TccP4: a novel effector identified in the Escherichia albertii strain 1551-2 required for attaching and effacing lesion formation on infected Nck-null cells
Iranildo do A. Fernandes, Tadasuke Ooka, Daiany R. P. de Lira, Fernando H. Martins, Henrique Orsi, Nina Jones, Waldir P. Elias, Tetsuya Hayashi, Tânia A. T. Gomes, Rodrigo T. Hernandes

TL;DR
Researchers discovered a new protein, TccP4, in a type of E. albertii bacteria that helps it form structures on host cells, contributing to its ability to cause diarrhea.
Contribution
The study identifies TccP4 as a novel TccP subtype essential for attaching and effacing lesion formation in Nck-null cells.
Findings
TccP4 is required for F-actin pedestal formation in infected Nck-null cells, with deletion causing a 96% reduction in this ability.
The tccP4 gene is widely distributed in E. albertii strains and likely acquired early during the species' diversification.
The genetic region containing tccP4 is highly variable and contains mobile genetic elements.
Abstract
Escherichia albertii is a pathogen that causes sporadic cases and outbreaks of diarrhea. The main virulence feature of this bacterium is the attaching and effacing (AE) lesion formation on infected intestinal epithelial cells, which is characterized by the formation of pedestal-like structures that are rich in F-actin. The Brazilian E. albertii 1551-2 strain can recruit F-actin using both the Nck-dependent and the Nck-independent pathways, the latter of which uses an adaptor protein named Tir-cytoskeleton coupling protein (TccP/EspFU). Genome analyses of the 1551-2 strain unveiled the existence of a gene encoding a putative novel TccP subtype in addition to a gene encoding for the TccP3 subtype. Amino-acid sequence comparison with known TccP subtypes (TccP/EspFU, TccP2, and TccP3) confirmed that the protein represents a novel TccP subtype—named here TccP4. Lack of TccP4 led to an…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
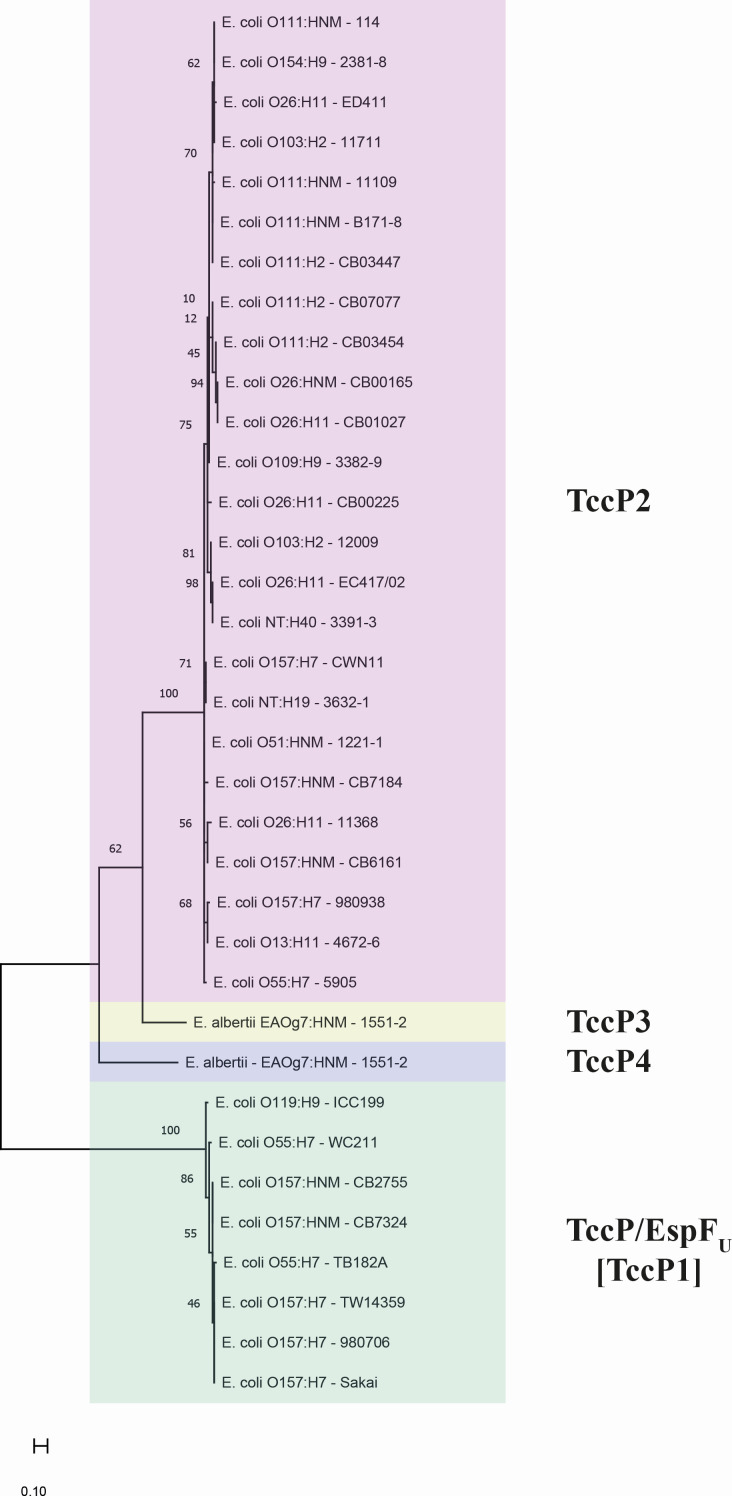 Fig 1
Fig 1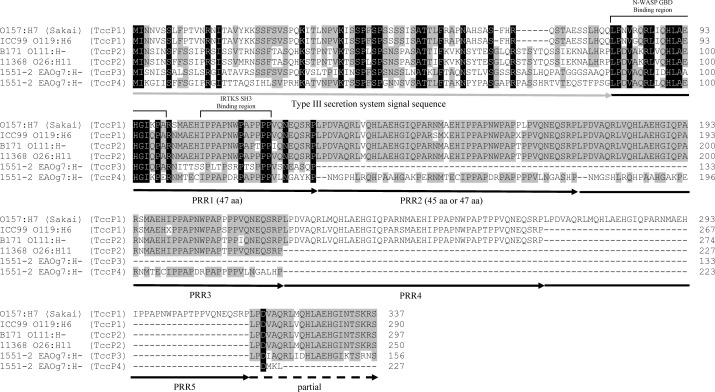 Fig 2
Fig 2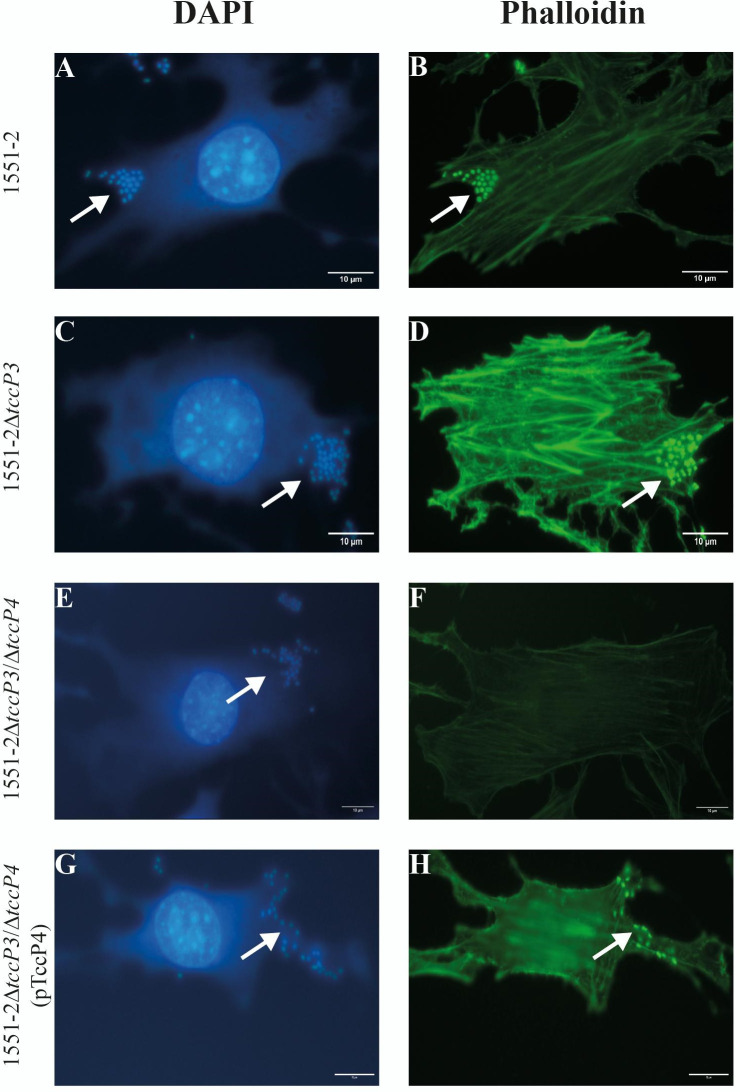 Fig 3
Fig 3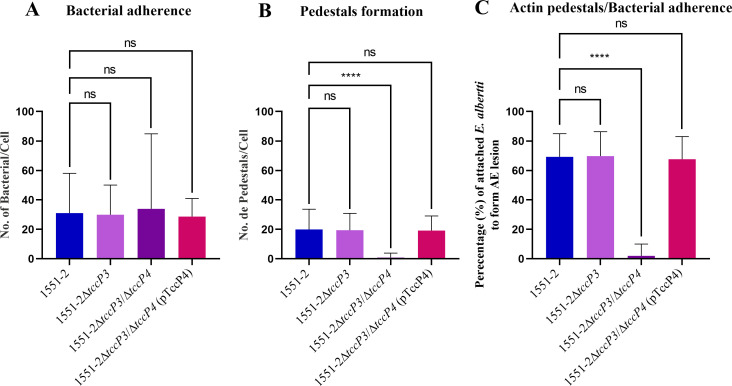 Fig 4
Fig 4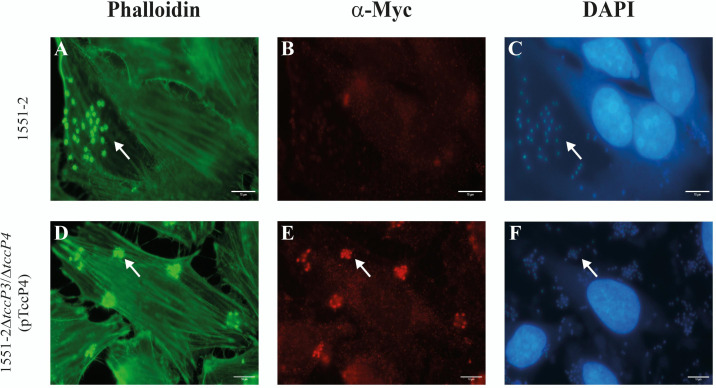 Fig 5
Fig 5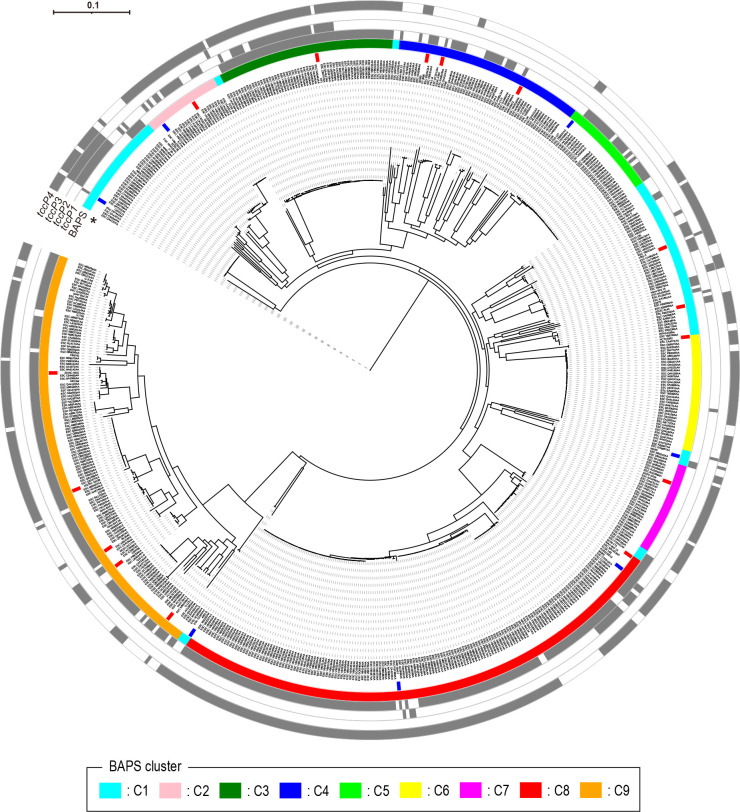 Fig 6
Fig 6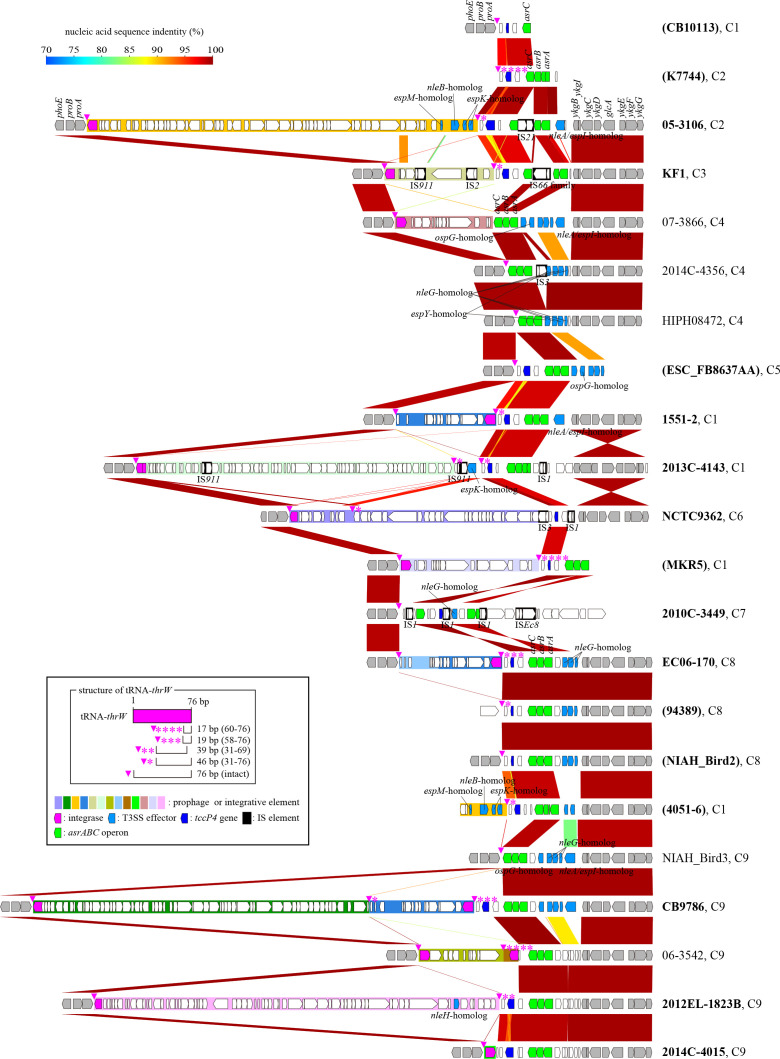 Fig 7
Fig 7| Strains and plasmids | Relevant characteristics | Reference |
|---|---|---|
| Strains | ||
| 1551-2 | ( | |
| 1551-2 | 1551-2 | ( |
| 1551-2 | 1551-2 | This study |
| 1551-2 | 1551-2 | This study |
| 1711-4Δ | Source of zeocin cassette | ( |
| Plasmids | ||
| pKOBEG-Apra | Red recombinase system plasmid, ApraR | ( |
| pKC471 | pK187 vector harboring the | ( |
| pTccP4 | pKC471 vector harboring the | This study |
| BAPS cluster | No. of strains | Distinct TccP-encoding genes subtypes detected: | |||
