BIOHAZ scientific opinion on the equivalence of an alternative heat treatment process of feathers and down
Ana Allende, Avelino Alvarez‐Ordóñez, Valeria Bortolaia, Sara Bover‐Cid, Wietske Dohmen, Laurent Guillier, Lieve Herman, Liesbeth Jacxsens, Lapo Mughini‐Gras, Maarten Nauta, Jakob Ottoson, Luisa Peixe, Fernando Perez‐Rodriguez, Panagiotis Skandamis, Elisabetta Suffredini

TL;DR
The paper evaluates if heating feathers and down at 100°C for 30 minutes is as effective as a standard method in killing pathogens.
Contribution
The study assesses the equivalence of a new heat treatment for feathers and down against a regulatory standard.
Findings
No evidence supports a 5 log10 reduction of Enterococcus faecalis with the proposed method.
Limited data prevents confirming a 5 log10 reduction of Salmonella Senftenberg.
Only one study on viruses showed a 1 log10 reduction, insufficient for regulatory equivalence.
Abstract
The European Commission requested EFSA to provide a scientific opinion on the equivalence between the heat treatment process of feathers and down with dry heat to a temperature of 100°C for 30 min and the treatment set up in Commission Regulation (EU) No 142/2011, in terms of inactivation of relevant pathogens. To be considered at least equivalent to the methods in the legislation, the alternative method should be able to reduce the concentration of Enterococcus faecalis or Salmonella Senftenberg by at least 5 log10 and the infectious titre of Anelloviridae and Circoviridae by at least 3 log10. An extensive literature search (ELS) was conducted to identify studies in which the log10 reduction or the D value of the indicators were determined after dry heating in matrices with low moisture/water activity. The ELS did not provide any study on the inactivation of E. faecalis by dry heating.…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
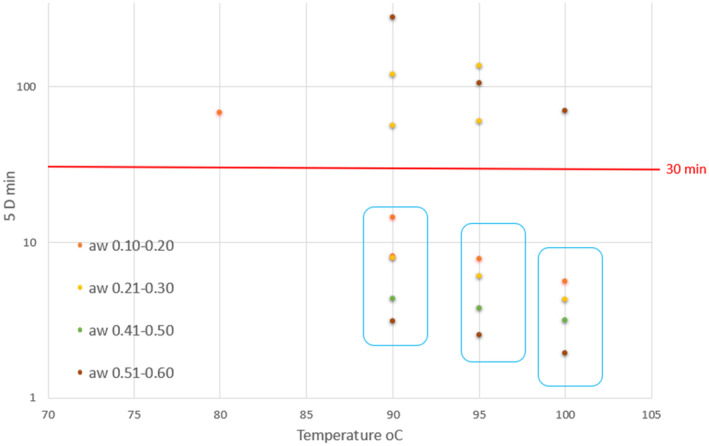 FIGURE 1
FIGURE 1| Product/matrix | Treatment | Temp (°C) | Time (min) | log10 reduction |
| Reference |
|---|---|---|---|---|---|---|
| Whole raw almond kernels | Wet heat (hot water) | 88 | 0.5 | 3.5 | 0.36 | Harris et al. (2012) |
| Homogenised defrozen mussels | Wet heat (cooking) | 70 | – | – | 0.5–1 | Riva et al. (1991) |
| Ground pork meat | Wet heat | 95 | – | – | 9.46 | Mroz et al. (1985) |
| Ground pork meat | Wet heat | 90 | – | – | 13.91 | |
| Milk | Wet heat | 68 | – | – | 4.19 | Roman (2003) |
| Ground beef | Wet heat | 70 | – | – | 0.15 | Saucier and Plamondon (2011) |
| Product/matrix | aw/moisture % | Temp (°C) | Time (min) | log10 reduction |
| Reference |
|---|---|---|---|---|---|---|
| Turkey litter | 0.839 | 75 | 51 | 3.2 | – | Chen and Jiang (2017) |
| Turkey litter | 0.839 | 85 | 49.2 | 4.73 | – | |
| Broiler litter | 0.808 | 75 | 57 | 3.43 | – | |
| Broiler litter | 0.808 | 85 | 52.8 | 3.79 | – | |
| Pet food (adult small breed dog food) | 0.33 | 70 | 25 | 1.65 ± 0.17 | 12.59 | Dhaliwal et al. (2021) |
| Skim milk powder | 0.33 | 70 | 25 | 1.75 ± 0.06 | 14.96 | |
| Carbohydrate‐based burger binder | 0.33 | 70 | 25 | 2.43 ± 0.15 | 0.53 | |
| Liquid whole eggs | – | 64 | – | – | 2.8 | Doyle and Mazzotta (2000) |
| Ground turkey | – | 65 | – | – | 3.4 | |
| Ground beef | – | 63 | – | – | 2.08 | |
| Ground beef | – | 68 | – | – | 0.22 | |
| Molten chocolate | – | 90 | – | – | 36 | |
| Homogenised oysters | – | 70 | – | – | 0.3 | |
| Dried growth medium | 0.56–0.58 | 100 | 60 | 4.30 ± 0.00 | 14.0 | Furtado et al. (2023) |
| Dried growth medium | 0.56–0.58 | 95 | 60 | 2.82 ± 0.45 | 21.3 | |
| Dried growth medium | 0.56–0.58 | 90 | 90 | 1.60 ± 0.35 | 56.3 | |
| Hazelnuts shells | 4% | 100 | 30 | 3.03 ± 0.26 | 9.90 | Izurieta and Komitopoulou (2012) |
| Cocoa shells | 4% | 100 | 30 | ≥ 4.47 | < 6.71 | |
| Milk powder | 0.11 | 100 | 3 | – | 1.12 | Lang et al. (2017) |
| Milk powder | 0.25 | 100 | 3 | – | 0.86 | |
| Milk powder | 0.44 | 100 | 3 | – | 0.63 | |
| Milk powder | 0.58 | 100 | 3 | – | 0.39 | |
| Milk powder | 0.11 | 95 | 3 | – | 1.57 | |
| Milk powder | 0.25 | 95 | 3 | – | 1.21 | |
| Milk powder | 0.44 | 95 | 3 | – | 0.75 | |
| Milk powder | 0.58 | 95 | 3 | – | 0.51 | |
| Milk powder | 0.11 | 90 | 3 | – | 1.62 | |
| Milk powder | 0.25 | 90 | 3 | – | 1.58 | |
| Milk powder | 0.44 | 90 | 3 | – | 0.87 | |
| Milk powder | 0.58 | 90 | 3 | – | 0.62 | |
| Non‐fat milk powder | 0.199 | 90 | 14 | 4.8 ± 0.3 | 2.92 | Sekhon et al. (2020) |
| Non‐fat milk powder | 0.199 | 80 | 60 | 4.4 ± 0.1 | 13.64 | |
| Raw shelled almonds |
0.54–0.22 4.27% | 95 | 15 | 1.24 | 12.10 | Song and Kang (2021) |
| Raw shelled almonds |
0.54–0.22 4.27% | 95 | 60 | 2.20 | 27.27 | |
| Raw shelled almonds |
0.54–0.22 4.27% | 90 | 15 | 1.33 | 11.28 | |
| Raw shelled almonds |
0.54–0.22 4.27% | 90 | 60 | 2.49 | 24.10 | |
| Turkey litter | 20% | 75 | 75 | 4.2 | – | Wang et al. (2021) |
| Turkey litter |
0.925 30% | 75 | 92 | > 5 | – | |
| Laying hen litter composts | 15% | 75 | 60 | > 5 | – | |
| Aged broiler litter | 20% | 75 | 90 | 4.2 | – | Wang et al. (2018) |
| Aged broiler litter | 20% | 85 | 100 | > 5.5 | – | |
| Basil leaves |
0.195 2.48% | 80 | 60 | > 6.2 | – | Zhou et al. (2020) |
| Basil leaves |
0.557 8.6% | 100 | 40 | > 6.2 | – |
| Virus | Matrix/substrate | Initial load | Treatment | Temp (°C) | Time (min) | Level of inactivation | Reference |
|---|---|---|---|---|---|---|---|
| PCV | Virus suspension | 6.0 log10 TCID50/0.1 mL | – | 70 | 15 | No inactivation | Allan et al. (1994) |
| CAV | Virus suspension | – | Water bath | 100 | 15 | Total inactivationb | Goryo et al. (1985) |
| CAV | Virus suspension | 6.5 log10 TCID50/0.1 mL | – | 70 | 30 | Total inactivationa | Goryo et al. (1987) |
| CAV | – | – | – | 70 | 60 | No inactivation | Gowthaman (2019) |
| CAV | Virus suspension | 5.50 log10 TCID50/mL | – | 70 | 5 | No or limited inactivation | McNulty et al. (1989) |
| CAV | Virus suspension | 5.0/5.25 log10 TCID50/mL | Water bath | 70 | 5 | No or limited inactivation | McNulty et al. (1990) |
| CAV | Minced meat | 5.4 log10 TCID50/g | Heating in a stirred water bath | 100 | 30 | Total inactivationa (at least 5 log10) | Urlings et al. (1993) |
| CAV | Human factor VIII concentrate | Virus:product ratio 1:21.8 | Dry heat | 120 | 30 | 1 log10 | Welch et al. (2006) |
| PCV | Human factor VIII concentrate | Virus:product ratio 1:21.8 | Dry heat | 120 | 30 | 1 log10 | |
| CAV | Human factor VIII concentrate | Virus:product ratio 1:21.8 | Dry heat | 80 | 72 h | 0.75 log10 | |
| PCV | Human factor VIII concentrate | Virus:product ratio 1:21.8 | Dry heat | 80 | 72 h | 1.25 log10 | |
| PCV | Virus suspension | – | Wet heat | 60–80 | 3 | PCV capsids start to disassemble at 65°C | Wang et al. (2019) |
| Source of uncertainty | Cause of the uncertainty | Impact of the uncertainty on the conclusions |
|---|---|---|
