS (Fig. 3D) were increased significantly in Fut8^+/−^ mice, but not in Fut8^+/+^ mice. Moreover, Fut8^+/−^ mice showed higher levels of Il1b mRNA than Fut8^+/+^ mice in the absence of CUS exposure. Treatment with exogenous L-fucose and FLX both obviously reversed the increased neuroinflammation in Fut8^+/−^ mice after CUS exposure. These findings suggest that L-fucose may reduce depressive-like behavior by mitigating neuroinflammation.Figure 3L-fucose suppressed neuroinflammation induced by CUS in Fut8^+/−^mice. The mRNA levels of neuroinflammatory factors were analyzed by real-time PCR, including interleukin-6 (Il6) (A), interleukin 1 beta (Il1b) (B), tumor necrosis factor-alpha (TNF-α) (C), and inducible nitric oxide synthase (iNOS) (D) in the hippocampus. Glyceraldehyde-3-phosphate dehydrogenase (Gapdh) was used as an internal control. Values were normalized to that of the Gapdh. The value for Fut8^+/+^ mice with vehicle treatment was set as 1.0. The one-way ANOVA test with Tukey's multiple comparison test was used to calculate the quantitative data, which were displayed as the mean ± SD of five independent experiments. n = 5 mice per group. n.s. p > 0.05; ∗p < 0.05; ∗∗p < 0.01; ∗∗∗p < 0.001.
L-fucose inhibited CUS-induced microglial activation in Fut8+/− mice
Microglia are innate immune cells of the CNS, and the alterations of number and function in microglia are closely related to the onset of depression (66, 67). It is widely known that chronic stress can induce the activation of microglia in specific brain regions and alter their phenotypic and functional properties (68). We performed immunofluorescence staining to detect the level of ionized calcium-binding adaptor molecule 1 (Iba1), a microglial activation marker in the dentate gyrus (DG) of the hippocampus (63, 69, 70, 71). After 28 days of CUS, the number of microglia in Fut8^+/−^ mice was significantly increased, which was suppressed after treatment with L-fucose and FLX (Fig. 4, A and B). Real-time PCR analysis also showed that the mRNA level of Iba1 was significantly elevated in Fut8^+/−^ mice exposed to CUS, but not in Fut8^+/+^ mice. In addition, L-fucose or FLX exerted an effective suppression on this increase (Fig. 4C). Without CUS exposure, Fut8^+/−^ mice had more Iba1-positive cells in immunofluorescence results than Fut8^+/+^ mice (Fig. 4B); although there was no significant difference in the mRNA levels of Iba1 between the two mice (Fig. 4C). These results further support the notion that core fucosylation negatively regulates microglia activation and consequently suppresses neuroinflammation.Figure 4**L-fucose inhibited CUS-induced microglial activation.**A, representative immunofluorescence staining images of the dentate gyrus (DG) in hippocampal coronal sections, showing Iba1 as a microglia marker. DAPI was used for nuclear staining. Green fluorescent cells co-localized with DAPI (blue) are identified as Iba1-positive microglia. Arrows indicate Iba1-positive cells. B, quantification of Iba1-positive microglia in the DG region using ImageJ software. The one-way ANOVA test with Tukey's multiple comparison test was used to calculate the quantitative data. Data from three independent experiments are presented as mean ± SD. n = 3 mice per group. n.s. p > 0.05; ∗p < 0.05; ∗∗p < 0.01. C, real-time PCR analysis of Iba1 mRNA expression, normalized to Gapdh (internal control), and Fut8^+/+^ mice with vehicle treatment were set as 1.0. All data were analyzed by one-way ANOVA test with Tukey's multiple comparison test and displayed as the mean ± SD from three independent experiments. n = 3 mice per group. n.s. p > 0.05; ∗∗∗p < 0.001.
L-fucose attenuated the JAK2/STAT3 signaling pathway
The dysregulation of the JAK/STAT (Janus kinase-signal transducer and activator of transcription) signaling pathway plays a significant role in neuroinflammatory diseases (72, 73, 74). STAT3, a downstream effector of IL-6, is involved in the pro-inflammatory activation of microglia and astrocytes in various neuroinflammatory contexts (75, 76, 77). The inhibition of the JAK2/STAT3 pathway has been shown to have anti-neuroinflammatory and neuroprotective effects (78, 79). Meanwhile, FLX attenuated the neuroinflammation by suppressing the activation of JAK/STAT3 signaling (80). Western blotting has been used to assess the phosphorylation levels of JAK2 and STAT3. Quantitative analysis revealed that the phosphorylation levels of JAK2 and STAT3 were significantly increased in Fut8^+/−^ mice after 28 days of CUS, but not in Fut8^+/+^ mice. L-fucose could inhibit the elevated phosphorylation levels of JAK2 and STAT3 in Fut8^+/−^ mice subjected to CUS. FLX also displayed a similar inhibitory trend regarding the phosphorylation levels of JAK2 and STAT3, although no significant differences were observed for p-JAK2 (Fig. 5A). IL-6 signaling includes the gp130 receptor and the JAK/STAT pathway (81). We used co-immunoprecipitation to explore the molecular mechanisms for the effect of exogenous L-fucose administration in the CUS model. According to immunoprecipitation results, the interaction between gp130 and IL-6R was increased upon exposure to CUS in Fut8^+/−^ mice, compared to without CUS. Concomitantly, gp130 in Fut8^+/−^ mice exposed to CUS contained less core fucosylation than in Fut8^+/+^ mice or Fut8^+/−^ mice treated with L-fucose (Fig. 5B). Moreover, exogenous L-fucose administration inhibited the association between gp130 and IL-6R (Fig. 5B), which is similar to the observation in LPS-induced neuroinflammation (45). We further investigated the effects of WP1066, a JAK2/STAT3 inhibitor (82), on neuroinflammation and depression-like behaviors induced by CUS. After the first injection on the fourth day of the CUS model, the intraperitoneal injection was carried out once every 3 days at 30 mg/kg for nine injections, based on dose usage in the previous study (83). The effects of WP1066 on the behavioral experiments of depression were quite similar to those of L-fucose (Fig. S2, A–C). The combined treatment of WP1066 and L-fucose did not observe additive effects. Furthermore, the impact of WP1066 on the expression of inflammatory factors and Fut8 in CUS-induced depression also showed a similar tendency with L-fucose (Fig. S3, A–E). However, WP1066 did not significantly suppress Il1b and iNOS expression. The combined treatment of WP1066 and L-fucose did not observe additive effects, except for the phosphorylated JAK2 levels (Fig. S3F). These results further suggest the suppression of CUS-induced depression by exogenous L-fucose via neutralizing neuroinflammation. These results indicate that L-fucose exerts its anti-inflammatory effects by inhibiting the JAK2/STAT3 pathway, thereby playing an antidepressant role.Figure 5**L-fucose attenuated the JAK2/STAT3 signaling pathway.**A, Western blot analysis of hippocampal tissues after 28 days of CUS, using antibodies against JAK2, phosphor-JAK2 (p-JAK2), STAT3, and phosphor-STAT3 (p-STAT3). GAPDH was used as a loading control. Data was quantified by ImageJ software. The ratio of p-JAK2 to JAK2 for Fut8^+/+^ mice treated with vehicle was set as 1.0. The ratio of p-STAT3 to STAT3 for Fut8^+/+^ mice treated with vehicle was set as 1.0. All data were analyzed by one-way ANOVA test with Tukey's multiple comparison test and displayed as the mean ± SD from three independent experiments. n = 3 mice per group. n.s. p > 0.05; ∗p < 0.05; ∗∗p < 0.01, ∗∗∗p < 0.001. B, equal amounts of hippocampus lysates were immunoprecipitated using an anti-gp130 antibody following a 28-days exposure to CUS. The immunoprecipitates were analyzed by western blotting with anti-IL-6R and anti-gp130 antibodies or lectin blotting with LCA lectin. ImageJ software was used to quantify the data. The ratio of LCA or IL-6R versus gp130 of Fut8^+/+^ mice treated with the vehicle was 1.0. All Data were shown as the mean ± SD from three independent experiments. n = 3 mice per group. n.s. p > 0.05; ∗p < 0.05; ∗∗p < 0.01, ∗∗∗p < 0.001. (One-way ANOVA test with Tukey's multiple comparison test).
Exogenous L-fucose reversed CUS-induced damage in synaptic plasticity
Through the activation of microglia, inflammatory cytokine overproduction causes synaptic dysfunction and neuronal damage in the hippocampus, which in turn induces depressive behaviors (24). Spines possess highly dynamic, and the alterations in their density and morphology are the basis of structural synaptic plasticity (84). Neuroinflammation following depression may lead to impaired synaptic plasticity, decreased dendritic spine density, and synaptic plasticity-related proteins, such as postsynaptic density protein 95 (PSD-95) (47, 85). FLX has the effect of improving synaptic plasticity (86, 87). Given the observed behavioral alterations and cytokine levels in Fut8^+/−^ mice, we hypothesized that neuroinflammation triggered from excessive microglia activation may affect neuronal plasticity. To explore the role of L-fucose in synaptic plasticity, we need to detect the spine density through Golgi staining. Decreased dendritic spine density in the CA1 hippocampus region is associated with depressive-like behavior following CUS (88). Therefore, we examined the synaptic density in the CA1 region of the mouse hippocampus. The results demonstrated that synaptic density in the hippocampus of Fut8^+/−^ mice was lower than that in Fut8^+/+^ mice. There was no significant change in synaptic density in Fut8^+/+^ mice after 28 days of CUS (Fig. 6A). After 28 days of CUS, synaptic densities in Fut8^+/−^ mice were significantly decreased, whereas L-fucose and FLX significantly alleviated this impairment (Fig. 6A). Additionally, the level of synaptic plasticity-related proteins was detected by western blotting. We found the expression level of PSD-95 was significantly suppressed in the hippocampus of Fut8^+/−^ mice exposed to the CUS, compared to the control without CUS (Fig. 6B). Importantly, L-fucose or FLX partially rescued the decreased PSD-95 expression in Fut8^+/−^ CUS mice (Fig. 6B). These data suggest that L-fucose may exert antidepressant effects by restoring synaptic plasticity.Figure 6**Exogenous L-fucose reversed CUS-induced damage in synaptic plasticity.**A, representative images of the dendritic spine in Golgi staining. Images of the hippocampal region under 10x and 40x magnifications, as well as representative synaptic spine images and ImageJ-processed images, are shown from left to right. Spine density is the number of spines divided by the dendritic shaft length. ImageJ software measured the number of branches and total dendritic length. Data from 10 dendritic segments across three mice is mean ± SD. n.s. p > 0.05; ∗∗p < 0.01; ∗∗∗p < 0.001 (one-way ANOVA test with Tukey's multiple comparison test). B, Western blotting analysis of PSD-95 levels in hippocampal tissues after 28 days of CUS, quantified using ImageJ. GAPDH was used as a loading control, and the ratio of PSD-95 to GAPDH for Fut8^+/+^ mice treated with vehicle was set as 1.0. All data were analyzed by one-way ANOVA test with Tukey's multiple comparison test and displayed as the mean ± SD from three independent experiments. n = 3 mice per group. n.s. p > 0.05; ∗p < 0.05; ∗∗∗p < 0.001.
