The role of PilU in the surface behaviors of Pseudomonas aeruginosa
Jingchao Zhang, Yan Luo, Yiwu Zong, Shangping Lu, Yi Shi, Fan Jin, Kun Zhao

TL;DR
This study explores how PilU affects the surface behaviors of Pseudomonas aeruginosa, revealing its role in cell movement and colony formation.
Contribution
The study identifies new roles of PilU in both single-cell and multicellular surface behaviors of Pseudomonas aeruginosa.
Findings
ΔpilU cells show increased TFP numbers but reduced movement and delayed microcolony formation.
ΔpilU cells form thick multilayered colony edges, slowing colony expansion on semi-solid surfaces.
Cell–cell collision responses shift from touch-turn to touch-upright dominance in ΔpilU mutants.
Abstract
In Pseudomonas aeruginosa, the dynamic activity of type IV pilus (TFP) is essential for various bacterial behaviors. While PilU is considered a homolog of the TFP disassembling motor PilT, its specific roles remain unclear. Using pilus visualization and single‐cell tracking techniques, we characterized TFP dynamics and surface behaviors in wild‐type and ΔpilU mutants. We found that ΔpilU cells displayed increased TFP numbers but reduced cell movement and delayed microcolony formation. Interestingly, beyond affecting the twitching motility, ΔpilU cells formed a thick multilayered colony edge on semi‐solid surfaces, slowing colony expansion. Cell–cell collision responses changed from touch‐turn dominance in wild type to touch‐upright dominance in ΔpilU, affecting colony morphology and expansion. These findings expand our understanding of PilU's physiological roles and provide potential…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
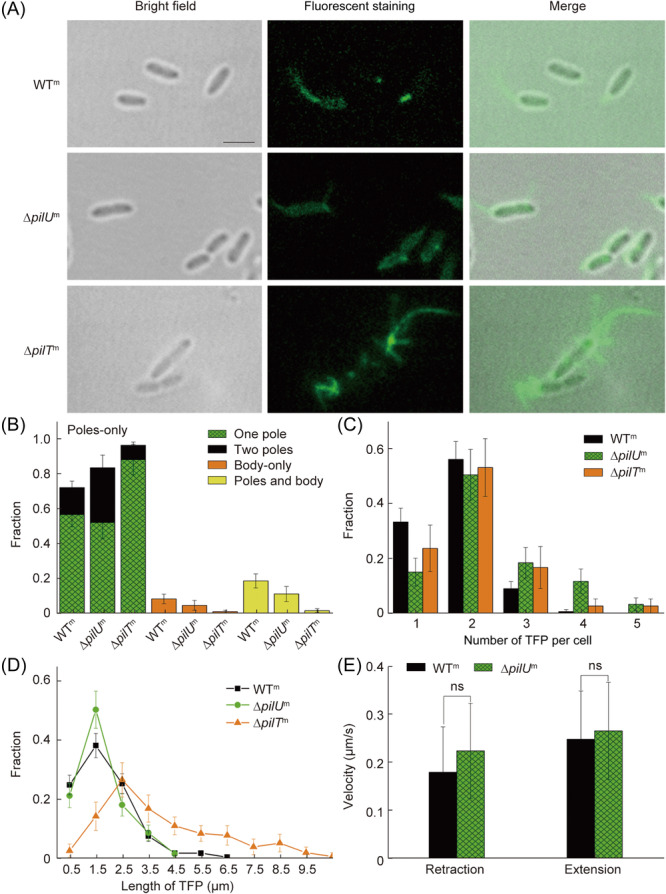 Figure 1
Figure 1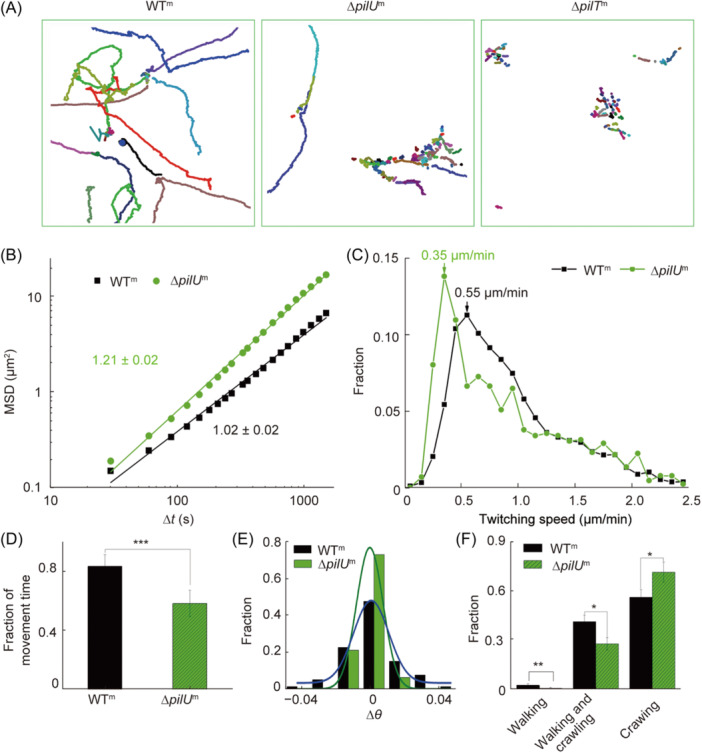 Figure 2
Figure 2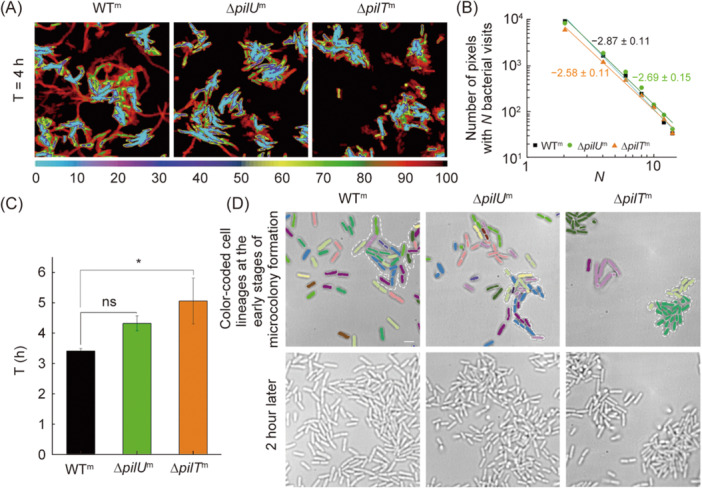 Figure 3
Figure 3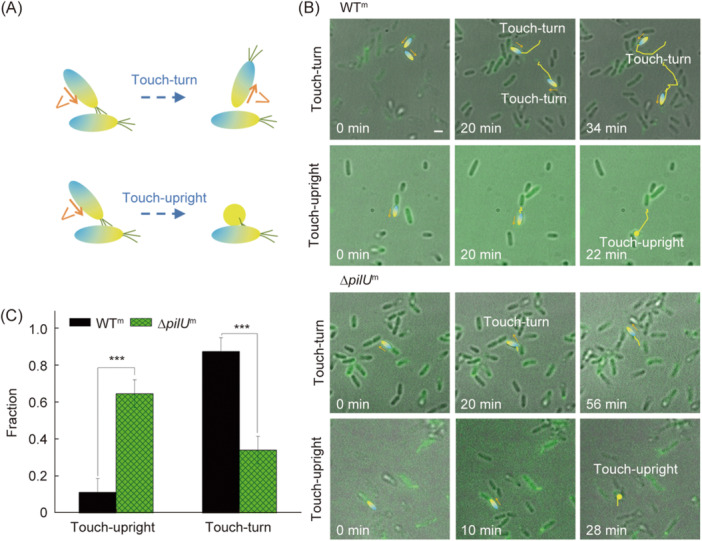 Figure 4
Figure 4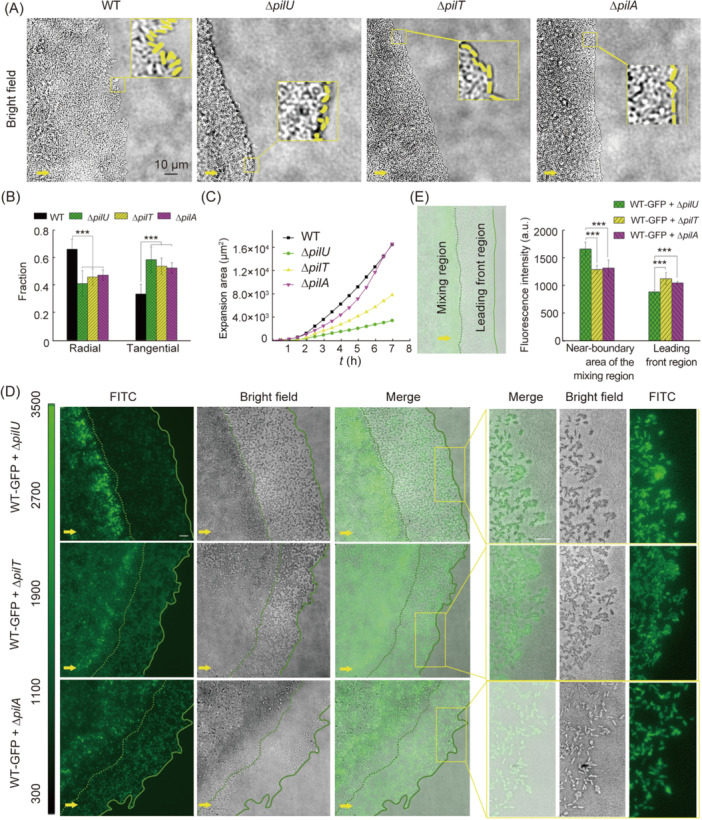 Figure 5
Figure 5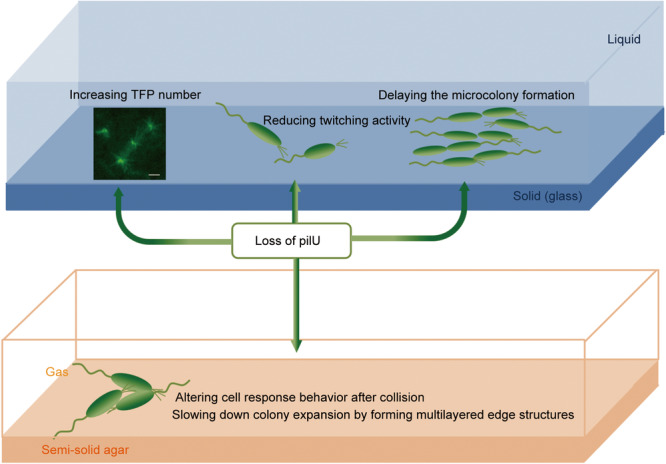 Figure 6
Figure 6- —National Key Research and Development Program of China
- —China Postdoctoral Science Foundation 10.13039/501100002858
- —Sichuan Provincial People's Hospital Research Fund of the Academy