| Inactivation data | Relevant references from which data on thermal reduction/inactivation of indicator microorganisms were extracted, were identified through literature searches using search strings, as described in the data and methodologies section. There is the possibility that some relevant reference was not identified or considered for data extraction. | This source can lead to an increase of the uncertainty associated to the answer to the AQs. |
| Inactivation data | The data on thermal reduction/inactivation of indicator microorganisms were extracted from experimental studies using different matrices from the one included in the mandate. Specific data from studies involving spiking in feathers and down with the indicator microorganisms of interest does not exist. The different composition in terms of dry matter (total solid contents, aw), fat content, etc. impacts on the capacity of bacteria and viruses to survive under different conditions of temperature and time. | This source can lead to an increase of the uncertainty associated to the answer to the AQs. |
| Inactivation data | The data on thermal reduction/inactivation of indicator microorganisms were extracted from experimental studies using particular strains/isolates of the relevant hazards and different analytical methods, which, for viral hazards, are not standardised. It is uncertain whether the tested strains are the most thermal resistant. In the case of viruses, a further source of uncertainty was the absence of specific data on duck circovirus, with the assessment relying only on data from porcine circoviruses, belonging to the same family. | These sources can lead to an increase of the uncertainty associated to the answer to the AQs. |
| Inactivation data | The lack of data or reports at the requested conditions of 100°C for 30 min. | This leads to increased uncertainty associated to the answer to the AQs. |
| Inactivation data | Lack of useful/ complete data for the viruses (no details on the initial load and/or the level of inactivation of the hazard) | This leads to increased uncertainty associated to the answer to the AQs. |
| Data on the process before the step of dry heating | The feathers and down pre‐treatments steps before the dry heating step were not assessed as were not part of this mandate. | The efficacy of dry heating to achieve the target reductions for selected indicators might be increased by previous steps in the process which were not assessed as were not part of this mandate. |
| Study design and sample preparation | The type of the study design and sample preparation may affect the log reduction and consequently the calculated | The log reduction could be over‐ or under‐estimated depending on the way the study is designed or the samples prepared. |
| Calculation of | For the graph related to the thermal inactivation of | The true |
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsInsect Utilization and Effects · Viral Infectious Diseases and Gene Expression in Insects · Meat and Animal Product Quality
SUMMARY
The Canadian Food Inspection Agency (CFIA) asked the Commission to verify whether or not a heat treatment consisting of dry heat to a temperature of 100°C or higher for at least 30 min can be considered equivalent to the one set out in Chapter VII, point C of Annex XIII to Regulation (EU) No 142/2011 in order to treat feathers and down to be exported to the European Union.
To address this request, the Commission requested EFSA to provide a scientific opinion concerning the equivalence of the heat treatment process of feathers and down consisting of dry heat to a temperature of 100°C or higher for at least 30 min, with the treatment set up in Commission Regulation (EU) No 142/2011, in terms of inactivation of relevant pathogens.
Over the years the approach followed by EFSA BIOHAZ for assessing the equivalence of alternative methods to the processing methods currently approved in the Commission Regulation (EU) No 142/2011 has been harmonised. To be considered at least equivalent to the processing methods approved in the legislation, the alternative methods for Category 3 animal by‐products (ABP) should be able of reducing the concentration of the relevant indicator bacteria by at least 5 log_10_ and the infectious titre of relevant viruses by at least 3 log_10_.
The BIOHAZ Panel determined that the bacterial and viral indicators identified for feathers and down in a previous EFSA Scientific Opinion (EFSA BIOHAZ Panel, 2021), which were selected based on the requirements outlined in point 1 of Section 2 of Chapter III of Annex V of Commission Regulation (EU) No 142/2011, were also considered for the current assessment. These indicators include the thermal resistant bacteria Enterococcus faecalis and Salmonella Senftenberg, as well as thermal resistant viruses from the Anelloviridae family (chicken anaemia virus (Gyrovirus chickenanemia)) and the Circoviridae family (duck circovirus (Circovirus duck)).
An extensive literature search (ELS) was conducted to identify relevant peer‐reviewed papers on the thermal inactivation of E. faecalis, S. Senftenberg, Anelloviridae (chicken anaemia virus) and Circoviridae (duck circovirus and porcine circovirus) at different time/temperature combinations by dry heat and/or in matrices with low water activity/moisture (even if different from those in the mandate). The experimental studies in which heat resistance parameters (i.e. D and/or z values) or number of log_10_ reduction of E. faecalis, S. Senftenberg or selected viruses were measured after thermal treatment in different matrices, similar (i.e. low moisture/low water activity) to the one included in the mandate, were further analysed. Overall, 46 papers were selected for data extraction: 35 for bacteria and 11 for viruses (chicken anaemia virus and porcine circovirus). No paper was retrieved for duck circovirus.
According to the Annex 2 submitted by the CFIA, before the heat treatment at 100°C for at least 30 min, the process of feather and down includes drying with natural heat on a stretched mesh frame in the sun outside or inside on a stretched mesh frame with a warm air flow at low temperature to drop the moisture content to around 20% followed by drying in an oven with temperature control to reduce the moisture content to around 10%. The moisture content was indicated only in a few papers retrieved through the ELS, while water activity (a_w_) was reported in almost all papers.
For E. faecalis, no articles were retrieved through the ELS describing the impact of dry heating on its inactivation. Based on the literature review, the 5 log_10_ reduction was obtained in wet heating conditions, where E. faecalis was rapidly inactivated even at temperatures < 100°C. Since there is evidence that low a_w_/dry heating conditions induce higher heat resistance of a wide range of microorganisms in comparison to wet heating, it cannot be assured that a 5 log_10_ reduction is achieved for E. faecalis in feathers and down with dry heating at 100°C for 30 min.