L-fucose recovered the CUS-induced down-regulation of Fut8 expression
To investigate the alteration in core fucosylation after CUS exposure, we checked the level of core fucosylation via lectin blotting using the Lens Culinaris Agglutinin (LCA). LCA can specifically recognize core fucosylation (89). After 28 days of CUS, Fut8^+/−^ mice exhibited a significant reduction in core fucosylation levels, which was reversed by exogenous L-fucose (Fig. 7A). Since L-fucose administration may also affect other fucosylation (90), and studies have found that mice lacking α1,3-fucosyltransferase IX exhibit increased anxiety-like behaviors (91), we also examined the expression levels of Lewis X by western blotting with anti-Lewis X antibody. After CUS exposure, curiously, both Fut8^+/+^ and Fut8^+/−^ mice exhibited decreased Lewis X expression levels, which could not be rescued by L-fucose treatment (Fig. S4, A and B), suggesting the effect of L-fucose on CUS is not dependent on expression levels of Lewis antigen. Given the notable increase in inflammatory factors in Fut8^+/−^ CUS mice, we hypothesized that inflammation might downregulate Fut8 gene expression. Real-time PCR analysis of Fut8 mRNA levels showed a significant decrease in Fut8 expression due to CUS-induced inflammation in Fut8^+/−^ mice. However, this downregulation was reversed in Fut8^+/−^ CUS mice treated with L-fucose or FLX (Fig. 7B). These findings suggest that neuroinflammation may downregulate Fut8 expression. To investigate if the suppression of inflammation or cell autonomy is responsible for the upregulation of Fut8 mRNA following L-fucose treatment, we established the neuroinflammation model by treating BV2 cells cultured in vitro with LPS (1 μg/ml) (92). The results of LCA lectin blotting demonstrated that the core fucosylation levels were increased by L-fucose. The dose of 5 μM L-fucose was adequate for the increasing effect (Fig. 7C). After LPS treatment, Il6 mRNA levels increased significantly (Fig. 7D), and Fut8 mRNA levels increased after 12 h and 24 h of LPS treatment. Interestingly, as the LPS treatment time was postponed, Fut8 mRNA levels decreased (Fig. 7E), suggesting bi-phase responses in the induction. BV2 cells were pretreated with L-fucose for 4 h and then stimulated with LPS for 48 h. The real-time PCR analysis showed decreased Fut8 expression while increasing Il6 expression, significantly rescued by L-fucose (Fig. 7, F and G). Curiously, L-fucose alone did not change Fut8 expression (Fig. 7F).Figure 7**L-fucose rescued the CUS-induced down-regulation of Fut8 expression.**A, Lectin blot analysis of hippocampal tissues after 28 days of CUS, using Lens Culinaris Agglutinin (LCA), specifically recognizing Core fucosylation. GAPDH was used as a loading control. The ratio of LCA versus GAPDH of Fut8^+/−^ mice with the vehicle was set as 1.0. All data were analyzed by one-way ANOVA test with Tukey's multiple comparison test and displayed as the mean ± SD from three independent experiments. ∗p < 0.05. B, real-time PCR analysis of Fut8 mRNA expression. Gapdh was used as an internal control. Each value was normalized to that of the Gapdh. The values of Fut8 versus Gapdh in Fut8^+/+^ mice treated with vehicle were set to 1.0. All data were analyzed by one-way ANOVA test with Tukey's multiple comparison test and displayed as the mean ± SD from six independent experiments. n = 6 mice per group. n.s. p > 0.05; ∗p < 0.05; ∗∗p < 0.01; ∗∗∗p < 0.001. C, BV2 cells were treated with L-fucose at the indicated concentrations and cultured for 48 h. The same amounts of cell lysates were lectin blotted with LCA. GAPDH served as a loading control. D and E, BV2 cells were treated with LPS at 1 μg/ml for the indicated times. Real-time PCR was used to analyze the mRNA levels of Il6 and Fut8. Gapdh was used as an internal control. Each value was normalized to that of the Gapdh. Il6 or Fut8 versus Gapdh values in BV2 cells without LPS treatment were set to 1.0. All data were analyzed by one-way ANOVA with Tukey's multiple comparison test and shown as the mean ± SD from three independent experiments. n.s. p > 0.05; ∗p < 0.05; ∗∗∗p < 0.001. F and G, after pretreatment with or without L-fucose at 5 μM for 4 h, the cells were stimulated with LPS for 48 h. Real-time PCR was used to analyze the mRNA levels of Il6 and Fut8. Gapdh was used as an internal control. Each value was normalized to that of the Gapdh. Il6 or Fut8 versus Gapdh values in BV2 cells without LPS and L-fucose treatment were set to 1.0. All data were analyzed by one-way ANOVA with Tukey's multiple comparison test and displayed as the mean ± SD from three independent experiments. n.s. p > 0.05; ∗p < 0.05; ∗∗p < 0.01; ∗∗∗p < 0.001.