- —National Natural Science Foundation of China 10.13039/501100001809
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsBacterial biofilms and quorum sensing · Vibrio bacteria research studies · Bacterial Genetics and Biotechnology
INTRODUCTION
Type IV pilus (TFP) is critical for a variety of bacterial behaviors, including cell movement, biofilm formation, adhesion, DNA uptake, surface sensing, and virulence1. One well‐recognized feature of TFP is that it can drive a special type of surface motion called twitching through the extension‐retraction cycles of TFP2, which has been observed in a broad range of bacteria species3, 4, 5, 6, 7. In addition to its role in the movement that leads to surface exploration, TFP also plays an important role in the transition from reversible to irreversible surface attachment8, 9. In fact, in Pseudomonas aeruginosa and Caulobacter crescentus, TFP is considered as one key part of surface sensing10, 11, 12, 13. For instance, Siryaporn et al. found that in P. aeruginosa, TFP contraction allowed cells to respond to surfaces of varying hardness and promoted the virulence of surface contact10. Similarly, in C. crescentus, Ellison et al. showed that upon surface contact, dynamic TFP activity stopped while fixation adhesins were produced, which then promoted irreversible adhesion12. Recent studies further revealed a role of TFP‐mediated surface sensing in triggering the initiation of the cell cycle of C. crescentus 14, 15.
TFP dynamic activity is critical for its function. In Gram‐negative bacteria, the TFP assembly system is composed of 10–18 different proteins, located in the inner membrane, periplasm, and outer membrane16. The main component of TFP is a protein subunit called major pilin, while minor pilins, although fewer in number, are also crucial for the assembly or specific functions of pilus17, 18. Biochemical studies have revealed detailed insights into the extension/retraction mechanisms of TFP, including the reverse rotational movement of PilC (pilus inner membrane core protein) driving the extension or retraction of the fiber, as well as the important roles of PilB (pilus assembling protein) and PilT (pilus disassembling protein) in pilus dynamics19, 20, 21, 22. Although PilU is a homology of PilT, its function is