For S. Senftenberg, several studies retrieved through the ELS showed that the heat resistance increases when the a_w_ decreases, as typically occurs during dry heating. Several articles reporting data in matrices (always different from feathers and down) with a low a_w_ at time/temperature combinations close to 100°C for 30 min, provide log reductions below 5 log_10_ for dry heat treatments at 100°C for 30 min and even up to 90 min. For the datasets for which moisture values were reported to be lower or equal to 20%, and where the temperature was 75°C–100°C, the time for 5 log_10_ reduction always exceeded 30 min. Only 2 datasets reported log_10_ reductions at 100°C and 30 min for matrices with a very low moisture value (4%) and showed a 3.03 log_10_ reduction in hazelnut shells and > 4.47 log_10_ reduction in cocoa shells. Therefore, there is no clear evidence demonstrating that dry heat treatment at 100°C for 30 min can lead to a 5 log_10_ reduction of S. Senftenberg.
S. Senftenberg is generally considered the Salmonella serovar with the highest heat resistance in high moisture foods, with D values in different model systems around 10‐ to 20‐fold higher than those of other serovars, such as S. Typhimurium or S. Enteritidis. However, when dry heat is applied or the results refer to heat resistance in low moisture matrices, some of the articles retrieved through the ELS showed that other serotypes, including S. Enteritidis and S. Typhimurium, are more resistant than S. Senftenberg.
For viruses of the Anelloviridae and Circoviridae families (chicken anaemia virus and duck circovirus, respectively), there is also limited evidence regarding their survival/inactivation under dry heat treatments. Only a few experimental articles provide data on their heat inactivation, and most of them were performed in liquid viral suspensions (i.e. high a_w_) at temperatures well below 100°C. The only experimental study providing data on the inactivation of Anelloviridae and Circoviridae under dry heat treatment reported low inactivation, of approximately 1 log_10,_ after 30 min treatment at a temperature of 120°C. The scarcity data, combined with this finding, raises doubts about the ability of a dry heat treatment at 100°C for 30 min to achieve a 3 log_10_ reduction of Anelloviridae and Circoviridae in feathers and down.
Based on the data extracted from the ELS, the identified sources of uncertainty, and the expertise in the working group (WG), the following conclusions were achieved through expert judgement:
- For E. faecalis, the ELS did not provide any studies describing the impact of dry heat on its inactivation in feathers and down or other matrices. Due to this lack of evidence, it was not possible to conclude that a 5 log_10_ reduction is achieved by applying dry heat at a temperature of 100°C for 30 min (AQ1).
- For S. Senftenberg, the available datasets are highly inconsistent and conflicting, with no clear trend emerging. Due to the conflicting evidence, it was not possible to conclude that a 5 log_10_ reduction is achieved by applying dry heat at a temperature of 100°C for 30 min (AQ1).
- For Anelloviridae and Circoviridae, only one study on dry heat was retrieved, which involved different conditions and matrices compared to those specified in the mandate. In this study, a 3 log_10_ reduction was not achieved. Therefore, it was not possible to conclude that a 3 log_10_ reduction is achieved by applying dry heat at a temperature of 100°C for 30 min (AQ2).
Based on the data extracted out of the ELS, no conclusive evidence was found to demonstrate that the proposed method can: (i) achieve at least a 5 log_10_ reduction in E. faecalis or S. Senftenberg concentrations and (ii) reduce the infectivity titre of Anelloviridae and Circoviridae by at least 3 log_10._
Therefore, based on data available to date, applying dry heat to feathers and down at a temperature of 100°C for 30 min cannot be considered equivalent to the treatment specified in Regulation (EU) No 142/2011, in terms of inactivation of relevant pathogens (AQ3).
INTRODUCTION
1
Background and Terms of Reference as provided by the requestor
1.1
The Canadian Food Inspection Agency (CFIA) asked the Commission to initiate a risk analysis to verify whether or not a heat treatment provided by CFIA, consisting of dry heat to a temperature of 100°C or higher for at least 30 min and described in Annex 2, can be considered equivalent to the one set out in Chapter VII, point C of Annex XIII to Regulation (EU) No 142/20111 for the export of treated feathers to the EU.
Chapter VII, Point C of Annex XIII to Regulation (EU) No 142/2011 defines an end point for feathers and down which have been factory‐washed and treated with hot steam at 100°C for at least thirty (30) min.
The Commission would like to explore if the heat treatment described in Annex 2 provided by CFIA offers equivalent safety of the respective derived product.
In the light of the above, and in accordance with Article 29 of Regulation (EC) No 178/2002,2 the Commission requests EFSA to provide a scientific opinion concerning the equivalence of the heat treatment process of feathers and down consisting of dry heat to a temperature of 100°C or higher for at least 30 min, with the treatment set up in Chapter VII, Point C of Annex XIII to Regulation (EU) No 142/2011, in terms of inactivation of relevant pathogens.
Interpretation of the Terms of Reference
1.2
Definition of equivalence
A previous EFSA Scientific Opinion on the Inactivation of indicator microorganisms and biological hazards by standard and/or alternative processing methods in Category 2 and 3 animal by‐products and derived products to be used as organic fertilisers and/or soil improvers (EFSA BIOHAZ Panel , 2021) evaluated the effectiveness of the treatment required for the declaration of feathers and down as an end point in the manufacturing chain for inactivating indicator microorganisms and biological hazards. This treatment involves factory‐washing the feathers and down and then treating them with hot steam at 100°C for at least 30 min.
EFSA determined that the necessary level of inactivation should meet the requirements for alternative transformation parameters for biogas and composting plants. These requirements are outlined in point 1 of Section 2 of Chapter III of Annex V to Regulation (EU) No 142/2011. According to the Regulation, the validation of the intended process must demonstrate that the process achieves the following overall risk reduction:
-
for thermal and chemical processes by:
-
– a reduction of 5 log _ 10 _ of E. faecalis or S. Senftenberg (775W, H _ 2 _ S negative),
-
– a reduction of infectivity titre of thermoresistant viruses, such as parvovirus, by at least 3 log _ 10 _, whenever they are identified as a relevant hazard; and
-
as regards chemical processes also by:
-
– a reduction of resistant parasites, such as eggs of Ascaris sp., by at least 99.9% (3 log _ 10 _ ) of viable stages;
EFSA BIOHAZ Panel (2021) assessed whether a treatment of factory‐washed feathers and down with hot steam at 100°C for at least 30 mins would achieve: (i) a reduction of 5 log_10_ of E. faecalis or S. Senftenberg (775W, H_2_S negative) and (ii) a reduction of infectivity titre by at least 3 log_10_ of Anelloviridae (chicken anaemia virus) and Circoviridae (duck circovirus), the most thermoresistant viruses families that were identified as a relevant hazard in the material to be treated.
The approach followed in EFSA BIOHAZ Panel (2021) is in line with the one regularly followed in EFSA BIOHAZ assessments of applications for new alternative methods for animal by‐products. The degree of hazard reduction achieved by the processing methods approved for the different categories of ABPs described in Chapter III, Annex IV of Commission Regulation (EU) No 142/2011 is not specified, and no definitive standards have been set down either in relation to hazard reduction levels for alternative methods dealing with ABPs. However, over the years the approach followed by EFSA BIOHAZ for assessing the equivalence of alternative methods to the processing methods currently approved in the Commission Regulation (EU) No 142/2011 has been harmonised, and in relation to Category 3 ABPs is as follows (EFSA BIOHAZ Panel, 2022):
In order to be considered at least equivalent to the processing methods approved in the legislation, the alternative methods for Category 3 ABP should be capable of reducing the concentration of the relevant pathogenic bacteria by at least 5 log _ 10 _ and the infectious titre of the relevant viruses by at least 3 log _ 10 _ *(*EFSA BIOHAZ Panel, 2005 ). (…). The determination of the relevant pathogenic bacteria and viruses should be defined by the hazard identification, specific for the material to be treated. If the hazard identification considers spore‐forming pathogenic bacteria to be relevant, the required level of inactivation will also be a 5 log _ 10 _ reduction of spores from these bacteria, with the exception of spores of C. botulinum for which a 12 log _ 10 _ reduction would be required, as for processing canned petfood.
