Natural infection of SARS-CoV-2 variant XBB.1.9.1.4.1 in laboratory Syrian hamsters
Chunmao Zhang, Zhendong Guo

Abstract
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
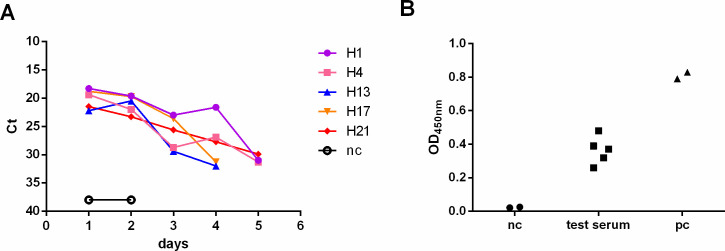 Fig 1
Fig 1Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsSARS-CoV-2 and COVID-19 Research · COVID-19 Clinical Research Studies · SARS-CoV-2 detection and testing
LETTER
At least 18 different animal species, spanning pets, farm animals, captives, and wildlife, have been reported to be infected with SARS-CoV-2 (1). These outbreaks have raised concerns about the potential for human-to-animal, animal-to-animal, and animal-to-human transmission. Notably, the Syrian hamster serves both as a household pet and as an experimental animal model. As a good small animal model, Syrian hamster has been used to study the pathogenesis and transmissibility of SARS-CoV-2 (2, 3), as well as to evaluate the efficacy of vaccines and antiviral drugs (4, 5). Syrian hamsters can also be experimentally infected by different SARS-CoV-2 variants, such as Alpha, Beta, Delta, and Omicron variants (6–12). Recently, two research teams from Hongkong University independently reported the potential transmission of the SARS-CoV-2 Delta strain from pet hamsters to humans (13, 14). Despite these findings, there have been no documented cases of natural SARS-CoV-2 infection in laboratory Syrian hamsters. Here, we presented the first report of natural infection with SARS-CoV-2 Omicron variant XBB.1.9.1.4.1 in laboratory Syrian hamsters.
In September 2023, prior to commencing our animal experimental study, we collected nasal wash samples from a batch of newly ordered Syrian hamsters and promptly conducted routine SARS-CoV-2 screening using a rapid antigen test kit. Unexpectedly, five of these samples tested strongly positive for SARS-CoV-2. To further assess the viral presence, we sequentially collected nasal washes from these five hamsters and determined their viral load using RT-qPCR. Nasal washes of two naive hamsters from another batch ordered before were set as the negative control. In the first 2 days, Ct values in nasal wash samples were below 22, indicating a relatively high viral load, which subsequently gradually reduced to a significantly lower level (Fig. 1A).
The cycle threshold values (Ct) for nasal washes and OD450nm for N protein antibody detection in hamster serum. (A) nc represents negative control of naive hamster nasal washes. (B) nc represents negative control of two naive hamster serum, and pc represents positive control.
Fourteen days after the initial positive tests, serum samples were collected from these five Syrian hamsters, and all serum samples tested positive for SARS-CoV-2 N protein antibody by ELISA (Fig. 1B). Moreover, for nasal wash samples with Ct values less than 22, we conducted virus isolation using Vero-E6 cells, and live SARS-CoV-2 was isolated from nasal washes of hamster H1. The full viral genome was then amplified by multiple overlapping PCR reactions using a panel of primers specific for SARS-CoV-2 and sequenced using an ABI3730XL sequencer. Using the Nextstrain platform, we identified the pangolin lineage of the isolated virus, which belonged to the FL.4.11 clade. The unaliased name assigned to this particular variant was XBB.1.9.1.4.1.
