Multi-omics analysis reveals a novel NFE2L3 variant impairing choroidal vasculature development in high myopia and myopic maculopathy
Jiangnan He, Luyao Ye, Hannan Xu, Huanjie Yang, Juan Shen, Menghan Li, Shijun Weng, Dongyue Jiao, Chen Chu, Qijun Liao, Haidong Zou, Jianfeng Zhu, Chenji Wang, Xun Xu

Abstract
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
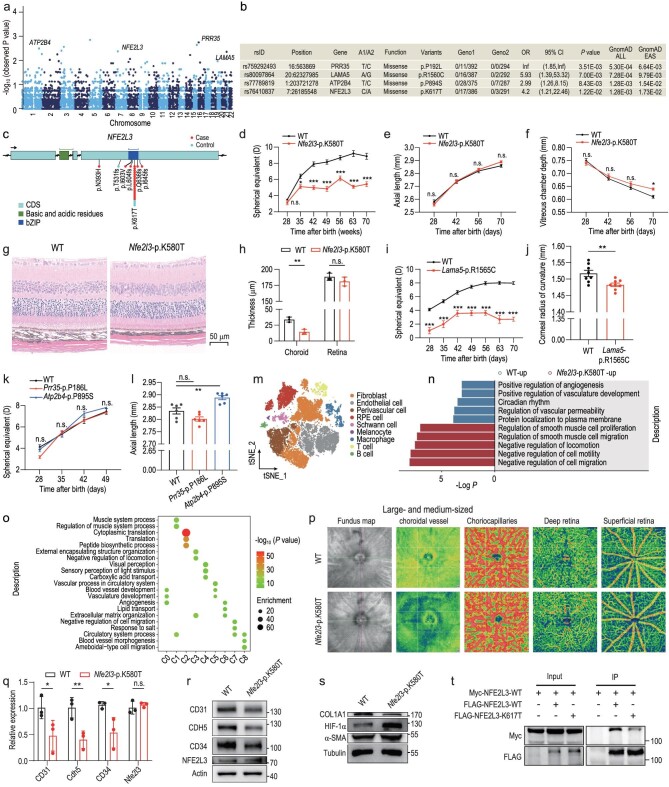 Figure 1
Figure 1- —National Key Research and Development Program of China10.13039/501100012166
- —National Natural Science Foundation of China10.13039/501100001809
- —Natural Science Foundation of Shanghai10.13039/100007219
- —Shanghai Key Disciplines to strengthen the construction of Public health system Three-year action Plan
- —Shanghai Shenkang Hospital Clinical Research Program
- —Science and Technology Research Program of Shanghai
- —China Postdoctoral Science Foundation10.13039/501100002858
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsRetinal Diseases and Treatments · Retinopathy of Prematurity Studies · Retinal Development and Disorders
Myopic maculopathy (MM) is a severe complication of high myopia, characterized by degenerative lesions in the macular retina, choroid and sclera [1]. The pooled prevalence of MM is estimated to be 2.1% in the world population and 47.4% among highly myopic adults [2]. It poses significant challenges due to its poor prognosis and limited treatment options, and has become a major cause of irreversible visual impairment worldwide [3,4]. Notwithstanding, comprehensive investigations into the genetic implications of high myopia and MM remain insufficient. To bridge this gap, we conducted a case–control association study in Chinese individuals, integrating whole-exome sequencing (WES) to identify potential causal genes and investigate their roles in the pathogenesis of high myopia and MM (Fig. S1).
We first analysed a WES data set from the Shanghai High Myopia Study for Adults, which included 697 Han Chinese individuals. Among them, 403 were highly myopic patients with MM, carefully chosen for their representativeness, while the remaining 294 were emmetropic controls with high education levels (Table S1). After the variants were filtered and annotated (Tables S2 and S3), a total of 763 251 coding variants were subjected to further analysis. These included 108 520 common variants (minor allele frequency [MAF] ≥ 5%) and 654 731 low-frequency variants (MAF < 5%). Inflation factor analysis indicated similar genetic backgrounds between the cases and controls (λ = 1.05, Fig. S2a). A total of 726 common variants reached suggestive significance (P < 0.005, Fig. S2b and Table S4). Also, we performed six nested gene-based analyses for low-frequency variants that exhibit larger per-allele effect sizes than common variants and offer crucial biological insights (Fig. S2c) [5]. Four genes, namely PRR35, ATP2B4, NFE2L3 and LAMA5, were identified as reaching suggestive significance (odds ratio > 2 and P < 0.005) based on a deleterious (strict) set (Fig. 1a, Fig. S2d and Table S5). The most significant leading variants for these four candidate genes are listed (Fig. 1b) and the genome location and protein structure of variants are illustrated (Fig. 1c, Figs S2e and S3a–f). Collectively, the WES analysis reveals four causal genes associated with an increased risk of high myopia and MM.