Based on the available background information, the BIOHAZ Panel agreed that the heat treatment process for feathers and down, involving dry heat at 100°C or higher for at least 30 mins, can be considered at least equivalent to the treatment outlined in Chapter VII, Point C of Annex XIII to Regulation (EU) No 142/2011 in terms of inactivation of relevant pathogens, provided that the process can (i) reduce E. faecalis or S. Senftenberg concentrations by 5 log_10_ and (ii) reduce the infectivity titre of Anelloviridae (chicken anaemia virus) and Circoviridae (duck circovirus), the most heat‐resistant virus families identified as relevant hazards in the material to be treated, by at least 3 log_10_.
The assessment of the equivalence between the two methods was performed by addressing the following assessment questions (AQs):
AQ1: Is a 5 log_10_ reduction of E. faecalis and S. Senftenberg concentrations achieved by the proposed process (i.e. dry heat to a temperature of 100°C or higher for at least 30 min)?
AQ2: Is a 3 log_10_ reduction of Anelloviridae family (chicken anaemia virus (Gyrovirus chickenanemia)) and Circoviridae family (duck circovirus (Circovirus duck)), achieved by the proposed process (i.e. dry heat to a temperature of 100°C or higher for at least 30 min)?
AQ3: Is the heat treatment process of feathers and down consisting of dry heat to a temperature of 100°C or higher for at least 30 min, equivalent to the treatment set up in Regulation (EU) No 142/2011, in terms of inactivation of relevant pathogens?
DATA AND METHODOLOGIES
2
Thermal inactivation of selected bacteria and virus indicators (AQ1, AQ2)
2.1
The BIOHAZ panel concluded that the bacterial and viral hazards identified for feathers and down in the 2021 EFSA Scientific Opinion (EFSA BIOHAZ Panel, 2021) (Section 1.2) were also relevant to the current assessment. Additionally, the Panel found no evidence of new relevant hazards in feathers and down since the 2021 publication. The same hazards were used in this assessment.
Extensive literature searches (ELS) were conducted to identify relevant peer‐reviewed literature on thermal inactivation of E. faecalis, S. Senftenberg (AQ1), Anelloviridae (chicken anaemia virus) and Circoviridae (duck circovirus) (AQ2), for different time/temperature combinations, by dry heat and/or in matrices with low moisture (even if different from those in the mandate).
The following electronic databases were searched: Biosis (Web of Science, Clarivate); CAB Abstracts (Web of Science, Clarivate); Embase (Embase.com); Scopus (Scopus.com); Web of Science Core Collection including Science Citation Index, Conference Proceeding Citation Index Science, Book Citation Index Science, Emerging Sources Citation Index (Web of Science, Clarivate).
A search strategy was designed for each assessment question. The search terms included controlled vocabulary (i.e. CAB Thesaurus, Emtree) and free‐text terms to represent the bacteria and virus of interest and elements related to thermal inactivation and thermo‐resistance. They were selected based on previous EFSA opinions, the WG experts and information specialist knowledge and enriched with the support of tools such as PubReMiner3 and ChatGPT.4
The only limit that was applied in the search was English as a language of publication. The reproducible searches for all databases are available in Appendix A.
The searches were conducted on 17 July 2024. The resulting citations of each database were imported into EndNote software (Clarivate). In total, the search yielded 3189 references for AQ1, and 348 references for AQ2. De‐duplication was performed using the de‐duplication functions included in EndNote, supplemented by the Deduplicator tool,5 a component of the Systematic Review Accelerator6 (Bond University). This process reduced the dataset to 1307 unique records for AQ1 and 152 for AQ2 with 3 articles in common for both questions. These datasets were then imported into the DistillerSR7 software (DistillerSR Inc.) for further review and analysis.
The criterion for selection of references was experimental studies in which resistance parameters (D‐ and/or z‐values) or the levels of reduction or inactivation of the selected bacteria and viruses (preferably measured in log_10_) had been measured after thermal treatment in matrices preferably like the ones included in the mandate.
The Title and abstract screening step was performed in parallel by two reviewers. Each reference was evaluated to determine if it was related to:
-
thermal inactivation at different time/temperature combinations in defined matrices (even if different from the one in the mandate) focusing on dry heat and/or low moisture matrices (representing dry heat); AND
-
any of the following microbial indicators:
-
○ Enterococcus faecalis OR Salmonella Senftenberg (bacterial indicators)
-
○ Selected thermoresistant viruses identified in the 2021 EFSA Scientific Opinion (EFSA BIOHAZ Panel, 2021) (i.e. family Anelloviridae (chicken anaemia virus (CAV)) and family Circoviridae (duck circovirus))
The possible reply options were ‘Yes’, ‘Doubtful’ if it was unclear and ‘No’. In case of disagreement between reviewers the conflicts were solved after discussion of both views and reach of an agreement, before proceeding to Full‐text screening.
The Full‐text screening step was performed in parallel by two reviewers. The same question asked at Title and abstract screening had to be answered. The possible reply options were ‘Yes’ and ‘No’. Articles that received a ‘Yes’ reply were then categorised as dealing with the following hazards: ‘Bacteria’, ‘Viruses’, ‘Both’, ‘Not applicable’. In case of disagreement the conflicts had to be solved.
Exclusion criteria: papers on liquid culture media, heat treatments < 60°C, aerobic and anaerobic digestions were excluded. Papers before 1984 were also excluded.
At the Data extraction step, the following information was extracted:
- product group in which the indicator was treated (e.g. liquid products, solid products, solid food products, etc.);
- product type/ matrix;
- type of treatment (i.e. dry heat, wet heat, not specified);
- temperature applied for the treatment in°C;
- time of exposure in minutes;
- values reported (e.g. mean D value in min, mean D, SD, D, range of D, 95% CI D or not reported).
For the articles dealing with viruses, the name of the virus was recorded while for articles dealing with bacteria, both the name of the bacterium and the strain were recorded.
Results from ELS:
- 1456 titles and abstracts were screened in parallel by two experts;
- 176 full papers were classified/ screened in parallel by two experts;
- 46 papers were used for data extraction: 35 for bacteria and 11 for the two selected viral families.
The data sets extracted from these 46 papers are available in Appendix C (S. Senftenberg and E. faecalis) and Appendix D, Table D.1 (selected thermoresistant viruses from Anelloviridae and Circoviridae families).
Uncertainty analysis and expert judgement procedure
2.2
Based on the EFSA guidance on Uncertainty Analysis in Scientific Assessments (EFSA Scientific Committee, 2018a) and the scientific opinion on the principles and methods behind EFSA's Guidance on Uncertainty Analysis in Scientific Assessment (EFSA Scientific Committee, 2018b), the sources of uncertainty associated with the available data were summarised in tabular format (Table 4 in Section 3.4), describing also the cause of the uncertainty and the expected impact on the outcome of the assessment.
To address AQ1 and AQ2, the following questions were answered by each individual expert within the working group, based on the collected evidence and indicated uncertainties:
- What is the probability that a 5 log_10_ reduction of E. faecalis is achieved by the application of dry heat to a temperature of 100°C for 30 min to feathers and down, assuming that the process is performed as prescribed and that the indicated process conditions are achieved?
- What is the probability that a 5 log_10_ reduction of S. Senftenberg is achieved by the application of dry heat to a temperature of 100°C for 30 min to feathers and down, assuming that the process is performed as prescribed and that the indicated process conditions are achieved?
- What is the probability that a 3 log_10_ reduction of Anelloviridae (chicken anaemia virus) and Circoviridae (duck circovirus) is achieved by the application of dry heat to a temperature of 100°C for 30 min to feathers and down, assuming that the process is performed as prescribed and that the indicated process conditions are achieved?
Due to the immediate and clear agreement among experts concerning the analysis of the data obtained, it was decided that a written procedure and a facilitator were not needed to reach a consensus on the probability range. The experts in the WG expressed the probability values to answer the questions above by selecting one of the probability ranges provided in the approximate subjective probability scale of the EFSA guidance on communication of uncertainty (EFSA, 2019). The individual judgements were discussed, and a probability range was agreed upon by consensus judgement through open discussion.