|---|---|---|---|---|---|
|
|
|
|
| ||
|
| 103 | 19 (18.4%) | 31 (30.1%) | 22 (21.4%) | 71 (68.9%) |
|
| 25 | 0 (0.0%) | 15 (60.0%) | 0 (0.0%) | 25 (100.0%) |
|
| 56 | 56 (100.0%) | 31 (55.4%) | 0 (0.0%) | 55 (98.2%) |
|
| 58 | 21 (36.2%) | 0 (0.0%) | 0 (0.0%) | 5 (8.6%) |
|
| 30 | 24 (80.0%) | 0 (0.0%) | 0 (0.0%) | 3 (10.0%) |
|
| 37 | 0 (0.0%) | 1 (2.7%) | 0 (0.0%) | 32 (86.5%) |
|
| 29 | 0 (0.0%) | 0 (0.0%) | 0 (0.0%) | 27 (93.1%) |
|
| 164 | 152 (92.7%) | 19 (11.6%) | 0 (0.0%) | 144 (87.8%) |
|
| 135 | 122 (90.4%) | 0 (0.0%) | 0 (0.0%) | 90 (66.7%) |
|
| 637 | 394 (61.9%) | 97 (15.2%) | 22 (3.5%) | 452 (71.0%) |
| Primer identification | Primer sequence (5′’→ 3′) | PCR conditions: | Fragment size (bp) | Reference | |
|---|---|---|---|---|---|
| Annealing temperature | Extension time | ||||
| tccP4-zeo-F |
| 55°C | 45 sec | 659 | This study |
| tccP4-zeo-R |
| ||||
| tccP4-F |
| 67°C | 45 sec | 540 | This study |
| tccP4-R |
| ||||
| zeo-F |
| 55°C | 45 sec | 559 | ( |
| zeo-R |
| ||||
| tccP4-Flank-F |
| 55°C | 1 min | 965 | This study |
| tccP4-Flank-R |
| ||||
| tccP4-XbaI-F |
| 55°C | 45 sec | 781 | This study |
| tccP4-BamHI-R |
| ||||
| tccP/EHEC-zeo-F |
| 55°C | 45 sec | 659 | This study |
| tccP/EHEC-zeo-R |
| ||||
- —Fundação de Amparo à Pesquisa do Estado de São Paulo (FAPESP)
- —MEXT | Japan Society for the Promotion of Science (JSPS)
- —Fundação de Amparo à Pesquisa do Estado de São Paulo (FAPESP)
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsEscherichia coli research studies · Enterobacteriaceae and Cronobacter Research · Viral gastroenteritis research and epidemiology
INTRODUCTION
The species Escherichia albertii represents one of the new members of the genus Escherichia that is responsible for sporadic cases, as well as outbreaks of diarrhea, in several countries (1–4). E. albertii colonizes the human intestinal mucosa and, similarly to enteropathogenic (EPEC) and enterohemorrhagic (EHEC) E. coli, induces the attaching and effacing (AE) lesion formation in the host cells (5–8). The AE lesion is characterized by intimate adherence, microvilli destruction, and the formation of a pedestal-like structure that is rich in F-actin and other eukaryotic cytoskeleton elements (9, 10).
The genes encoding proteins involved in the AE lesion formation are located within a pathogenicity island (PAI) termed the locus of enterocyte effacement or LEE region (11). The LEE region is organized in five polycistronic operons, termed LEE1 to LEE5, two bicistronic operons (espG-rorf1 and grlA-grlR), and four individual genes (etgA, cesF, map, and escD) (12). This PAI encodes a type 3 secretion system (T3SS), as well as the adhesin intimin (13) and Tir (translocated intimin receptor) which is the intimin receptor translocated to host cells via the T3SS (14).