Effects of L-fucose on Fut8+/+ mice exposed to CUS
We examined the effects of L-fucose on Fut8^+/+^ mice after CUS exposure to further explore the role of L-fucose in Fut8^+/+^ mice. Following 35 days of CUS, Fut8^+/+^ mice displayed increased immobility time in TST (Fig. S5A) and FST (Fig. S5B), alongside reduced sucrose preference in the SPT (Fig. S5C) compared to the controls without CUS. Importantly, these alterations were ameliorated by L-fucose treatment. Additionally, the real-time PCR analysis showed that the elevated mRNA levels of Il6 (Fig. 8A), Il1b (Fig. 8B), TNF-α (Fig. 8C), and iNOS (Fig. 8D) in Fut8^+/+^ mice following CUS were significantly suppressed by exogenous L-fucose. The levels of phospho-JAK2 (Fig. 8E) and phospho-STAT3 (Fig. 8F) were also significantly increased in Fut8^+/+^ mice after CUS, which were inhibited by L-fucose treatment. Measurements of core fucosylation using LCA revealed that CUS exposure resulted in a significant decrease in LCA reactivity, which was partially rescued by L-fucose (Fig. 8G). Notably, Fut8 mRNA expression levels were downregulated in Fut8^+/+^ mice after CUS (Fig. 8H), and this downregulation could also be partially reversed by L-fucose treatment. These results strongly indicate that core fucosylation is crucial for CNS maintenance, and L-fucose treatment can mitigate depressive-like behaviors in Fut8^+/+^ mice.Figure 8Effects of L-fucose on Fut8^+/+^**mice exposed to CUS at 35 days.**Fut8^+/+^ mice were subjected to the CUS for 35 days and treated with L-fucose. A–D, the mRNA levels of cytokines were analyzed by real-time PCR, including Il6 (A), Il1b (B), TNF-α (C), and iNOS (D) in the hippocampus. The Gapdh was used to normalize each value. The value of Il6, Il1b, TNF-α or iNOS versus Gapdh in Fut8^+/+^ mice treated with the vehicle was set as 1.0. All data were analyzed by one-way ANOVA test with Tukey's multiple comparison test and displayed as the mean ± SD from three independent experiments. n = 3 mice per group. n.s. p > 0.05; ∗p < 0.05; ∗∗p < 0.01; ∗∗∗p < 0.001. E and F, Western blotting analyses for p-JAK2 (E) and p-STAT3 (F) were conducted on hippocampal tissues, with data quantified using ImageJ software. The ratios of p-JAK2 to JAK2 and p-STAT3 to STAT3 for Fut8^+/+^ vehicle-treated mice were set as 1.0. GAPDH was used as a loading control. All data were analyzed by one-way ANOVA test with Tukey's multiple comparison test and displayed as the mean ± SD from three independent experiments. n = 3 mice per group. ∗p < 0.05; ∗∗p < 0.01. G, lectin blot analysis with LCA, normalized to GAPDH, with the Fut8^+/+^ vehicle-treated mice set as 1.0. Data were shown as mean ± SD from three independent experiments. n = 3 mice per group. ∗p < 0.05; ∗∗p < 0.01. (One-way ANOVA test with Tukey's multiple comparison test). H, real-time PCR analysis of Fut8 mRNA level. Gapdh was used as a loading control. The Gapdh was used to normalize each value. The value of Fut8 versus Gapdh in Fut8^+/+^ vehicle-treated mice set as 1.0. Data are shown as mean ± SD from three independent experiments. n = 3 mice per group. ∗p < 0.05; ∗∗∗p < 0.001. (One-way ANOVA test with Tukey's multiple comparison test).
Discussion
Our current study demonstrated that excessive microglia activation caused Fut8^+/−^ mice to exhibit depressive-like behaviors and increased neuroinflammatory factors earlier than Fut8^+/+^ mice following exposure to CUS. However, increased core fucosylation through exogenous L-fucose inhibited the overactivation of hippocampal microglia via the JAK2/STAT3 pathway, suppressed subsequent hippocampal neuroinflammation, reversed CUS-induced impairments in synaptic plasticity, and ultimately abolished the depressive-like behavior in Fut8^+/−^ mice (Fig. 9).Figure 9**L-fucose inhibits neuroinflammation and acts as an antidepressant.**Fut8^+/−^ mice exhibited depressive-like behaviors earlier than Fut8^+/+^ mice following CUS and displayed excessive microglia activation along with elevated cytokine expression levels. These changes may impair neuronal synaptic plasticity. Administration of exogenous L-fucose enhanced core fucosylation of gp130 through the salvage pathway for GDP-fucose synthesis, subsequently preventing the complex formation between gp130 and IL-6R, then inhibiting hippocampal microglia activation and subsequent neuroinflammation via the JAK2/STAT3 pathway. This intervention may reverse the synaptic plasticity disorder induced by CUS and ultimately alleviate depressive-like behaviors. Our previous research showed that the suppressive effect of L-fucose on IL-6 induction by LPS was observed only in the Fut8^+/+^ cells, not in the Fut8^−/−^ microglia (45), highlighting the significance of core fucosylation in neuroinflammation. Additionally, complex formation between gp130 and IL-6R was significantly increased in the Fut8^−/−^ cells compared to Fut8^+/+^ cells. Thus, we conclude that L-fucose inhibits neuroinflammation and acts as an antidepressant, at least partly through the upregulation of core fucosylation, which negatively regulates specific cytokine receptors, including the interaction between gp130 and IL-6R. This figure is created with BioRender.com.
Changes in microglia activity are associated with the onset of depression (21, 22). Multiple studies have reported an increase in microglia numbers in the hippocampus of animal models of depression exposed to chronic stress (93, 94, 95, 96). It has also been shown that the density of glial cells in the dentate gyrus of the hippocampus is significantly increased in MDD (97). Our results indicate that the total number of Iba1-positive microglia in the hippocampus DG area of Fut8^+/−^ mice after 28 days of CUS was significantly higher than that in control mice, suggesting that CUS exposure increases microglia numbers, which is suppressed by L-fucose administration. Numerous studies highlight the central role of neuroinflammation in the development of MDD (14, 15). Stress can cause homeostasis dysregulation, resulting in excessive microglia activation, and cause the production of proinflammatory cytokines. This neuroinflammation impairs neurons subsequently and contributes to depressive symptoms (25, 26). This study focused only on microglia. That does not mean that other cells, such as astrocytes and neurons, are not involved in neuroinflammation because core fucosylation is highly expressed in brain tissue (42). Depression is also known to be associated with astrocyte-mediated neuroinflammation (98). This study mainly focused on the relationship between IL-6/gp130-mediated signaling, a key inflammatory response pathway, and core fucosylation, while the IL-6R is present in microglia but not in astrocytes (99). In addition to the neuroinflammation hypothesis, there are other hypotheses about depression, such as the monoamine hypothesis, hypothalamic-pituitary-adrenal axis dysfunction hypothesis, etc. (7). In subsequent studies, we will use new technologies to explore whether other cells or other mechanisms are involved in the relationship between core fucosylation and depression.