ASSESSMENT
3
ABP materials considered in the mandate
3.1
Regulation (EC) No 1069/20098 defines ‘animal by‐products’ as ‘entire bodies or parts of animals, products of animal origin or other products obtained from animals, which are not intended for human consumption, including oocytes, embryos and semen’; and ‘derived products’ as ‘products obtained from one or more treatments, transformations or steps of processing of animal by‐products’.
The use or disposal of ABP depends on their risk classification in three different categories: (i) Category 1 material consists mainly of material that is considered at transmissible spongiform encephalopathy (TSE) risk, and as such represents the highest risk material; (ii) Category 2 material includes fallen stock, manure and gastrointestinal contents; (iii) Category 3 materials are considered of a lower risk level and include parts of animals that have been considered fit for human consumption at the slaughterhouse, but that are not intended for human consumption for production or commercial reasons (e.g. trimmings of carcasses, consumer rejection to certain organs, etc.).
Within the framework of this assessment, Article 10 of Regulation (EC) No 1069/2009 lists feathers among the Category 3 ABP materials originating from: ‘(b) (…) animals that have been slaughtered in a slaughterhouse and considered fit for human consumption following the ante‐mortem inspection or game killed for human consumption in accordance with Community legislation; (h) (…) live animals that did not show any signs of diseases communicable through that product to humans or animals; (n) (…) dead animals that did not show any sign of disease communicable to humans or animals’.
Under Regulation (EC) No 1069/2009, an end point is determined in the manufacturing chain for animal by‐products which are considered to no longer present a risk to public or animal health. Products which have reached this end point are exempt from controls under the Regulation and are allowed to be placed on the market without restriction under the Regulation and to be handled and transported by operators which have not been approved or registered in accordance with the Regulation.
Article 3 of Regulation (EU) No 142/2011 sets out the end point for feathers and down by stating that these materials must fulfil the specific requirements set out in point C of Chapter VII of Annex XIII. Point C states that ‘Feathers, parts of feathers and down which have been factory‐washed and treated with hot steam at 100°C for at least 30 minutes may be placed on the market without restrictions in accordance with this Regulation’.
Due to the diverse range of chemical hazards and microbiota, including pathogenic organisms, that may be present on the feathers, they must undergo prompt treatment.
Feathers and down are used for a variety of purposes, including clothing, bedding, fashion, textiles, crafts and decor. More industrial applications include sound‐proofing materials and insulating components. Poultry feathers are rich in keratin protein, which makes them a good source of nitrogen fertiliser (EFSA BIOHAZ Panel, 2021). In the EU, feather waste can be incinerated, composted with manure, hydrolysed or treated in dimethyl sulfoxide or other solvents (EFSA BIOHAZ Panel, 2021).
Indicator microorganisms considered in the mandate
3.2
Data on inactivation of indicator microorganisms on industrial‐scale systems are generally recommended to determine the inactivation efficacy of a process for biological hazards. Ideally, the performance and validity of an indicator should be established for each selected inactivation process and matrix of concern. However, some indicator organisms are widely recognised as valuable for process validation tests. In the case of the indicator organisms mentioned in the requirements for alternative composting and biogas treatments, according to point 1 of Section 2 of Chapter III of Annex V Commission Regulation (EU) No 142/2011, these indicator organisms are E. faecalis, S. Senftenberg and the most thermal resistant viruses relevant in the matrix under assessment (EFSA BIOHAZ Panel, 2021). In the EFSA 2021 opinion, the most thermal resistant viruses identified in feather and down were: chicken anaemia virus (G. chickenanemia), family Anelloviridae and duck circovirus (Circovirus duck), family Circoviridae.
Thermal inactivation data for the indicator microorganisms
3.3
Inactivation of biological hazards by dry heat versus wet heat
3.3.1
The ELS performed for this mandate, including specifically the term ‘dry heat’, provided evidence that the target microorganisms were more resistant when exposed to dry heat than to wet heating conditions.
Dry heating inactivates bacteria by causing oxidative damage to vital cellular components and denaturing proteins, leading to loss of function and structural integrity, ultimately resulting in cell death. The process is generally slower and requires higher temperatures compared to wet heating due to the absence of moisture, which enhances heat transfer and denaturation efficiency (Russell, 2003).
Moisture content has a crucial role in the effectiveness of heat destruction (Song and Kang, 2021). Finn et al. (2013) revised the responses and survival cell mechanisms in low‐moisture conditions. These authors explain that in low‐moisture conditions, bacterial cells attempt to quickly balance osmotic pressure, such as an influx of K^+^, followed by a longer‐term adaptation, through the uptake of compatible solutes (Csonka, 1989).
According to the Annex 2 submitted by the CFIA, before the heat treatment at 100°C for at least 30 min, the process of feathers and down includes ‘drying (with) natural heat on a stretched mesh frame in the sun outside or inside on a stretched mesh frame with a warm air flow al low temperature to drop the moisture content to around 20%’ followed by ‘drying in an oven with temperature control to reduce the moisture content to around 10%’.
In the ELS, few papers were retrieved on the impact of dry heating on the selected indicators in matrices for which the moisture content was indicated while most of the papers reported the water activity (a_w_).
Water activity and moisture content are related, but they are not the same. Water activity measures the availability of water for microbial growth and chemical reactions, while moisture content refers to the total amount of water in a substance as a percentage of its total weight (Garces‐Vega et al., 2019).
The relationship between moisture content and a_w_ is specific to each material and often needs empirical data or a moisture sorption isotherm (MSI) curve for accurate calculations. The a_w_ is temperature dependent and determines the resistance of the microorganisms to the heat inactivation. Differences in microbial inactivation in different foods at the same a_w_ have been attributed to specific MSI. However, studies investigating whether differences in MSI of different foods impact the heat resistance of bacteria, as Salmonella, were confounded by experimental protocols that dried and equilibrated the bacterial inoculum in different food matrices prior to thermal treatment (Dhaliwal et al., 2021).
It is generally acknowledged that a_w_ influences thermal inactivation of microorganisms, and that in matrices with high‐water content, the resistance of microorganisms to thermal inactivation is lower (Syamaladevi et al., 2016). Low‐a_w_ matrices lead to an increased thermal resistance in several microorganisms (e.g. Laroche & Gervais, 2003; Lang et al., 2017). Microbial viability at low‐a_w_ conditions is dependent on many factors, including heat or acid stress during drying, temperature and time of exposure, type of strains used, medium composition and the nature of the solutes present in the food matrix (Doyle & Mazzotta, 2000).
Enterococcus faecalis
3.3.2
The ELS did not provide papers describing the impact of dry heating on inactivation of E. faecalis in feather and down. Only five papers were retrieved, describing the impact of wet heating on solid food matrixes. Details on a_w_ or moisture content were not provided but they were always high, except for whole raw almond kernels (Table 1). See Appendix C, Table C.1, for details.
In Harris et al. (2012), the D value for E. faecalis was calculated from tryptic soy agar enumeration data obtained onto inoculated almonds, before and after a treatment in hot water at 88°C. The log_10_ reduction during the first 30 sec of treatment corresponded to 3.5 log_10_ and the D value to 0.36 min. Riva et al. (1991) experimentally spiked E. faecalis onto homogenised defrosted mussels then cooked at 70°C. The D value for the indicator microorganism was 30–60 s. Similar D values were obtained by Saucier and Plamondon (2011) in ground beef. Mroz et al. (1985) experimentally spiked E. faecalis in ground pork meat and then wet heated it at both 90°C and 95°C. The D values calculated for E. faecalis at the two temperatures were 13.9 and 9.46 min, respectively. Other studies were performed at lower temperatures, which were not considered relevant for this assessment.
Salmonella Senftenberg
3.3.3
None of the 31 articles retrieved in the ELS, listed in Appendix B, evaluated the heat resistance of S. Senftenberg in feathers or down. However, there were several articles that examined the impact of dry heat treatments, or treatments of matrices with low water activity or moisture at time/temperature combinations close to the ones considered in the mandate (100°C, 30 min). From all datasets extracted through the ELS, only the papers applying dry heat at a temperature higher than 60°C were further analysed. The relevant datasets gathered from these articles are summarised in Table 2.