During the establishment of the AE lesion, the recruitment of F-actin for pedestal formation in host cells can occur via two distinct pathways in which the participation of the eukaryotic adapter protein Nck (non-catalytic region of tyrosine kinase) may be necessary or not (15). These two pathways have been referred to in the literature as Nck-dependent and Nck-independent (13, 16, 17). In the Nck-dependent pathway, first described in the typical EPEC prototype strain E2348/69 (14, 18), the tyrosine residue 474 (Y_474_) present in the C-terminal domain of the Tir protein is phosphorylated by host cell kinases triggering the recruitment of Nck to the bacterial adherence site. Subsequently, the actin nucleation-promoting factor neuronal Wiskott-Aldrich syndrome protein (N-WASP) and the actin-related protein 2/3 (Arp2/3) complex are sequentially activated, initiating the actin polymerization processes (18, 19).
On the other hand, the Tir protein produced by EHEC strains of serotype O157:H7 lacks the Y_474_ residue and, therefore, this pathogen does not use the Nck adaptor protein to recruit actin for the AE lesion formation (16). Instead, in an Nck-independent pathway, EHEC O157:H7 requires another bacterial effector translocated by the T3SS, termed Tir-cytoskeleton coupling protein (TccP) or EspF_U_, which is linked to the C-terminal domain of Tir via the eukaryotic adaptor protein insulin receptor tyrosine kinase substrate (IRTKS), resulting in the activation of N-WASP and Arp2/3 (17, 20, 21). Of note, IRTKS and insulin receptor substrate of 53 kDa protein (IRSp53) are related proteins serving as adaptors of F-actin recruitment (22). So far, three distinct subtypes of TccP/EspF_U_ have been described and designated as follows: TccP/EspF_U_ (referred to as TccP1 in this study), TccP2, and TccP3 (17, 20, 23, 24).
Previous studies have shown that the Brazilian E. albertii prototype 1551-2 strain can employ both the Nck-dependent (25) and the Nck-independent pathways for F-actin polymerization during AE lesion formation (24). Even though TccP3 was first described in the 1551-2 strain, a tccP3 deletion mutant preserved its ability to recruit F-actin in an Nck-independent pathway in assays performed with mouse embryonic fibroblasts lacking Nck (Nck-null MEF cells) (24).
Further, genomic analyses searching for genes with the potential to encode novel T3SS effectors revealed that the E. albertii 1551-2 strain harbors a gene encoding a novel TccP subtype termed, in the present study, tccP4. In view of this scenario, our main objective was to evaluate the contribution of TccP4 in the AE lesion formation in an Nck-independent pathway, as well as understand the distribution of the tccP4 gene among other sequenced E. albertii strains.