Several studies have reported that the levels of multiple inflammatory markers were increased in patients with MDD (15, 16). Notably, alterations in IL-6 serum levels are among the most reproducible abnormalities in depression patients (16). Children with high circulating IL-6 levels have a 10% higher risk of developing MDD in adulthood compared to the general population or children with low IL-6 levels (100). Furthermore, it has been reported that two-thirds of mice exhibit depressive-like behaviors after exposure to social defeat stress (101). In our study, we found that Fut8^+/−^ mice had higher Il6 levels than the Fut8^+/+^ group (Fig. 3A), indicating that diminished core fucosylation predisposes mice to depression (Fig. 2). Additionally, other studies also suggest a link between core fucosylation and neuroinflammation in the central nervous system (CNS). For instance, core-fucosylated N-glycans were significantly decreased in mouse brain tissues treated with LPS (102). FUT8 expression was also significantly reduced in schizophrenia patients compared to controls (103). Patients with Multiple sclerosis exhibited lower levels of core-fucosylated N-glycans compared to matched controls (104), a similar finding also observed in Alzheimer's disease (105). Interestingly, it has been reported that female patients with MDD show decreased core fucosylation and increased IL-6 levels in serum (106). Consistent with these studies, we observed decreased core fucosylation levels in the hippocampus of mice exposed to the CUS model (Figs. 7A, 8G), which was supported by the observation in the BV2 cell model treated with LPS (Fig. 7F). L-fucose treatment of BV2 cells alone did not cause changes in Fut8 mRNA levels. However, L-fucose could reduce the LPS-induced inflammation level in BV2 cells, thereby affecting the Fut8 mRNA level, indicating that inflammation is the cause of alterations in Fut8 mRNA levels (Fig. 7, F and G). It is notable to mention that there are bi-phase responses in LPS induction. The mRNA levels of Fut8 were significantly increased during the shorter time treatment, such as 12 and 24 h, but decreased during the longer time treatment (Fig. 7E). Thus, the phenomenon could be explained by the possibility that Fut8 mRNA levels are upregulated in response to acute inflammatory stimulation and downregulated in response to chronic inflammatory stimulation. This is consistent with the findings that our chronic depression model CUS stimulation mice exhibit reduced Fut8 mRNA levels (Figs. 7B; 8H). The underlying mechanism for L-fucose-upregulating Fut8 has yet to be entirely understood. However, based on the observations in (Fig. 7, D–F), we speculate that chronic inflammation may suppress Fut8 expression, while L-fucose can reduce chronic inflammation, which results in upregulation of Fut8. On the other hand, acute inflammation induces the mRNA level of Fut8 (Fig. 7E). The underlying mechanism for the different responses to acute and chronic inflammation remains to be studied further. These above results further suggest that core fucosylation is important for CNS maintenance and negatively regulates neuroinflammation.
The reduction or loss of core fucosylation may upregulate the pro-inflammatory reaction. Previous research indicated that a lack of core fucosylation leads to emphysema-like changes in Fut8^−/−^ mice (107). Moreover, in a cigarette smoke-induced emphysema model, Fut8^+/−^ mice were more sensitive than Fut8^+/+^ mice (108). After LPS treatment, IL-6 expression levels in the lung and brain tissues of Fut8^+/−^ mice were significantly higher than in Fut8^+/+^ mice, and L-fucose treatment could reduce the IL-6 levels in Fut8^+/−^ mice (45, 108). The GDP-fucose level in Fut8^+/−^ mice was lower than that in Fut8^+/+^ mice, and exogenous administration of L-fucose could significantly increase the GDP-fucose level in Fut8^+/−^ mice (45). Therefore, exogenous L-fucose administration can increase core fucosylation in Fut8^+/−^ mice. Mechanistic studies suggested that core fucosylation negatively regulates the formation of IL-6R and gp130 complex by increasing core fucosylation of gp130 in Fut8^+/−^ mice, this complex can mediate the JAK2/STAT3 pathway (Fig. 5). Similarly, we found that after CUS stress exposure, neuroinflammatory factors, including IL-6, were significantly higher in Fut8^+/−^ mice compared to Fut8^+/+^ mice, while exogenous L-fucose significantly inhibited their expression. It is noteworthy that GDP-fucose from exogenous L-fucose is preferentially utilized by Fut8 (109). This may explain why the expression levels of Lewis X were not increased in mice exposed to CUS with L-fucose treatment (Fig. S4, A and B). Although core fucosylation and other fucosylation, such as α1,3-fucosylation, were reduced after CUS, core fucosylation, not α1,3-fucosylation on Lewis X, could partially be rescued by L-fucose (Figs. 7A; 8G). Our previous study also showed that administration of exogenous L-fucose rescued the reduced IgG amount in Fut8^+/−^ mice (110). We believe that the effects of L-fucose on CUS-induced depression are Fut8-dependent, as a previous study indicated that supplementation of L-fucose suppressed the enhanced IL-6 expression level in Fut8^+/+^ cells, but not in Fut8^−/−^ cells (45). We do not exclude other potential functions of L-fucose, which warrant further investigation. It has been reported that serotonin release is reduced in patients with depression (111). Lower levels of serotonin release in key parts of the brain may lead to stronger suicidal thoughts and higher mortality rates in people with depression (112). In our previous study, we observed the levels of serotonin metabolites were reduced in the Fut8^−/−^ mice compared with Fut8^+/+^ mice (42), suggesting that core fucosylation may be associated with serotonin metabolism. Furthermore, in this experiment, we found that the effect of L-fucose was similar to that of FLX. Perhaps L-fucose administration can increase the level of serotonin in the brain and thus alleviate the depressive-like behavior caused by CUS. We will conduct further exploration on this aspect in the future. Taken together, these data suggest that core fucosylation negatively regulates inflammation induced by CUS exposure.