In some cases, log_10_ reductions were reported, in others, the corresponding D values. When only one log_10_ reduction for a single treatment time was reported, a proxy D value was calculated by dividing the treatment time by the log_10_ reduction value, assuming isothermal treatment and linearity between log_10_ reduction and time increments. 5D values were also calculated assuming linearity (five times the duration needed to achieve 1 log_10_ reduction).
Chen and Jiang (2017) investigated the resistance to heat of desiccation‐adapted S. Senftenberg 775W in aged broiler litter and composted turkey litter heat‐treated at 75°C and 85°C for approximately 50 min. The mean log_10_ reduction achieved for S. Senftenberg was always < 5 log_10_ but such inactivation occurred during the temperature come‐up time making it difficult to estimate the impact of the temperature treatment only. Dhaliwal et al. (2021) examined the impact of drying conditions, food composition and water activity (a_w_) on the desiccation tolerance and thermal resistance of S. Senftenberg ATCC 43845 in different food matrices, including pet food, binder formulations and skim milk powder. Following experimental spiking with Salmonella and equilibration to an a_w_ of 0.33, the samples underwent isothermal treatment at 70°C. Thermal inactivation data were fitted to the Weibull model, revealing that Salmonella inactivation was matrix‐dependent, with pet food exhibiting the lowest reduction. The authors attributed this difference to the potential non‐uniform distribution of the aqueous culture within multi‐component food systems, leading to localised microenvironments that influence Salmonella survival (Li et al., 2014).
The impact of both matrix and a_w_ is also clear in the thermal inactivation dataset from Doyle and Mazzotta (2000), with S. Senftenberg showing a D value at 64°C of 2.8 min in liquid whole eggs while in chocolate it showed a D value at 90°C of 36 min (Doyle and Mazzotta, 2000).
Furtado et al. (2023) observed a reduction of 4.3 log_10_ of S. Senftenberg at 100°C for 60 min in dried growth medium. Izurieta and Komitopoulou (2012) exposed shells of hazelnuts and cocoa to dry heat at 100°C for 30 min and reached 3.03 and ≥ 4.47 log_10_ reductions of S. Senftenberg, respectively. Lang et al. (2017) investigated the impact of thermal treatment cycles on the inactivation of dried S. Senftenberg incorporated into whole milk powder and equilibrated to various a_w_ levels (0.11, 0.25, 0.44 and 0.58). Thermal treatments were conducted at 90°C, 95°C and 100°C for 3 min in sealed vessels, to ensure a_w_ remained constant throughout the treatment process. The extent of microbial inactivation was then quantified following exposure to these conditions. The D values calculated at the different temperature and a_w_ conditions showed that a lower a_w_ corresponded to a greater thermal resistance. Sekhon et al. (2020) dry heated inoculated non‐fat milk powder at 80°C for 60 min and at 90°C for 14 min obtaining reductions of S. Senftenberg ranging between 4.4 and 4.8 og_10_, respectively. Song and Kang (2021) evaluated the efficacy of dry heat treatment at 95°C for 60 min to inactivate S. Typhimurium, S. Senftenberg and S. Enteritidis PT on raw almonds. Thermal treatments at 95°C reduced S. Senftenberg by 1.24 log_10_ in 15 min and by 2.20 log_10_ in 60 min. Wang et al. (2018) performed laboratory studies on thermal resistance of desiccation‐adapted S. Senftenberg 775W in aged broiler litter and predicted at least 5‐log_10_ reduction of S. Senftenberg in poultry litter compost, depending on the heating temperature and poultry litter moisture combination. The validation of these results was obtained in one turkey litter compost processor and one laying hen litter compost processor (Wang et al., 2021). In both plants, the conditions applied were adequate to reduce any potential Salmonella contamination by at least 5 log_10_ depending on heating temperature, moisture content and types of poultry litter compost. In both articles (Wang et al., 2018, 2021) the pathogen inactivation occurred during the come‐up time, making it difficult to estimate the impact of the treatment only. Zhou et al. (2020) assessed the thermal resistance of S. Senftenberg 775W on basil leaves during hot air drying at 80° for 60 min and 100°C for 40 min. By the end of the treatments S. Senftenberg was inactivated by more than 5 log_10_. Also in this paper, the drying time is included in the calculation, making difficult to estimate the impact of the treatment temperature for a given a_w_/moisture value.
To better visualise the most relevant data found in the literature and the effect of the time–temperature combinations to achieve a 5 log_10_ reduction of S. Senftenberg in the different matrices, a scatter chart was prepared for the thermal inactivation data (including only data related to temperatures ≥ 80°C). Results were plotted considering the time in minutes to reach 5 log_10_ reductions at different temperatures. Datasets with Salmonella serovars other than Senftenberg or mixes of serovars were excluded. Reported a_w_ values were categorised in 4 intervals ranging between 0.10 and 0.60. A total of 21 datasets (Table C.2 in Appendix C) were used to prepare the scatter chart (Figure 1). Studies in which the log_10_ reduction was calculated considering the initial come‐up time were not included.
Time–temperature combinations to achieve a 5 log10 reduction of S. Senftenberg in different matrices obtained from the literature search in relation to aw and for T ≥ 70°C. The red line corresponds to the mandate processing time (30 mins). The points correspond to the time for the 5 log10 reduction as extracted from the literature datasets. Points of datasets inside the three blue rectangles are from the same article (Lang et al. 2017). They correspond to milk powder matrices with different aw and subject to different treatment temperatures. These are the only data coming from true kinetic experiments (not proxy D values).
Most of the values were proxy D values derived from total log_10_ reduction for a specific treatment. The D values were explicitly reported only in one paper (Lang et al., 2017).
Clear trends are observable within datasets from the same study (Lang et al., 2017), where decreasing a_w_ correlates with an increase in resistance (higher D value), whereas increasing temperature (T) results in a decrease in resistance. These trends are visible as the three to four data points in the lower‐right quadrant of the plots (indicated inside the blue rectangles).
However, one study (Song and Kang, 2021) presents an opposite trend, where higher temperatures correlate with higher thermal resistance. This may be due to the fact that the heat treatment in this study was conducted under ‘open’ conditions (not sealed) and over a longer duration, leading to greater reductions in a_w_ and/or moisture content, which in turn increased resistance to temperature. Dry heat treatment of raw almonds at 95°C for 15 and 60 min reduced S. Senftenberg by 1.24 and 2.20 log_10_, respectively. At 90°C for 15 and 60 min, S. Senftenberg was reduced by 1.33 and 2.49 log_10_, respectively.
While it is possible to identify trends in datasets from single studies, it is challenging to draw overarching conclusions about the influence of temperature and a_w_ on D values across different studies. The impact of dry heating on Salmonella inactivation does not depend only on the temperature of the treatment and the a_w_ of the matrix but is the result of a complex interaction between different factors. Factors of relevance highlighted in the retrieved studies include strain tested, type of treatment applied (e.g. time/temperature profile, environmental conditions, etc.), intrinsic parameters and composition of the treated matrix, extrinsic factors, but also the study design/sample preparation.
Therefore, based on the data found in the literature, it is not possible to conclude if a 5 log_10_ reduction of S. Senftenberg can be achieved in feathers and down by dry heat to a temperature of 100°C for at least 30 min. The time needed for 5D values calculated from the available literature for similar temperatures, matrices, and low a_w_ conditions fluctuates above and below the 30‐min threshold (Figure 2) as the result of the effect of the additional factors described above.
In some of the papers retrieved in the ELS moisture values are reported instead of a_w._ Izurieta and Komitopoulou (2012) reported log_10_ reductions of 3.03 and ≥ 4.47 respectively in hazelnut shells and cocoa shells with 4% w/w moisture after 30 min at 100°C, Zhou et al. (2020) found log_10_ reductions > 6.2 after treatment of basil leaves with an 8.6% moisture content at 100°C for 40 min (but in this case the come‐up time is included).