RESULTS
TccP4: a novel TccP subtype identified in the E. albertii 1551-2 strain
During the genomic analysis of the E. albertii 1551-2 strain, we identified a gene able to potentially encode a novel TccP subtype with 227 amino acids in length and approximately 24 kDa in molecular weight (Table S1; supplemental material is found at DOI 10.6084/m9.figshare.28152620). To test this hypothesis, its amino acid sequence (Protein ID: AUS64398.1) was employed to carry out a phylogenetic analysis through the construction of a maximum likelihood (ML) tree, as well as to determine the percentage of amino acid sequence identity between different subtypes of TccP described to date (TccP1, TccP2, and TccP3). In the phylogenetic tree constructed, TccP proteins were separated into four main branches, three of which were represented by the three TccP subtypes already described in the literature (TccP1, TccP2, and TccP3), while the fourth branch represented the novel TccP subtype—termed TccP4—identified in this study (Fig. 1). Furthermore, TccP4 showed 60% or less of identity to the representative amino acid sequences of other TccP subtypes (Table S1; supplemental material is found at DOI 10.6084/m9.figshare.28152620), thus providing additional evidence that the protein is a novel TccP subtype.
Identification of a novel TccP subtype—termed TccP4—in the E. albertii 1551-2 strain. A phylogenetic analysis of the amino acid sequences of the distinct TccP subtypes (TccP1, TccP2, and TccP3), using the maximum likelihood method, showed four main branches representing the three TccP subtypes described to date, as well as the novel TccP subtype—TccP4—identified in this study. Each TccP subtype is indicated by a specific color as follows: green: TccP/EspF (TccP1), red: TccP2, yellow: TccP3, and blue: the novel TccP subtype—TccP4.
TccP4 of the 1551-2 strain harbored three proline-rich repeats (PRRs). While the first one was 47 amino acids in length and showed a high sequence identity to the PRRs of other TccP subtypes, the second and third ones were 45 amino acids in length and their sequences were divergent from the first PRR, as well as from the PRR present in other TccP subtypes (Fig. 2). In addition, TccP4 of 1551-2 strain lacked the partial PRR located at the C-terminus observed in the other subtypes.
Amino acid sequence comparison of the TccP4 protein, identified in the E. albertii 1551-2 strain, with previously described TccP subtypes. The gray arrow indicates the type III secretion system signal sequence of the TccP family proteins. Proline-rich repeats (PRRs) are indicated by black arrows, while a partial PRR is indicated by a dashed black arrow. Amino acids marked in black and gray indicate amino acid residues, perfectly and partially conserved among the TccP subtypes, respectively. The type III secretion system signal sequence and the PRRs were marked as previously identified in the TccP/EspFU (also referred to as TccP1) protein from O157:H7 enterohemorrhagic E. coli (26).
TccP4 is required for attaching and effacing lesion formation in infected host cells
To evaluate the functionality of TccP4 in the recruitment of F-actin for AE lesion formation, we generated a tccP4 deletion mutant in the 1551-2ΔtccP3 background, thus generating the 1551-2ΔtccP3/tccP4 double mutant. The replacement of tccP4 by the zeocin-encoding resistance gene (ble) was confirmed by Sanger sequencing (data not shown). The 1551-2ΔtccP3/tccP4 double mutant carrying pTccP4 was also generated (Table 1), in which the expression of the recombinant TccP4-Myc protein was confirmed by immunoblotting using monoclonal α-Myc antibodies (Fig. S1; supplemental material is found at DOI 10.6084/m9.figshare.28152620). All four strains used in the subsequent steps of this study exhibited similar growth rates, as evidenced by the bacterial growth curves (Fig. S2A; supplemental material is found at DOI 10.6084/m9.figshare.28152620).
Nck-null MEF cells were used to evaluate the role of the TccP4 protein in F-actin recruitment for AE lesion formation in a Nck-independent pathway. First, we tested two prototype E. coli strains, E2348/69 (that uses the Nck-dependent pathway) and EDL933 (that uses the Nck-independent pathway), in both HeLa and Nck-null MEF cells. As expected, while both strains induced the recruitment of F-actin in HeLa cells, only the EDL933 strain, which produces the adapter protein TccP/EspF_U_ (referred to in the present study as TccP1), was able to trigger F-actin to the bacterial adherence site in Nck-null MEF cells (Fig. S3; supplemental material is found at DOI 10.6084/m9.figshare.28152620).
First, we confirmed that both 1551-2 and the 1551-2ΔtccP3 strains were able to produce F-actin polymerization underneath the adherent bacteria in infected Nck-null MEF cells (Fig. 3). By contrast, we observed that the 1551-2ΔtccP3/tccP4 double mutant was unable to polymerize F-actin for AE lesion formation in Nck-null MEF cells. This phenotype was restored in the complemented 1551-2ΔtccP3/tccP4(pTccP4) strain (Fig. 3). As a control, we observed that the additional deletion of the tccP4 gene in the 1551-2ΔtccP3 mutant did not affect its ability to induce F-actin polymerization in HeLa cells (Fig. S4; supplemental material is found at DOI 10.6084/m9.figshare.28152620).
TccP4 is required for F-actin polymerization by 1551-2 strain in Nck-null MEF cells. Host cells and adherent bacteria were visualized with DAPI (4′,6-diamidino-2-phenylindole dihydrochloride) staining (A, C, E, and G), while the phalloidin staining allowed the visualization of polymerized F-actin underneath adherent bacteria (B, D, and H). Of importance, the E. albertii 1551-2ΔtccP3/ΔtccP4 double mutant was unable to trigger F-actin for AE lesion formation in infected Nck-null MEF cells (F), thus indicating the importance of the TccP4 protein for the establishment of this phenotype. White arrows in panels A, C, E, and G indicate bacteria adhered to the host cells, while in panels B, D, and H indicate polymerized F-actin. Scale bar: 10 µm.
To better represent the results described above, quantitative assays of bacterial adherence and formation of F-actin-rich pedestals were carried out in Nck-null MEF cells. First, we confirmed that the ability of the 1551-2ΔtccP3 mutant to induce F-actin polymerization in infected Nck-null MEF cells did not differ from that of the wild-type. By contrast, the lack of TccP4 led to a reduction of approximately 96% in the ability of the 1551-2ΔtccP3/tccP4 double mutant strain to induce F-actin-rich pedestals formation in the infected Nck-null MEF cells when compared with the wild-type 1551-2 E. albertii strain (P < 0.0001) (Fig. 4). The ability to form F-actin-rich pedestals was quantitatively restored in the complemented 1551-2ΔtccP3/tccP4(pTccP4) strain (Fig. 4B and C). Of note, the mutation in the tccP4 gene did not affect the adherence ability of the 1551-2ΔtccP3/tccP4 double mutant strain (Fig. 4A). Moreover, we showed that the recombinant TccP4-Myc protein, produced by the 1551-2ΔtccP3/tccP4(pTccP4) complemented strain, colocalized with the polymerized F-actin, detected underneath the adherent bacteria, in infected HeLa cells (Fig. 5).
*TccP4 is crucial for E. albertii 1551-2 to polymerize F-actin during AE lesion formation in infected Nck-null MEF cells. Fifty MEF cells, infected with wild-type, mutant, or complemented strains, were used to determine the number of adherent bacteria (A), the number of pedestals (B), and the percentage of adherent bacteria that formed pedestals (C). Note that the mutation in the tccP4 gene did not affect the ability of the double mutant 1551-2ΔtccP3/ΔtccP4 to adhere to the host cells. However, it directly affected its ability to trigger F-actin recruitment, which is necessary for the formation of pedestals characteristic of the AE lesion. ns = non-significant, ***P < 0.0001.