One of the key contributors to refractory depression is the dysregulation of the JAK2/STAT3 signaling pathway (74). In the nervous system, the JAK-STAT pathway plays a crucial role in neurons, astrocytes, and oligodendrocytes (113, 114, 115). This pathway is also critical in the complex relationship between depression and systemic manifestations of chronic inflammation (116, 117). Inhibition of STAT3 can block IL-6 effects in vitro and regulate depressive-like behaviors in vivo (118). Microglial STAT3 inhibition may improve depressive behavior through neuron-microglia interaction (119). These findings suggest that targeting this pathway could enhance existing depression treatments. In our study, we found that L-fucose administration inhibited the JAK2/STAT3 signaling pathway and reduced neuroinflammation in microglia induced by CUS exposure, indicating that L-fucose can target the JAK2/STAT3 pathway to exert its antidepressant effect. It is notable that although there was no significant difference between L-fucose and WP1066, JAK2/STAT3 inhibitor, in their antidepressant effects (Fig. S2, A–C), the combined administration of L-fucose and WP1066 had a better inhibitory effect on p-JAK2 than L-fucose alone (Fig. S3F). This may indicate that L-fucose exerts anti-inflammatory and antidepressant effects through not only the JAK2/STAT3 pathway but also other signal pathways. And this point needs further exploration.
Higher depression severity has been linked to lower synaptic density in the hippocampus, dorsolateral prefrontal cortex, and anterior cingulate cortex in imaging synaptic density in depressed patients (120). Strong inflammatory activation disrupts the normal microglia structure and function, contributing to depression and neuroplasticity impairment (24). Chronic stress, such as CUS, inhibits the expression of synaptic proteins, including PSD-95, synaptophysin, and decreases dendritic spine density (47, 121, 122). Our study found that the synaptic density in Fut8^+/−^ mice exposed to CUS was significantly reduced in the hippocampus, and the protein expression of PSD-95 was also diminished, likely due to the release of inflammatory factors by microglia, which disrupts synaptic plasticity (Fig. 6, A and B). We also found that synaptic density in Fut8^+/−^ mice was lower than that in Fut8^+/+^ mice (Fig. 6A); this suggests that decreased core fucosylation may affect synaptic density in mice. JAK2 and STAT3 are highly expressed in the brain and are involved in synaptic plasticity (123). Inhibition of neuroinflammation caused by spinal cord injury through the JAK2/STAT3 signaling pathway can prevent neuronal apoptosis (124). IL-6 inhibits LTP, and blocking endogenous IL-6 can enhance LTP duration and long-term memory while reducing neuronal damage (125, 126). The absence of the Fut8 gene significantly suppressed LTP in Fut8^−/−^ mice (43). Interestingly, 30 years ago, Matthies et al. reported that L-fucose and fucosyllactose could enhance hippocampal LTP in vitro (127), although the underlying mechanism was unclear. This study provides a molecular mechanism by which exogenous L-fucose increases core fucosylation of gp130, negatively regulating the JAK2/STAT3 signaling pathway to ameliorate neuroinflammation of CUS maintenance (Figs. 5B; 9).
In summary, L-fucose negatively regulates CUS-induced neuroinflammation and improves synaptic plasticity impairment through the JAK2/STAT3 pathway, ultimately exerting an antidepressant effect, indicating that L-fucose possesses significant anti-neuroinflammatory activity and antidepressant potential.
Experimental procedures
Antibodies and reagents
Biotinylated LCA (J207) was obtained from J-oil Mills. The anti-GAPDH antibody (G9545) and anti-Mouse IgG antibody (AP124P) were obtained from Sigma. Anti-JAK2 (catalog no.: 3230), anti-Phospho-JAK2 (catalog no.: 3771), anti-STAT3 (catalog no.: 9139), anti-Phospho-STAT3 (catalog no.: 9145) antibodies and anti-rabbit IgG, HRP-linked antibody (catalog no.: 7074) were acquired from Cell Signaling Technology. The Iba1 antibody (catalog no. 019–19741) was purchased from FUJIFILM Wako Pure Chemical Corporation, and the anti-PSD-95 antibody (catalog no.:04–1066) was obtained from Millipore Corporation. The ABC kit (catalog no.: PK-4000) was from Vector Laboratories, and the goat anti-rabbit antibody Alexa Fluor 488 (catalog no.: A-11008) was obtained from Invitrogen. L-fucose (F0065) and Anti-Lewis X (A2578) were acquired from TCI, while fluoxetine (56,296–78–7) was sourced from Merck. WP1066 (HY-15312) was acquired from MCE. LPS was obtained from Sigma and purified from Escherichia coli O111:B4 (L2630). Ab-Capcher MAG2 was from ProteNova. The anti-IL-6R (#MA5-29721) and anti-gp130 (#A304–929A) antibodies were purchased from Thermo Fisher Scientific.