Thermal resistance of other Salmonella serotypes
3.3.4
The scientific literature suggests that S. Senftenberg is the most heat‐resistant serovar in high‐water activity foods, with D values in different model systems around 10‐fold to 20‐fold higher than those of other serovars, such as S. Typhimurium or S. Enteritidis (Doyle and Mazzotta, 2000).
However, interestingly, in some of the articles retrieved in the ELS, where the dry heat resistance, or the heat resistance in low moisture/a_w_ matrices, of S. Senftenberg was compared with that of other serovars of S. enterica, this behaviour was not observed.
Furtado et al. (2023) investigated the heat tolerance of different Salmonella serovars previously adapted to desiccation showing differences in the heat tolerance in all temperature–time combinations tested. The reductions in the counts of S. Abony, S. Bredeney, S. Brooklyn, S. Heidelberg, S. Ohio and S. Typhimurium were approximately 1 log_10_ after 90 min exposure at 90°C, while that of S. Senftenberg was > 1.50 log_10_. After a thermal treatment at 100°C for 60 min S. Typhimurium showed a decrease of 3.37 log_10_ while S. Senftenberg of 4.30 log_10_. The results of this study highlight that there are two cellular mechanisms linked to dry heating inactivation: a_w_ adaptation and desiccation. Moreover, they suggest that the strategies to mitigate Salmonella in low‐a_w_ foods must consider the existence of high‐stress resistant serotypes and their multiple‐stress adaptability profiles, including effects of processing and food composition.
Sekhon et al. (2020) compared the thermal resistance of S. Senftenberg 775W with that of S. Enteritidis, S. Montevideo, S. Newport and S. Typhimurium in non‐fat dry milk. The heat resistance of S. Senftenberg 775W was found to be lower than or comparable to that of other serovars in non‐fat dry milk at 80°C and 90°C due to its weaker adaptability in low a_w_ environment. According to the results of this study the authors suggested to perform Salmonella thermal inactivation studies of low a_w_ matrices including a cocktail of serotypes and not only S. Senftenberg.
Dhaliwal et al. (2021) evaluated the effects of drying conditions, food composition and water activity on the desiccation tolerance and thermal resistance of S. Enteritidis FUA 1946, S. Senftenberg ATCC 43845 and S. Typhimurium ATCC 13311 in pet food, binder formulation and skim milk powder. The results showed that irrespective of the a_w_, food composition and physical structure of the selected foods, S. Enteritidis FUA 1946 displayed the highest desiccation and thermal resistance, followed by S. Senftenberg ATCC 43845 and S. Typhimurium ATCC 13311.
ELS was conducted only for thermal resistance data of S. Senftenberg because it is the only serovar listed in point 1 of Section 2 of Chapter III of Annex V Commission Regulation (EU) No 142/2011.
Based on evidence that certain strains of other Salmonella serovars may exhibit greater resistance to dry heat than S. Senftenberg, future evaluations of dry heat processes should consider other serovars as potential indicator organisms to assess inactivation efficacy.
Thermal resistant viruses
3.3.5
There is very limited evidence in the literature on the thermal inactivation of Anelloviridae and Circoviridae by dry heat or on low moisture/a_w_ matrices (Table 3) or in feather and down. The literature search conducted on viruses from these families resulted in the full‐text analysis of only nine articles assessing resistance to heat of chicken anaemia virus (CAV) and/or porcine circovirus (PCV). In the search for Circoviridae data on porcine circovirus were retrieved while no study addressed duck circovirus.
In most cases, the retrieved studies subjected the viruses to a wet heat treatment performed with a viral suspension in liquid medium (e.g. porcine plasma or a lab made liquid medium) or in a high moisture solid matrix (e.g. minced meat) at temperatures ranging from 60 to 100°C, with variable reductions in infectivity being reported. In those cases where resistance at 100°C was studied, the following was observed:
- –Complete inactivation of the inoculum concentration of chicken anaemia virus added in an aqueous fraction (initial load unknown) after a 15 min wet heat treatment (Goryo et al., 1985).
- –Complete inactivation (at least 5 log_10_ reduction) of chicken anaemia virus in minced meat from chicken carcasses after discarding feathers, skin and feet after a 30 min wet heat treatment (Urlings et al., 1993).
A single research study detailing thermal inactivation of Anelloviridae and Circoviridae by dry heat was found (Welch et al., 2006). It assessed the inactivation of chicken anaemia virus and porcine circovirus resuspended in human Factor VIII concentrate, freeze‐dried and heated at 80°C for up to 72 h and 120°C for 30 min. The final reductions observed were: 0.75 log_10_ for porcine circovirus after 72 h at 80°C; 1.25 log_10_ for chicken anaemia virus after 72 h at 80°C; approximately 1 log_10_ reduction in infectivity for both chicken anaemia virus and porcine circovirus after 30 min at 120°C. The evidence in Welch et al. (2006) thus highlights that chicken anaemia virus and porcine circovirus display significant thermal resistance to dry heat treatments.
Uncertainty analysis
3.4
Experts identified uncertainties which can affect the outcome of the ToRs. Table 4 includes a list of uncertainty sources affecting the different AQs and their impact on the conclusions.
Answers to assessment questions on the reduction of the selected indicators by dry heating at 100°C for 30 min (AQ1 and AQ2)
3.5
The ELS did not provide any papers describing the impact of dry heat on the inactivation of the selected indicators in feathers and down.
For E. faecalis, no study showed if dry heat at 100°C for 30 min achieves a 5 log_10_ reduction. Based on the literature review, the 5 log_10_ reduction was obtained in wet heating conditions, where E. faecalis was rapidly inactivated even at temperatures < 100°C. Since there is evidence that low a_w_ /dry heating conditions induce higher heat resistance of a wide range of microorganisms in comparison to wet heating, it cannot be assured that for E. faecalis a 5 log_10_ reduction is achieved in feathers and down with the treatment under assessment. According to the EFSA guidance on communication of uncertainty in scientific assessments (EFSA, 2019), considering all the evidence and sources of uncertainty, the experts judged that it is 0%–100% certain (inconclusive) that a 5 log_10_ reduction of E. faecalis is achieved by applying dry heat at a temperature of 100°C for 30 min to feathers and down, assuming that the process is performed as prescribed and that the indicated process conditions are met.
For S. Senftenberg, several studies available in the scientific literature showed that the heat resistance of this serovar is influenced by water activity, which typically decreases during dry heating (Song and Kang, 2021). Zhou et al. (2020) showed that the thermal resistance of S. Senftenberg increases significantly as the a_w_ decreases. Several articles reporting data in matrices with low a_w_ at time/temperature combinations close to the one considered in the mandate provide reductions below 5 log_10_ for dry heat treatments at 100°C for 30 min and even up to 90 min (Furtado et al., 2023; Izurieta and Komitopoulou, 2012). As none of the studies assessed the dry heat resistance of S. Senftenberg in feathers and down, and the a_w_ of feathers and down is not known (and can be influenced by the pre‐processing steps), there is no clear evidence demonstrating that dry heat treatment at 100°C for 30 min can achieve a 5 log_10_ reduction of S. Senftenberg in the matrix under assessment. According to the to the above mentioned guidance (EFSA, 2019), considering all the evidence and sources of uncertainty, the experts judged that it is 33%–66% certain (as likely as not) that a 5 log_10_ reduction of S. Senftenberg is achieved by applying dry heat at a temperature of 100°C for 30 min to feathers and down, assuming that the process is performed as prescribed and that the indicated process conditions are met.