Colocalization of the recombinant TccP4-Myc protein with polymerized F-actin underneath adhered bacteria in infected HeLa cells. The co-localization of TccP4-Myc recombinant protein with polymerized F-actin, in infected epithelial HeLa cells, is indicated by arrows. Note that the TccP4-Myc recombinant protein was produced only by 1551-2ΔtccP3/ΔtccP4(pTccP4) complemented strain (E). Scale bar: 10 µm.
TccP4 acts in F-actin recruitment in Nck-null cells infected with the EHEC O157:H7 strain EDL933
To test the hypothesis that TccP4 could replace TccP1 in F-actin recruitment for AE lesion formation in MEF Nck-null cells infected with the EHEC O157:H7 EDL933 strain, we constructed a tccP/espF_U_ mutant strain (termed EDL933ΔtccP) and further transformed the mutant strain with the pKC471 or pTccP4 plasmids, the former of which carry the tccP1 gene. The wild-type EDL933 as well as the derived strains exhibited similar growth rates, as evidenced by the bacterial growth curves (Fig. S2B; supplemental material is found at DOI 10.6084/m9.figshare.28152620).
As expected, we observed that the deletion of the tccP1 gene abolished the ability of the EDL933ΔtccP1 mutant strain to induce AE lesion formation in infected MEF Nck-null cells. Moreover, this phenotype was restored when the EDL933ΔtccP1 mutant was transformed with pKC471 as well as with pTccP4 (Fig. S5; supplemental material is found at DOI 10.6084/m9.figshare.28152620).
The tccP4 gene is the most frequent tccP-encoding gene subtype present in E. albertii strains
An in silico screening for the presence of genes encoding the distinct TccP subtypes (tccP1, tccP2, tccP3, and tccP4) in a collection of 637 sequenced E. albertii strains revealed that the tccP4 was the most frequent subtype detected (71.0%), followed by tccP1 (61.9%), tccP2 (15.2%), and tccP3 (3.5%) (Table 2), while in 65 E. albertii strains (10.2%) no TccP-encoding gene was detected (Table S2; supplemental material is found at DOI 10.6084/m9.figshare.28152620). In addition, 330 E. albertii strains (51.8%) harbored two or three tccP subtypes concomitantly (Table S2; supplemental material is found at DOI 10.6084/m9.figshare.28152620), with the most common combination being tccP1 +tccP4 (35.9%). Curiously, tccP4 was not found in any of the genomes of other species of the Enterobacteriaceae family investigated, including E. coli (data not shown).
The phylogenetic analysis of the 637 E. albertii genomes used in this study confirmed the existence of two major clades, referred to as clade 1 and clade 2, in the E. albertii population. Furthermore, Bayesian Analysis of Population Structure (BAPS) subdivided the E. albertii strains analyzed in 9 BAPS clusters (C1–C9), with 8 of them belonging to clade 1 (C1–C8) and one (C9) to clade 2 (Fig. 6; Fig. S6; supplemental material is found at DOI 10.6084/m9.figshare.28152620). The distribution of tccP genes in the E. albertii genomes analyzed revealed that tccP1, tccP2, and tccP3 were distributed in limited BAPS clusters, while tccP4 was found in all BAPS clusters identified (Table 2; Fig. 6). Notably, four E. albertii strains (ESC_QA4137AA, ESC_KB0840AA, ESC_SA4787AA, and ESC_FB7444AA) that were separated early from other strains harbored only the tccP4 gene (Fig. 6; Fig. S6; supplemental material is found at DOI 10.6084/m9.figshare.28152620). This result, together with the wide distribution of the tccP4 gene, suggests that E. albertii acquired tccP4 at the very early stage of E. albertii diversification.
Phylogenetic relationships of the 637 genome-sequenced E. albertii strains analyzed in this study and the distribution of tccP genes in these strains. A maximum likelihood tree was constructed based on the 93,457 SNPs identified in the core genes ( = 1,879) of the 637 E. albertii genomes analyzed. The tree was rooted by E. coli K-12 strain MG1655 and strain names are indicated at each tip. BAPS clusters and the presence/absence of tccP1, tccP2, tccP3, and tccP4 genes are shown in the outer circles. An asterisk indicates the innermost circle, where strains used for genetic structure comparison of tccP4-containing chromosome regions (shown in Fig. 7) are highlighted. In the innermost circle, strains for which only draft genome sequences were available are indicated in blue, while strains with the complete genome are indicated in red. Of importance, tccP4 was the most frequent tccP subtype detected and found in all BAPS clusters.
The tccP4-containing chromosome region possesses a highly variable genetic organization
To gain better insights into how E. albertii strains acquired tccP4, we performed the structural comparison of the tccP4-containing genomic regions of 16 tccP4-positive and the analogous genomic regions of five tccP4-negative strains, which were selected from different BAPS clusters. In this analysis, we observed that in all E. albertii strains evaluated the tccP4 was present on the chromosome and close to the tRNA-thrW gene, located between the proAB and asrABC operons (Fig. 7). Note that the lengths of tccP4 genes remarkably differed among the E. albertii strains due to the difference in the numbers of PRRs, ranging from one to six repeats. Moreover, we observed that the genetic organization of this region was highly variable among the E. albertii strains analyzed.
Genetic structure comparison of the tccP4-containing chromosome regions of 16 tccP4-positive E. albertii strains and the analogous regions of five tccP4-negative strains. BAPS clusters of each strain (C1–C9) are indicated after the strain name. tccP4-positive strains are indicated in bold, and strains, for which only draft genome sequences were available, are indicated by parenthesis. Note that in all E. albertii strains analyzed tccP4 was present on the chromosome and close to the tRNA-thrW gene located between the proAB and asrABC operons. The genetic organization of this region was highly variable among the E. albertii strains evaluated.
A variety of prophages and integrative elements, or traces of their integration, were observed integrated into the tRNA-thrW gene, even in some tccP4-negative strains. Of importance, the tccP4 gene does not appear to be located on these mobile genetic elements (Fig. 7; Fig. S7; supplemental material is found at DOI 10.6084/m9.figshare.28152620). In addition, the genetic structure of this region has been more complicated in some E. albertii strains due to the insertion of several types of insertion sequences (ISs) and local genomic inversions. In the face of these findings, it was not possible to predict the ancestral genetic structure of this region and identify the genetic event involved in the acquisition of the tccP4 gene.
DISCUSSION
The AE lesion formation on the surface of epithelial cells infected by E. albertii strains is the main virulence strategy used by this pathogen to cause diarrheal disease in the human host (5, 7). The polymerization of F-actin, underneath adherent bacteria, is a key step in the AE lesion formation and involves the recruitment and activation of several eukaryotic components. Tir protein from EHEC of serotype O157:H7 lacks the equivalent of EPEC TirY_474_, and, consequently, does not recruit the eukaryotic protein Nck to polymerize F-actin for the AE lesion formation, a fact that intrigued researchers trying to understand the bacterial and eukaryotic proteins required for AE lesion formation induced by EHEC O157:H7. This question began to be answered when two independent research groups identified a bacterial protein, translocated to the eukaryotic cells by T3SS and termed TccP/EspF_U_ (referred to as TccP1 in this study), which is necessary for F-actin polymerization during the AE lesion formation by EHEC O157:H7 in a scenario independent of Nck (17, 20). After the discovery of TccP/EspF_U_ (TccP1), two additional subtypes of this adaptor protein— TccP2 and TccP3—were identified in the typical EPEC strain B171 and the E. albertii strain 1551-2, respectively (23, 24). The observation that the E. albertii 1551-2 strain can recruit F-actin in a scenario lacking Nck, and independently of TccP3, motivated us to further explore the potential for novel TccP subtypes in the 1551-2 strain, which culminated in the identification of TccP4.