Animal
Fut8^+/−^ heterozygous mice with the ICR genetic background were intercrossed to generate Fut8^+/+^ littermates and Fut8^+/−^ mice (42). To investigate the underlying mechanism, we used a sensitive inflammation-monitoring mouse system containing the human interleukin-6 (hIL6) bacterial artificial chromosome (BAC) transgene modified with luciferase (Luc) reporter cassette (128). The Fut8^+/−^ mice were mated with hIL6-BAC-Luc reporter transgenic mice to produce Fut8^+/+^::hIL6-Luc and Fut8^+/−^::hIL6-Luc compound transgenic mice for LPS-induced inflammation as described (45). All experiments were conducted using 5- to 6-week-old male mice weighing approximately 25 to 30 g. Mice were housed in groups under standard vivarium conditions, with free access to food and water, a 12-h light-dark cycle (lights on from 7:00–19:00), an ambient temperature of 22 ± 2 °C, and relative humidity of 55 ± 5%. Mice received oral L-fucose twice daily via gavage at a dose of 36 mg/day. All animal experiments adhered to protocols approved by the Animal Care and Use Committee of the Graduate School of Pharmaceutical Sciences, Tohoku Medical and Pharmaceutical University.
Cell culture
BV2 (mouse microglia cell line) was generously gifted by Professor Elisabetta Blasi (University of Modena and Reggio Emilia, Modena). At 37 °C and 5% CO2, BV2 cells were cultured in Dulbecco's modified Eagle's medium supplemented with 10% fetal bovine serum in a humidified environment. The cells were confirmed free from mycoplasma using the e-Myco Mycoplasma PCR Detection kit (iNtRON Biotechnology, Republic of Korea).
Chronic unpredictable stress (CUS)
Following established procedures (62), we constructed the CUS model by randomly exposing mice to two stressors daily for 28 days (Fut8^+/−^ mice) or 35 days (Fut8^+/+^ mice), as detailed in Table 1. Stressors included 1 h of cage shaking, 12 h of lights-on during the night, 3 h of lights-off during the day, 2 h of light restraint (housing in small cages), 1 h of restraint (placing mice in a 50 ml plastic tube with breathing holes), 14 h of 45°cage tilt, 14 h of wet cage, and 12 h of food or water deprivation during the dark period.Table 1. Construction of the CUS model, in which mice were randomly exposed to two stressors dailyTimesMondayTuesdayWednesdayThursdayFridaySaturdaySundayWeek 1lights-offcage tiltshakinglights-onno waterno foodrestraintlights-onlight restraintwet cagelights-offlights-onshakingcage tiltWeek 2restraintno waterlight restraintno foodlights-offlights-onrestraintshakinglights-offcage tiltshakingwet cagelight restraintno waterWeek 3lights-offcage tiltlight restraintlights-onrestraintno foodshakingcage tiltno waterwet cagelights-offno foodlight restraintlights-onWeek 4restraintno waterlights-offcage tiltshakingwet cagelight restraintlights-onrestraintno foodshakingwet cagelights-offcage tiltWeek 5shakingno foodlights-offlights-onrestraintcage tiltshakingno waterlight restraintwet cagelights-offno foodshakingcage tilt
Tail suspension test (TST)
The TST was performed as previously described (62). Mice were suspended 50 cm above the ground with tape attached to their tails (1 cm from the tip) for 6 min. A blinded researcher recorded the duration of immobility during the last 4 min. Mice were considered immobile only when they were passively suspended and completely motionless. Any mice that climbed up the tail were excluded from the analysis.
Forced swimming test (FST)
The FST was conducted as previously outlined (62). Mice were placed individually in a transparent glass cylinder (height: 25 cm, diameter: 10 cm) filled with 10 cm of water at 25 ± 1 °C for 6 min. An investigator unaware of the study recorded the immobility time during the last 4 min, defined as the duration the mouse floated in the water without struggling, only making movements necessary to keep its head above water.
Sucrose preference test (SPT)
This experiment followed established protocols (62). Mice were given access to two bottles in their cages: one containing 1% sucrose solution and the other containing water. All mice were acclimated to the two-bottle choice condition for 2 days, with bottle positions changed every 6 h to prevent positional preference. After a 24-h period of food and water deprivation, mice had free access to both bottles for 12 h, after which the positions were swapped. The volumes of water and sucrose solution consumed were recorded, and sucrose preference was calculated as the percentage of sucrose solution consumed relative to total fluid intake.
Western blotting and lectin blotting
Equal amounts of proteins were separated by sodium dodecyl sulfate-polyacrylamide gel electrophoresis (SDS-PAGE) and transferred to polyvinylidene difluoride membranes (Millipore Sigma) at 10 V for 1 h. For Western blotting, membranes were blocked with 5% nonfat dry milk for 1.5 h at room temperature, incubated with specific primary antibodies overnight at 4° C, and then with appropriate HRP-conjugated secondary antibodies. For lectin blotting, membranes were blocked in 5% BSA for 1.5 h at room temperature, followed by incubation with biotinylated LCA lectin overnight at 4° C. The Vectastain ABC kit (Vector Laboratories) was then used to probe the immunoreactive bands. Specific proteins were visualized using ECL select reagent (Amersham).
Immunoprecipitation
The MT-360 Micro Tube Mixer was used to combine 1.5 μl anti-gp130 antibody with 15 μl Ab-Capcher MAG2 at 4 °C for 2 hours. After three times of washes with PBS, 500 μg of proteins from each tissue were added and incubated overnight at 4 °C. After two PBS washes, these immunoprecipitates were identified using western blotting and lectin blotting.