To assess the potential impact of these missense variants on refraction development, we generated four strains of systemic knock-in (KI) mice (Fig. S4a–d and Table S6). Compared with wild-type (WT) mice, Nfe2l3-p.K580T (homologous to p.K617T in humans) mice displayed myopic refractive shift from P35 to P70 (all P < 0.05; Fig. 1d). Ocular biometric measurements revealed a trend for longer axial length (AL) without statistical significance (2.86 ± 0.03 versus 2.89 ± 0.02 mm; P = 0.33; Fig. 1e) and significantly longer vitreous chamber depth (VCD; 0.61 ± 0.02 versus 0.64 ± 0.01 mm; P = 0.02; Fig. 1f) in Nfe2l3-p.K580T mice at P70. The anterior chamber depth, lens thickness and corneal radius of curvature (CRC) were similar between the two groups (Fig. S5a–c). Additionally, hematoxylin and eosin (H&E) staining demonstrated that Nfe2l3-p.K580T mice exhibited significantly thinner choroidal thickness but indistinguishable retinal thickness compared with WT mice at P70 (Fig. 1g and h). Lama5-p.R1565C (homologous to p.R1560C in humans) mice exhibited myopia from P28 to P70 compared with WT mice (all P < 0.001; Fig. 1i). There were no significant differences in axial components between the two groups (Fig. S5d–g) but the CRC of Lama5-p.R1565C mice was significantly smaller than that of WT mice at P70 (1.517 ± 0.022 versus 1.483 ± 0.011 mm; P = 0.007; Fig. 1j). H&E staining and optical coherence tomography imaging showed normal retinal structure of Lama5-p.R1565C mice at P70 (Fig. S5h–j). However, transmission electron microscopy revealed disrupted membrane discs in both Nfe2l3-p.K580T and Lama5-p.R1565C mice (Fig. S5k). Refraction and ocular biometric measurements indicated that Atp2b4-p.P895S (homologous to p.P894S in humans) and Prr35-p.P186L (homologous to p.P192L in humans) mice displayed comparable ocular development to WT mice, except that Atp2b4-p.P895S mice had a significantly longer AL (2.83 ± 0.02 versus 2.89 ± 0.02 mm; P = 0.004) (Fig. 1k and l). Collectively, we validate that Nfe2l3-p.K580T and Lama5-p.R1565C mice displayed myopic refractive shifts that were attributed to elongated VCD and smaller CRC, respectively.
Given that neither Atp2b4-p.P895S nor Prr35-p.P186L mice displayed a myopic phenotype, and the reduced CRC observed in Lama5-p.R1565C mice did not coincide with the continuous axial elongation seen in humans with high myopia and MM, we focused our investigations on discerning the mechanisms underlying the myopic phenotype observed in Nfe2l3-p.K580T mice. Considering the thinner choroidal thickness observed in the mutant mice, which is a typical characteristic during the development and progression of high myopia and MM [6], we conducted single-cell ribonucleic acid sequencing (scRNA-seq) on choroid samples from Nfe2l3-p.K580T and WT mice at P70 (Table S7). By analysing the expression of markers and referring to recent scRNA-seq studies of choroids [7], we identified nine distinct cell types (Fig. 1m, Fig. S6a–c and Table S8). Enrichment analysis using the differentially expressed genes within each subcluster was conducted on gene ontology (Table S9). The 49 downregulated genes in endothelial cells (ECs) were enriched in processes related to the positive regulation of angiogenesis, vasculature development and vascular permeability. Conversely, the 43 upregulated genes in ECs were enriched in processes such as the negative regulation of locomotion, cell mobility and cell migration (Fig. 1n and Fig. S6d). We further identified eight transcriptionally distinct subpopulations of ECs (Fig. S6e). Subpopulations that were predominant in WT mice (C0, C2 and C5) demonstrated enrichment in genes associated with blood vessel development, vasculature development and angiogenesis. In contrast, subpopulations that were predominant in Nfe2l3-p.K580T mice (C1, C3 and C7) were associated with extracellular matrix organization and negative regulation of locomotion and cell migration (Fig. 1o and Fig. S6f). These results indicated a potential association between ECs in Nfe2l3-p.K580T mice and the negative regulation of vasculature development.