In summary, we have reported the first natural infection of the SARS-CoV-2 Omicron variant XBB.1.9.1.4.1 in laboratory Syrian hamsters, as well as the successful isolation and characterization of this virus. Given the highly experimental susceptibility of Syrian hamsters to a series of SARS-CoV-2 variants (Alpha, Beta, Gamma, Delta, and Omicron) and the emerging variants (6–12, 15), and considering the transmission of Delta variant AY.127 from pet hamsters to humans, which led to a pet-shop-related COVID-19 outbreak in Hong Kong (13, 14), it is imperative to strengthen the monitoring of COVID-19 in laboratory animals, pet animals, and wild animals and reinforce biosafety management measures for laboratory animals.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Cui SJ, Liu YM, Zhao JC, Peng XM, Lu GL, Shi WX, Pan Y, Zhang DT, Yang P, Wang QY. 2022. An updated review on SARS-Co V-2 infection in animals. Viruses 14:1527. doi:10.3390/v 1407152735891507 PMC 9323600 · doi ↗ · pubmed ↗
- 2Chan JF-W, Zhang AJ, Yuan S, Poon VK-M, Chan CC-S, Lee AC-Y, Chan W-M, Fan Z, Tsoi H-W, Wen L, Liang R, Cao J, Chen Y, Tang K, Luo C, Cai J-P, Kok K-H, Chu H, Chan K-H, Sridhar S, Chen Z, Chen H, To KK-W, Yuen K-Y. 2020. Simulation of the clinical and pathological manifestations of coronavirus disease 2019 (COVID-19) in a golden Syrian hamster model: implications for disease pathogenesis and transmissibility. Clin Infect Dis 71:2428–2446. doi:10.1093/cid/ciaa 32532215622 PMC 7184405 · doi ↗ · pubmed ↗
- 3Sia SF, Yan L-M, Chin AWH, Fung K, Choy K-T, Wong AYL, Kaewpreedee P, Perera RAPM, Poon LLM, Nicholls JM, Peiris M, Yen H-L. 2020. Pathogenesis and transmission of SARS-Co V-2 in golden hamsters. Nature 583:834–838. doi:10.1038/s 41586-020-2342-532408338 PMC 7394720 · doi ↗ · pubmed ↗
- 4Tostanoski LH, Wegmann F, Martinot AJ, Loos C, Mc Mahan K, Mercado NB, Yu J, Chan CN, Bondoc S, Starke CE, et al.. 2020. Ad 26 vaccine protects against SARS-Co V-2 severe clinical disease in hamsters. Nat Med 26:1694–1700. doi:10.1038/s 41591-020-1070-632884153 PMC 7671939 · doi ↗ · pubmed ↗
- 5Rosenke K, Jarvis MA, Feldmann F, Schwarz B, Okumura A, Lovaglio J, Saturday G, Hanley PW, Meade-White K, Williamson BN, Hansen F, Perez-Perez L, Leventhal S, Tang-Huau T-L, Callison J, Haddock E, Stromberg KA, Scott D, Sewell G, Bosio CM, Hawman D, de Wit E, Feldmann H. 2020. Hydroxychloroquine prophylaxis and treatment is ineffective in macaque and hamster SARS-Co V-2 disease models. JCI Insight 5:e 143174. doi:10.1172/jci.insight.14317433090972 PMC 7714406 · doi ↗ · pubmed ↗
- 6Ulrich L, Halwe NJ, Taddeo A, Ebert N, Schön J, Devisme C, Trüeb BS, Hoffmann B, Wider M, Fan X, et al.. 2022. Enhanced fitness of SARS-Co V-2 variant of concern Alpha but not Beta. Nature 602:307–313. doi:10.1038/s 41586-021-04342-034937050 PMC 8828469 · doi ↗ · pubmed ↗
- 7Saito A, Irie T, Suzuki R, Maemura T, Nasser H, Uriu K, Kosugi Y, Shirakawa K, Sadamasu K, Kimura I, et al.. 2022. Enhanced fusogenicity and pathogenicity of SARS-Co V-2 Delta P 681R mutation. Nature 602:300–306. doi:10.1038/s 41586-021-04266-934823256 PMC 8828475 · doi ↗ · pubmed ↗
- 8Rosenke K, Okumura A, Lewis MC, Feldmann F, Meade-White K, Bohler WF, Griffin A, Rosenke R, Shaia C, Jarvis MA, Feldmann H. 2022. Molnupiravir inhibits SARS-Co V-2 variants including Omicron in the hamster model. JCI Insight 7:e 160108. doi:10.1172/jci.insight.16010835579953 PMC 9310535 · doi ↗ · pubmed ↗