Real-time PCR
RNA was extracted using TRIzol reagent (Invitrogen) and reverse-transcribed (1 μg total RNA) into cDNA using PrimeScript RT reagent with gDNA Eraser (Takara). Primer sequences for real-time PCR are shown in Table 2. Real-time PCR was performed using TB Green Premix Ex Taq II (Tli RNaseH Plus) (Takara) under the following conditions: initial denaturation at 95 °C for 30 s, followed by 40 cycles of denaturation at 95 °C for 5 s and annealing at 60 °C for 30 s.Table 2. Primer sequences for real-time PCRTarget genesPrimer sequences (5′-3′)Forward sequencesReverse sequencesIl6CTGCAAGAGACTTCCATCCAGAGTGGTATAGACAGGTCTGTTGGTNF-αAAGTCAACCTCCTCTCTGCCCCGGACTCCGCAAAGTCTAAIl1bTGTCTGAAGCAGCTATGGCTGTTGATGTGCTGCTGCGiNOSATGACTCCCAGCACAAAGGGAACAGCACTCTCTTGCGGACCIba1TCTGAGGAGCTATGAGCCTCCATGTACTTCGTCTTGAAGGCGapdhGTCGTGGAGTCTACTGGTGTCTTGAGATGATGACCCTTTTGGCFut8AACAGAAGCAGCCTTCCACCTACCGATTGTGTAGTCCAGC
Immunofluorescence
Mice were deeply anesthetized with sodium pentobarbital and perfused intracardially with 4% paraformaldehyde in 0.01 M phosphate buffer. After perfusion, brains were removed and fixed in 4% paraformaldehyde for 24 h, followed by dehydration in 30% sucrose solution. Brains were frozen and sectioned at 25 μm using a cryostat. Sections were collected in 24-well plates, washed three times with PBS, permeabilized with 0.3% Triton X-100 for 30 min, and blocked with 4% BSA in PBS for 30 min at room temperature. They were then incubated overnight at 4 °C with 1% BSA, 0.3% Triton X-100, and anti-Iba1 antibody (1:500) in PBS. After three washes with PBS, sections were incubated with the goat anti-rabbit antibody Alexa Fluor 488 for 2 h at room temperature. Following three additional washes in PBS, sections were incubated with 4′,6-diamidino-2-phenylindole (DAPI) for 10 min to label cell nuclei. Sections were then transferred to slides, sealed with fluorescence mounting medium, and examined using a ZEISS LSM 900 confocal microscope. The number of Iba1-labeled microglia was quantified using ImageJ software (https://imagej.net/ij/).
Golgi staining
Golgi staining was performed using FD Rapid GolgiStain Kit (PK401). Following deep anesthesia, the mouse brain was removed and washed with double-distilled water. Brain tissue was treated according to the manufacturer’s protocol. Tissue was sectioned into 100 μm slices, transferred to gelatin-coated microscope slides (catalog no. PO101), and air-dried at room temperature. Sections were rinsed with double-distilled water, stained in the staining solution for 10 min, and then rinsed again. Stained sections were dehydrated in a series of ethanol concentrations (50%, 75%, 95% ethanol, and anhydrous ethanol) and finally cleared with xylene. Mounting with Permount Mounting Medium (E17986–01). Three or four neurons were selected per mice in the CA1 region of the hippocampus, and three mice were used per group. Images were captured using an Olympus IX71 microscope at 10X and 40X magnification. Spine density is defined as the number of protrusions divided by the dendritic shaft length. Total dendritic length and number of branches were measured using ImageJ software.
Statistical analysis
All data are presented as mean ± SD from at least three independent experiments. Statistical analysis was conducted using a one-way analysis of variance (ANOVA) with Tukey's post hoc test via GraphPad Prism 5.0 software (GraphPad Software, Inc). A probability value of p was considered as follows: n.s. (no significance), p > 0.05; ∗p < 0.05; ∗∗p < 0.01; ∗∗∗p < 0.001.
Data availability
All data presented in the figures and tables, and supplementary information of this paper are available.
Supporting information
This article contains supporting Information.
Conflicts of interest
The authors declare that they have no conflicts of interest with the contents of this article.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Zhu Y.J.Huang J.Chen R.Zhang Y.He X.Duan W.X.Autophagy dysfunction contributes to NLRP 1 inflammasome-linked depressive-like behaviors in mice J. Neuroinflammation 21202463817819610.1186/s 12974-023-02995-4PMC 10765763 · doi ↗ · pubmed ↗
- 2Kong S.Chen Y.Huang H.Yang W.Lyu D.Wang F.Efficacy of transcranial direct current stimulation for treating anhedonia in patients with depression: a randomized, double-blind, sham-controlled clinical trial J. Affect. Disord.35020242642733823277610.1016/j.jad.2024.01.041 · doi ↗ · pubmed ↗
- 3Zu X.Xin J.Xie H.Xu X.Shen Y.Wang J.Characteristics of gut microbiota and metabolic phenotype in patients with major depressive disorder based on multi-omics analysis J. Affect. Disord.34420245635763786336210.1016/j.jad.2023.10.104 · doi ↗ · pubmed ↗
- 4Ren F.F.Hillman C.H.Wang W.G.Li R.H.Zhou W.S.Liang W.M.Effects of aerobic exercise on cognitive function in adults with major depressive disorder: a systematic review and meta-analysis Int. J. Clin. Health Psychol.24202410044710.1016/j.ijchp.2024.100447 PMC 1086991938371396 · doi ↗ · pubmed ↗
- 5Zhang Y.Lai S.Zhang J.Wang Y.Zhao H.He J.The effectiveness of vortioxetine on neurobiochemical metabolites and cognitive of major depressive disorders patients: a 8-week follow-up study J. Affect. Disord.35120247998073831107310.1016/j.jad.2024.01.272 · doi ↗ · pubmed ↗
- 6Yu X.Wang S.Wu W.Chang H.Shan P.Yang L.Exploring new mechanism of depression from the effects of virus on nerve cells Cells 122023176710.3390/cells 12131767 PMC 1034031537443801 · doi ↗ · pubmed ↗
- 7Cui L.Li S.Wang S.Wu X.Liu Y.Yu W.Major depressive disorder: hypothesis, mechanism, prevention and treatment Signal Transduct. Target. Ther.92024303833197910.1038/s 41392-024-01738-y PMC 10853571 · doi ↗ · pubmed ↗
- 8Malhi G.S.Mann J.J.Depression Lancet.3922018229923123039651210.1016/S 0140-6736(18)31948-2 · doi ↗ · pubmed ↗