The proline-rich effector TccP/EspF_U_ activates the eukaryotic actin nucleation-promoting factor N-WASP but does not bind Tir directly (31, 32). Further studies identified that the IRTKS eukaryotic protein binds to the TccP/EspF_U_ effector via its SH3 domain, while the IRSp53/MIM homology domain (IMD) of the IRTKS binds to the motif NPY_458_ of the C-terminal portion of Tir, thus bridging Tir and the adaptor effector TccP/EspF_U_ (21). TccP/EspF_U_ can bind to eukaryotic proteins, such as IRTKS and N-WASP, through specific motifs present in PRRs, leading to the recruitment of F-actin and subsequent AE lesion formation on the surface of infected epithelial cells (15). Progressive deletions within TccP/EspF_U_ revealed that the presence of the N-terminal translocation signal and at least two PRRs are the minimum requirements necessary for triggering F-actin polymerization in EHEC-infected epithelial cells (32). Drawing a parallel with what was described above, the observation that TccP4 harbored an N-terminal type 3 secretion system (T3SS) signal sequence and three PRRs led us to hypothesize that this effector could be functional after being translocated to the infected epithelial cells, a fact that was evidenced through the experimental approaches employed in this study.
The first 20 amino acids of the PRR domain comprise the portion of the EHEC O157:H7 TccP/EspF_U_ effector that binds to the autoinhibitory GTPase binding domain (GBD) in the N-WASP proteins, leading to its activation (33). Comparing the first 20 amino acids of the PRR1 of TccP4 and the PRR5 of the EHEC O157:H7 TccP/EspF_U_, we observed that they differ by only two amino acids. Taking the PRR5 of TccP/EspF_U_ as a reference, we observed the substitution of methionine (M) at position 9 by leucine (L), both with hydrophobic side chains; and the substitution of alanine (A), an amino acid with a hydrophobic side chain, at position 20 by glutamic acid (E), an amino acid with a negatively charged side chain. Considering that TccP4 was crucial for F-actin polymerization in Nck-null MEF cells infected by E. albertii 1551-2, these amino acid substitutions do not affect TccP4 functionality.
If, on the one hand, the N-terminal portion of TccP/EspF_U_ activates N-WASP (32, 33), the C-terminal portion binds to the IRTKS (26), acting as a link between these two eukaryotic proteins in the F-actin recruitment necessary for the AE lesion formation in infected epithelial cells. The interaction of TccP/EspF_U_ with the SH3 domain of the IRTKS/IRSp53 involves two adjacent PxxP motifs present in the C-terminal portion of the TccP/EspF_U_ adaptor protein, thus explaining the high affinity and selectivity of this interaction (26). These two PxxP motifs are conserved in the C-terminal portion of the three PRRs of the TccP4 protein, thus suggesting that, like TccP/EspF_U_ of EHEC O157:H7, TccP4 has a high affinity to bind to IRTKS/IRSp53 in the F-actin recruitment cascade. This comparative scenario with TccP4 (from the 1551-2 strain) and TccP/EspF_U_ (from EHEC O157:H7) indicates that both adaptor proteins act similarly in the F-actin recruitment process for the AE lesion formation.
The collective analysis of the data from the present study and previous publications from our laboratories (24, 25) allowed us to conclude that the E. albertii 1551-2 strain can polymerize F-actin using the Nck-dependent, as well as the Nck-independent pathways. The simultaneous use of both pathways for F-actin polymerization was first identified in a typical EPEC strain of serotype O119:H6 (34) and later observed in several typical and atypical EPEC strains of distinct serotypes (35–37). However, to date, studies with in vivo experimental models have not demonstrated that the coexistence of two distinct pathways for F-actin polymerization, during AE lesion formation, would result in bacteria that are more pathogenic for the host (38). However, we can hypothesize that the coexistence of two distinct pathways involved in the establishment of the AE lesion may represent a way to persevere this virulence characteristic in cases where the bacteria undergo mutations and/or genetic rearrangements that culminate in the modification and/or loss of genes responsible for encoding proteins involved in the establishment of this phenotype by one of the pathways.
Our analysis of the phylogenetic relationship of 637 genome-sequenced E. albertii strains and the distribution of the tccP genes in these strains provided interesting findings on the process of tccP4 acquisition by E. albertii. Although several tccP subtypes were present in many E. albertii strains, the tccP4 gene was the most frequent and observed in all E. albertii lineages identified (BAPS clusters C1-C9). Interestingly, the tccP4 gene was detected in E. albertii strains that were first separated from other E. albertii. These findings added to the evidence that tccP4 is located at the same chromosome locus in the E. albertii strains analyzed, suggesting that this gene was acquired at a very early stage during the diversification of E. albertii from its ancestor. In this regard, it is intriguing to note that the tccP4 gene, or its close homolog, has not been found in E. coli strains, as well as in other bacteria of the Enterobacteriaceae family sequenced so far.
The highly variable genetic organization of the tccP4-containing regions and the presence of several mobile genetic elements in this region were also important findings, because although the genetic event involved in the tccP4 acquisition by E. albertii was not predicted due to this variation, repeated genomic deletion mediated by these genetic elements may explain the lack of tccP4 in E. albertii strains.
In conclusion, we identified a novel TccP subtype—termed in the present study as TccP4—and demonstrated that this protein is required for the recruitment of F-actin during the AE lesion formation in infected cells by the E. albertii 1551-2 strain. Furthermore, the gene encoding this TccP subtype unique to E. albertii is widely distributed in this species and suggests that the tccP4 gene was acquired at a very early stage during the diversification process of E. albertii.
MATERIALS AND METHODS
Bacterial strains, plasmids, and genomes used in this study
The E. albertii 1551-2 strain was isolated from a diarrheic child at 23 months of age, during an epidemiological study performed at the Federal University of São Paulo (UNIFESP)—Brazil, in 1989 (27, 28, 39). All bacterial strains and plasmids used in the present study are listed in Table 1. The strains were routinely grown in Lysogeny Broth (LB) at 37°C and kept in LB supplemented with 30% glycerol at −80°C.
For in silico analysis, a total of 637 genome sequences of E. albertii strains registered in the EnteroBase website v1.2.0 (https://enterobase.warwick.ac.uk) (40) were used, after excluding 38 strains from the 675 strains registered as E. albertii (accessed on the 1st of May 2024) due to the lack of genome sequence information, low completeness (<95%) or high contamination (>5%) as estimated by CheckM (41). The final set of E. albertii strains used for in silico analysis is shown in Table S3 (Supplemental material is found at DOI 10.6084/m9.figshare.28152620).
Phylogenetic analysis and amino acid sequence comparison of TccP4 with known TccP subtypes
The genome sequence of strain 1551-2 (accession number CP025317.1) was searched by tblastn using the amino acid sequences of TccP family proteins (TccP1, TccP2, and TccP3) listed in Table S4 as queries (Supplemental material is found at DOI 10.6084/m9.figshare.28152620). Coding sequences of TccP subtypes were manually annotated with GENETYX-MAC Version 16.0.9 (GENETYX Corp., Japan). The criterion of screening and subtyping was >97% amino acid sequence identity to the N-terminal 56 amino-acids sequence of any of the known TccP family proteins.