To further validate the choroidal vascular dysfunction in Nfe2l3-p.K580T mice, we performed an in vivo experiment involving optical coherence tomography angiography (OCT-A) imaging of the fundus (Fig. 1p). Quantitative analysis confirmed a reduction in blood flow within large and medium-sized choroidal vessels, choriocapillaris and deep retina in the Nfe2l3-p.K580T group compared with the control group (Fig. S7a–c). No significant difference was found in superficial retinal blood flow layers between the two groups (Fig. S7d). These results indicated a decreased blood flow perfusion in the choroid of Nfe2l3-p.K580T mice. Furthermore, quantitative real-time reverse transcription polymerase chain reaction and Western blot analyses revealed decreased expression of vascular EC markers in the choroids of Nfe2l3-p.K580T mice (Fig. 1q–r and Fig. S7e and f). In the sclera of Nfe2l3-p.K580T mice, an elevated expression of HIF-1α and a decreased expression of COL1A1 were observed (Fig. 1s and Fig. S7g). Collectively, these findings indicate that the Nfe2l3-p.K580T variant impairs choroidal vascularization and causes scleral hypoxia and extracellular matrix remodeling [8], contributing to the development of myopia in mice.
The NFE2L3-p.K617T, identified as a potential pathogenic candidate, was exclusive to the East Asian population (Fig. S8a). Its origins can be traced back by ∼44 503 years, which corresponds to 1780 generations (Fig. S8b). NFE2L3 is a transcription factor belonging to the CNC-bZIP family [9], with the bZIP domain playing a crucial role in dimerization and DNA binding [10]. Most of the variants of NFE2L3 found in the WES data were located in the bZIP domain (Fig. 1c). We examined the interaction affinity between FLAG- and Myc-tagged NFE2L3 proteins. Compared with the FLAG-NFE2L3-WT, less Myc-NFE2L3 was immunoprecipitated by FLAG-NFE2L3-p.K617T (Fig. 1t). These results indicated that variants in the bZIP domain, specifically p.K617T, hinder the dimerization of NFE2L3. This impairment of dimerization may have negative effects on the function of NFE2L3 as a transcription factor.
In conclusion, we have identified a novel variant, NFE2L3-p.K617T, that increases the risk of high myopia and MM in Chinese heterozygous carriers using WES and demonstrated its deleterious roles using KI mice models. Mechanistic investigations revealed that this variant impairs choroidal vasculature development, leading to decreased choroidal blood flow perfusion, ultimately resulting in scleral hypoxia and myopia development. Although this low-frequency variant may only explain a portion of the phenotypic variance, our findings have enhanced the understanding of the biological mechanisms involved in high myopia and MM, and provide insights for potential therapeutic targets.
Supplementary Material
nwae291_Supplemental_Files
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Ohno-Matsui K, Lai TY, Lai CC et al. Prog Retin Eye Res 2016; 52: 156–87. 10.1016/j.preteyeres.2015.12.00126769165 · doi ↗ · pubmed ↗
- 2Zou M, Wang S, Chen A et al. Br J Ophthalmol 2020; 104: 1748–54. 10.1136/bjophthalmol-2019-31529832188679 · doi ↗ · pubmed ↗
- 3Fang Y, Yokoi T, Nagaoka N et al. Ophthalmology 2018; 125: 863–77. 10.1016/j.ophtha.2017.12.00529371011 · doi ↗ · pubmed ↗
- 4Fricke TR, Jong M, Naidoo KS et al. Br J Ophthalmol 2018; 102: 855–62. 10.1136/bjophthalmol-2017-31126629699985 PMC 6047154 · doi ↗ · pubmed ↗
- 5Weiner DJ, Nadig A, Jagadeesh KA et al. Nature 2023; 614: 492–9. 10.1038/s 41586-022-05684-z 36755099 PMC 10614218 · doi ↗ · pubmed ↗
- 6Fang Y, Du R, Nagaoka N et al. Ophthalmology 2019; 126: 1018–32. 10.1016/j.ophtha.2019.01.01230703442 · doi ↗ · pubmed ↗
- 7Voigt AP, Mulfaul K, Mullin NK et al. Proc Natl Acad Sci USA 2019; 116: 24100–7. 10.1073/pnas.191414311631712411 PMC 6883845 · doi ↗ · pubmed ↗
- 8Wu H, Chen W, Zhao F et al. Proc Natl Acad Sci USA 2018; 115: E 7091–100. 10.1073/pnas.172144311529987045 PMC 6064999 · doi ↗ · pubmed ↗