For Anelloviridae family (chicken anaemia virus (Gyrovirus chickenanemia)) and Circoviridae family (duck circovirus (Circovirus duck)), there is also limited evidence regarding their survival/inactivation under dry heat treatments. Just a few experimental articles provide data on their heat inactivation, and most of them were performed in liquid viral suspensions and at temperatures well below 100°C. The only experimental study providing data on the inactivation of Anelloviridae and Circoviridae under dry heat reported low reductions, of approximately 1 log_10_ after 30 min treatment at a temperature of 120°C. Alongside the scarcity of data, this finding casts doubts on the capacity of a dry heat treatment at 100°C for 30 min to achieve a 3 log_10_ reduction of Anelloviridae and Circoviridae in feathers and down. According to the to the above mentioned guidance (EFSA, 2019), considering all the evidence and sources of uncertainty, the experts judged that it is 50%–100% certain (more likely than not) that a 3 log_10_ reduction of Anelloviridae and Circoviridae is not achieved by applying of dry heat at a temperature of 100°C for 30 min to feathers and down, assuming that the process is performed as prescribed and that the indicated process conditions are met.
Based on the data extracted out of the ELS (Tables 1, 2, 3, Figure 1), the summarised sources of uncertainties (Table 4) and the expertise in the WG, the following conclusions were achieved:
- For E. faecalis, the ELS did not provide any studies describing the impact of dry heat on its inactivation in feathers and down or in other matrices. Due to this lack of evidence, it was not possible to conclude that a 5 log_10_ reduction is achieved by applying dry heat at a temperature of 100°C for at least 30 min (AQ1).
- For S. Senftenberg, the available datasets are highly discrepant and conflicting, with no clear trend emerging, as heat inactivation is the result of complex interactions between strain tested, type of treatment and intrinsic and extrinsic parameters. Due to the conflicting evidence, it was not possible to conclude that a 5 log_10_ reduction is achieved by applying dry heat at a temperature of 100°C for at least 30 min (AQ1).
- For Anelloviridae and Circoviridae families, only one study on dry heat was retrieved (with different conditions and matrices compared to those of the mandate), in which a 3 log_10_ reduction was not achieved (AQ2). Therefore, it was not possible to conclude that a 3 log_10_ reduction is achieved by applying dry heat at a temperature of 100°C for at least 30 min.
CONCLUSIONS
In reply to AQ3, based on the data available to date, no conclusive evidence was found to demonstrate that the proposed method can (i) reduce E. faecalis or S. Senftenberg concentrations by at least 5 log_10,_ and (ii) reduce the infectivity titre of Anelloviridae family and Circoviridae family, by at least 3 log_10_. Therefore, applying dry heat to feathers and down at a temperature of 100°C for 30 min cannot be considered equivalent to the treatment set up in Commission Regulation (EU) No 142/2011, in terms of inactivation of relevant pathogens.GLOSSARY5D valuesFive times the decimal reduction time (the time required to achieve a five‐log reduction in the microbial population)Animal by‐product (ABP)Entire bodies, parts of animals or products of animal origin that are not intended for human consumption, including oocytes, embryos and semen. D valuesTime required to reduce the microbial population at a given temperature by 1 log_10_ unitlog_10_ Logarithmic scale base 10 (used to express microbial reduction) ABBREVIATIONSABPAnimal by‐productATCCAmerican Type Culture CollectionAQAssessment Question a w Water activityBIOHAZ PanelEFSA Panel on Biological HazardsCAVChicken anaemia virusCFIACanadian Food Inspection AgencyELSExtensive literature searchMSIMoisture Sorption IsothermPCVPorcine circovirusToRTerms of ReferenceTSETransmissible Spongiform EncephalopathyWGWorking Group
REQUESTOR
European Commission
QUESTION NUMBER
EFSA‐Q‐2024‐00169
COPYRIGHT FOR NON‐EFSA CONTENT
EFSA may include images or other content for which it does not hold copyright. In such cases, EFSA indicates the copyright holder and users should seek permission to reproduce the content from the original source.
PANEL MEMBERS
Ana Allende, Avelino Alvarez‐Ordóñez, Valeria Bortolaia, Sara Bover‐Cid, Alessandra De Cesare, Wietske Dohmen, Laurent Guillier, Lieve Herman, Liesbeth Jacxsens, Maarten Nauta, Lapo Mughini‐Gras, Jakob Ottoson, Luisa Peixe, Fernando Perez‐Rodriguez, Panagiotis Skandamis, and Elisabetta Suffredini.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Csonka, L. N. (1989). Physiological and genetic responses of bacteria to osmotic stress. Microbiological Reviews, 53(1), 121–147. 10.1128/mr.53.1.121-147.1989 2651863 PMC 372720 · doi ↗ · pubmed ↗
- 2EFSA (European Food Safety Authority) , Hart, A. , Maxim, L. , Siegrist, M. , Von Goetz, N. , da Cruz, C. , Merten, C. , Mosbach‐Schulz, O. , Lahaniatis, M. , Smith, A. , & Hardy, A. (2019). Guidance on communication of uncertainty in scientific assessments. EFSA Journal, 17(1), 5520, 73 pp. 10.2903/j.efsa.2019.5520 PMC 729219132626067 · doi ↗ · pubmed ↗
- 3EFSA BIOHAZ Panel (EFSA Scientific Panel on Biological Hazards) . (2005). Scientific opinion on vis‐à‐vis biological risks of biogas and compost treatment standards of animal by‐products (ABP). EFSA Journal, 3(9), 264, 21 pp. 10.2903/j.efsa.2005.264 · doi ↗
- 4EFSA BIOHAZ Panel (EFSA Scientific Panel on Biological Hazards) . (2021). Scientific opinion on the inactivation of indicator microorganisms and biological hazards by standard and/or alternative processing methods in category 2 and 3 animal by‐products and derived products to be used as organic fertilisers and/or soil improvers. EFSA Journal, 19(12), 6932, 111 pp. 10.2903/j.efsa.2021.6932 PMC 863856134900004 · doi ↗ · pubmed ↗
- 5EFSA BIOHAZ Panel (EFSA Panel on Biological Hazards) , Koutsoumanis, K. , Allende, A. , Bolton, D. , Bover‐Cid, S. , Chemaly, M. , Davies, R. , De Cesare, A. , Herman, L. , Hilbert, F. , Lindqvist, R. , Nauta, M. , Peixe, L. , Ru, G. , Simmons, M. , Skandamis, P. , Suffredini, E. , Fernández Escámez, P. , Griffin, J. , … Alvarez‐Ordoñez, A. (2022). Scientific opinion on the evaluation of a multi‐step catalytic co‐processing hydrotreatment for the production of renewable fuels · doi ↗ · pubmed ↗
- 6EFSA Scientific Committee , Benford, D. , Halldorsson, T. , Jeger, M. J. , Knutsen, H. K. , More, S. , Naegeli, H. , Noteborn, H. , Ockleford, C. , Ricci, A. , Rychen, G. , Schlatter, J. R. , Silano, V. , Solecki, R. , Turck, D. , Younes, M. , Craig, P. , Hart, A. , Von Goetz, N. , … Hardy, A. (2018 a). Guidance on uncertainty analysis in scientific assessments. EFSA Journal, 16(1), 5123, 39 pp. 10.2903/j.efsa.2018.5123 PMC 700972732625671 · doi ↗ · pubmed ↗
- 7EFSA Scientific Committee , Benford, D. , Halldorsson, T. , Jeger, M. J. , Knutsen, H. K. , More, S. , Naegeli, H. , Noteborn, H. , Ockleford, C. , Ricci, A. , Rychen, G. , Schlatter, J. R. , Silano, V. , Solecki, R. , Turck, D. , Younes, M. , Craig, P. , Hart, A. , Von Goetz, N. , … Hardy, A. (2018 b). Scientific opinion on the principles and methods behind EFSA's guidance on uncertainty analysis in scientific assessment. EFSA Journal, 16(1), 5122, 235 pp. 10.2903/j.efsa.2018 · doi ↗ · pubmed ↗
- 8Finn, S. , Condell, O. , Mc Clure, P. , Amézquita, A. , & Fanning, S. (2013). Mechanisms of survival, responses and sources of Salmonella in low‐moisture environments. Frontiers in Microbiology, 4, 5. 10.3389/fmicb.2013.00331 24294212 PMC 3827549 · doi ↗ · pubmed ↗