A maximum likelihood tree was built using the JTT matrix-based model (42) and analyzed using the MEGA11 software (43) to compare the amino acid sequence of the TccP4 effector from the 1551-2 strain (accession number: AUS64398.1) with those of other TccP subtypes previously described (Table S5; supplemental material is found at DOI 10.6084/m9.figshare.28152620). Subsequently, six representative amino acid sequences of the distinct TccP subtypes were used to be aligned using the ClustalW tool in MEGA 11 software. Percentage sequence identities between TccP subtypes were determined using blastp (https://blast.ncbi.nlm.nih.gov/Blast.cgi?PAGE=Proteins). The predicted molecular weight of the TccP4 protein was calculated using Expasy (https://web.expasy.org/cgi-bin/compute_pi/pi_tool).
Deletion mutant construction and in trans complementation
A tccP4 deletion mutant strain was constructed by the one-step allelic exchange recombination method (44), using the thermosensitive plasmid pKOBEG-Apra^R^, which carries lambda Red recombination genes (30). First, the 1551-2ΔtccP3 strain (24) was transformed by electroporation with the pKOBEG-Apra^R^, and strains carrying this plasmid were selected on LB agar containing 100 µg/mL of apramycin after incubation for approximately 18 hours at 30°C.
Then, a DNA fragment harboring the gene encoding zeocin resistance (ble) flanked by 50 base pairs (bp) of the 5′ and 3′ ends of tccP4 was prepared by polymerase chain reaction (PCR), using the primers tccP4-zeo-F and tccP4-zeo-R and the genomic DNA of the 1711-4ΔfliC mutant strain (29) as a template for the ble gene amplification. All primer sequences and conditions used in the PCR for DNA amplification are described in Table 3. Next, this DNA fragment was electroporated into strain 1551-2ΔtccP3, containing pKOBEG, and the recombinant bacteria were selected on LB low salt agar plates supplemented with 60 µg/mL of zeocin. To obtain tccP4-deletion mutants, bacteria recovered from zeocin-containing LB plates were screened for the loss of tccP4 and the presence of ble using the set of primers tccP4-F/tccP4-R and zeo-F/zeo-R, respectively (Table 3).
Primers flanking the tccP4 gene were then designed (tccP4-flank-F and tccP4-flank-R) (Table 3) and used to amplify a DNA fragment that was subsequently sequenced at the Biotechnology Institute/UNESP/Botucatu (https://www.ibtec.unesp.br/#!/servicos68/sequenciamento-sanger/) to confirm the correct insertion of the ble gene replacing tccP4. Once the loss of tccP4, pKOBEG was eliminated by growing the double mutant strain (1551-2ΔtccP3/tccP4) at 42°C in the presence of 60 µg/mL of zeocin.
To complement the double mutant 1551-2ΔtccP3/tccP4 with the tccP4 gene, a DNA fragment, from 100 bp upstream of the ATG start codon until just before the tccP4 ORF stop codon, was amplified with primers tccP4-XbaI-F and tccP4-BamHI-R (Table 3) and cloned into XbaI and BamHI sites of the pKC471 plasmid (Table 1), thus creating the recombinant plasmid termed pTccP4. This recombinant plasmid (pTccP4) was electroporated into the double mutant to generate the complemented 1551-2ΔtccP3/tccP4 (pTccP4) strain (Table 1).
Growth curves were constructed with at least three independent experiments for 6 hours with the E. albertii 1551-2 strain and its respective mutants and the tccP4 complemented strain. The strains were grown in 3 mL of LB broth for 18 h at 37°C, and subsequently transferred to 25 mL of LB at a 1:50 dilution, followed by incubation at 37°C under constant shaking (250 rpm). Aliquots were analyzed every 30 min for optical density (OD) reading under λ = 600 nm in an ELISA reader (BioTek, USA). Two-way ANOVA was used for statistical analysis, and P values ≤ 0.05 were considered statistically significant.
Immunoblotting for detection of the TccP4-Myc recombinant protein
The bacterial strains were grown in 3 mL of LB medium at 37°C for 18 hours. The cultures were centrifuged at 3,500 × g for 15 minutes to harvest the bacteria. The supernatant was discarded, and the resulting pellet was resuspended in 200 µL of phosphate-buffered saline (PBS). The preparations were boiled at 95°C for 5 minutes, according to the methodology described by Laemmli (46). Then, 20 µL of the preparations were mixed with 20 µL of sample buffer for sodium dodecyl sulfate-polyacrylamide gel electrophoresis (SDS-PAGE) and boiled again to denature the proteins. Approximately 15 µL of each preparation was added to a 12% SDS-polyacrylamide gel and proteins were separated by electrophoresis at 110 V until the tracking dye in the sample buffer reached the end of the gel. Subsequently, the proteins were transferred to nitrocellulose membranes at 10 V for 45 min and blocked with 5% non-fat milk in PBS for approximately 18 hours (47). The membranes were probed first with 1:5,000-diluted mouse monoclonal anti-Myc antibody (clone 9E40 sc-40, Santa Cruz Biotechnology, USA), and then, with 1:4,000-diluted anti-mouse secondary antibody (Sigma-Aldrich, USA) conjugated with peroxidase, diluted 1:4,000. Both antibodies were diluted in PBS containing 5% non-fat milk and 0.05% Tween-20 (LGC Biotecnologia, Brazil). Finally, the protein bands were detected by West Pico PLUS Chemiluminescent Substrate (Thermo Fisher Scientific, USA) and visualized on the Amersham Imager 600 (GE Healthcare, USA) transilluminator.
F-Actin pedestals quantification
HeLa and Nck-null MEF cells were routinely cultivated in Dulbecco’s modified Eagle medium (DMEM) (Sigma, USA) supplemented with 10% fetal bovine serum (FBS) (Gibco, USA) and 1% antibiotic mixture (penicillin—10,000 U/mL and streptomycin—10 mg/mL, ThermoFisher, USA) in an atmosphere of 5% CO_2_ at 37°C.
For the fluorescent actin staining (FAS) assay, 1 mL of a cell suspension, prepared with DMEM supplemented with 2% FBS and containing 1 × 10^5^ HeLa cells and 4 × 10^4^ Nck-null MEF cells (48), was added to each well of a 24-well microplate (TPP, Switzerland). Then, the wild-type E. albertii 1551-2 strain, and the mutant and complemented strains were grown overnight in LB at 37°C, and a volume of 20 µL of a bacterial culture containing approximately 10^7^ CFU (colony-forming units) was added to the cell preparations and incubated for a total period of 6 hours, with a washing step after 3 hours and replacement of the culture medium, in a 5% CO_2_ atmosphere. The coverslips were washed six times with PBS and fixed with 3% formaldehyde for 24 hours. After, the wells were washed twice with PBS, permeabilized with 1% Triton X-100 (INLAB, Brazil) in PBS, and incubated with a solution containing FITC-phalloidin (Life Technologies, USA) (diluted at 1:125) and DAPI 4′,6-diamidino-2-phenylindole dihydrochloride (Life Technologies, USA) (1:500), prepared in PBS. The coverslips were mounted and observed with the Olympus BX60 fluorescence microscope under an immersion oil objective (100×).
For quantification of the number of F-actin pedestals present in the Nck-null cells infected with the wild-type, mutants, and complemented strains, 50 Nck-null MEF cells infected with each of the tested strains were randomly selected, and the number of bacteria, as well as the number of F-actin pedestals, were counted. The statistical analyses were performed with GraphPad Prism Version 8.0.1. One-way ANOVA was used for the parametric data, followed by the post hoc Tukey HSD test. Kruskal-Wallis was used for non-parametric data followed by the post hoc Dunn. P values ≤ 0.05 were considered statistically significant.
Immunofluorescence to demonstrate the colocalization of TccP4 and polymerized F-actin
Hela cells were infected with the strains 1551-2 and 1551-2ΔtccP3/ΔtccP4 (pTccP4) and incubated for 6 hours in an atmosphere of 5% CO_2_. Then, the cells were washed three times with PBS, fixed with 3% formaldehyde, permeabilized with 1% Triton X-100 in PBS for 5 min, and blocked with PBS supplemented with 2% bovine serum albumin (BSA) for 1 hour.
For detection of the TccP4-Myc recombinant protein, the infected cells were probed with mouse monoclonal anti-Myc diluted 1:100 in PBS (clone 9E40 sc-40, Santa Cruz, Biotechnology, EUA) for 1 hour. After two washes with PBS, the preparations were probed with Alexa Fluor 568-goat anti-mouse secondary antibody (Molecular Probes) diluted 1:1,000 in PBS. Subsequently, polymerized F-actin and DNA were stained with FITC-phalloidin (1:125) (Life Technologies, Carlsbad, EUA) and DAPI (1:500) (Life Technologies, Carlsbad, EUA) in PBS, respectively. The preparations were visualized with an Olympus BX60 fluorescence microscope under oil immersion.
Mutant construction and in trans complementation in EDL933 strain
The EDL933ΔtccP mutant strain was constructed by the one-step allelic exchange recombination method (41), as described above, using the primers tccP/EHEC-zeo-F and tccP/EHEC-zeo-R (Table 3). The pKC471 and pTccP4 plasmids (Table 1) were introduced in the EDL933ΔtccP mutant strain by electroporation, generating the EDL933ΔtccP(pKC471) and EDL933ΔtccP(pTccP4) strains, respectively (Table 1). The growth rate of wild-type EDL933 and derivative strains were measured using growth curves performed as previously described.
Identification and subtyping of tccP in E. albertii-sequenced genomes
To identify and subtype the TccP-encoding gene(s) present in the 637 genome-sequenced E. albertii strains, we performed tblastn using amino acid sequences of the representative TccP subtype as queries, as indicated in Table S4 (Supplemental material is found at DOI 10.6084/m9.figshare.28152620). The criterion for classifying TccP sequences as belonging to one of the four distinct subtypes was >97% amino acid sequence identity with the N-terminal 56 amino acid sequences of representative TccP family proteins used as queries.
Search of tccP4 in E. coli and other strains from the Enterobacteriaceae family
To determine whether TccP4 or its close homolog is encoded by E. coli and other strains from the Enterobacteriaceae family, tblastn analysis was performed on microbial genome blast (https://blast.ncbi.nlm.nih.gov/Blast.cgi?PROGRAM=tblastn&PAGE_TYPE=BlastSearch) using the amino acid sequence of TccP4 from 1551 to 2 strain as a query.
Analysis of tccP4-containing regions
In 16 tccP4-positive and five tccP4-negative E. albertii genomes, the tccP4-containing regions in the tccP4-positive genomes and analogous regions in the tccP4-negative genomes were compared. Gene annotation was carried out with DFAST and manually curated using the in silico Molecular Cloning Genomics Edition software (IMC-GE ver. 8.61; In Silico Biology, Inc., Yokohama, Japan). Sequence comparison of the regions and dot-plot analysis were performed by blastn using the “CompareSequences” function of the GenomeMatcher software (49).
Phylogenetic analysis of genome-sequenced E. albertii strains
Core gene SNP-based maximum likelihood (ML) tree of the 637 E. albertii strains was constructed as follows: the genome assemblies of a total of 637 strains (including strain 1551-2) were annotated using DFAST, and core genes (n = 1,879) were identified using Roary v3.13,0 (50) with a 90% amino acid sequence identity cut-off. Core gene SNPs (n = 93,457) were extracted using the core gene alignment tool in Roary and used as inputs for ML inference with RAxML v8 (51) and displayed and annotated using iTOL v4 (https://itol.embl.de) (52). Identical E. albertii genomes showing no SNPs were deduplicated (excluded strains were indicated in Table S3; supplemental material is found at DOI 10.6084/m9.figshare.28152620). BAPS clusters were identified using RhierBAPS (53).
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Huys G, Cnockaert M, Janda JM, Swings J. 2003. Escherichia albertii sp. nov., a diarrhoeagenic species isolated from stool specimens of Bangladeshi children. Int J Syst Evol Microbiol 53:807–810. doi:10.1099/ijs.0.02475-012807204 · doi ↗ · pubmed ↗
- 2Ooka T, Seto K, Kawano K, Kobayashi H, Etoh Y, Ichihara S, Kaneko A, Isobe J, Yamaguchi K, Horikawa K, Gomes TAT, Linden A, Bardiau M, Mainil JG, Beutin L, Ogura Y, Hayashi T. 2012. Clinical significance of Escherichia albertii. Emerg Infect Dis 18:488–492. doi:10.3201/eid 1803.11140122377117 PMC 3309589 · doi ↗ · pubmed ↗
- 3Ooka T, Tokuoka E, Furukawa M, Nagamura T, Ogura Y, Arisawa K, Harada S, Hayashi T. 2013. Human gastroenteritis outbreak associated with Escherichia albertii, Japan. Emerg Infect Dis 19:144–146. doi:10.3201/eid 1901.12064623260717 PMC 3557987 · doi ↗ · pubmed ↗
- 4Gomes TAT, Ooka T, Hernandes RT, Yamamoto D, Hayashi T. 2020. Escherichia albertii pathogenesis. Eco Sal Plus 9. doi:10.1128/ecosalplus.esp-0015-2019 PMC 1116857632588811 · doi ↗ · pubmed ↗
- 5Albert MJ, Alam K, Islam M, Montanaro J, Rahaman AS, Haider K, Hossain MA, Kibriya AK, Tzipori S. 1991. Hafnia alvei, a probable cause of diarrhea in humans. Infect Immun 59:1507–1513. doi:10.1128/iai.59.4.1507-1513.19912004829 PMC 257869 · doi ↗ · pubmed ↗
- 6Albert MJ, Faruque SM, Ansaruzzaman M, Islam MM, Haider K, Alam K, Kabir I, Robins-Browne R. 1992. Sharing of virulence-associated properties at the phenotypic and genetic levels between enteropathogenic Escherichia coli and Hafnia alvei. J Med Microbiol 37:310–314. doi:10.1099/00222615-37-5-3101433251 · doi ↗ · pubmed ↗
- 7Yamamoto D, Hernandes RT, Liberatore AMA, Abe CM, Souza R de, Romão FT, Sperandio V, Koh IH, Gomes TAT. 2017. Escherichia albertii, a novel human enteropathogen, colonizes rat enterocytes and translocates to extra-intestinal sites. P Lo S One 12:e 0171385. doi:10.1371/journal.pone.017138528178312 PMC 5298312 · doi ↗ · pubmed ↗
- 8Lima MP, Yamamoto D, Santos A de M, Ooka T, Hernandes RT, Vieira MAM, Santos FF, Silva RM, Hayashi T, Gomes TAT. 2019. Phenotypic characterization and virulence-related properties of Escherichia albertii strains isolated from children with diarrhea in Brazil. Pathog Dis 77:ftz 014. doi:10.1093/femspd/ftz 01430865776 · doi ↗ · pubmed ↗