not clear yet. This is partially because PilU has been relatively poorly studied compared to PilT and the phenotypes of pilU mutants in different organisms are inconsistent22, 23, 24, 25, 26, 27, 28. For instance, in P. aeruginosa and Dichelobacter nodosus, loss of PilU has been shown to impair twitching motility23, 24, while in Neisseria gonorrhoeae, PilU mutant cells could still twitch and were capable of DNA transformation25. In Vibrio cholerae, ΔpilU mutants showed a slightly reduced retraction rate (~1.3‐fold) whereas in ΔpilT mutants the retraction rate was reduced to ~50‐fold, indicating that PilT is the true retraction motor of TFP22, 26. This is also true in other species including P. aeruginosa, Acinetobacter baylyi, and Acidithiobacillus ferrooxidans 22, 26, 27, 28. It has been suggested that PilU functions as a PilT‐dependent retraction ATPase and the coordination between PilT and PilU may be the mechanism for efficient pilin retraction22, 26, 27. However, the exact coordination of these two motors during this process is still not fully understood. Moreover, beyond its interaction with PilT, more broader impacts of PilU on bacterial surface behaviors still need to be explored.
The advances in pilin visualization techniques have greatly facilitated the research on TFP. Traditional fluorescence techniques such as immunofluorescence labeling and large‐volume fluorescent avidin compounds have been used to visualize TFP7, 8, 29, 30, 31, but these methods cannot get detailed dynamic information of native (intact) TFP‐associated activities and do not provide temporal resolution. Succinimide dyes combined with exposed primary amines have also been used to visualize the TFP of P. aeruginosa. Although this method enables the monitoring of pili extension and retraction, the affinity of fluorescent labels for pili is relatively non‐specific, resulting in the cell body also being stained with a more intense fluorescence due to its comparatively larger surface area, which increased the difficulties in analyzing TFP5. Recently, new techniques including a cysteine‐substitution‐based technique and a label‐free interference scattering (iSCAT) microscopy technique have been developed12, 27, which enables to directly observe TFP in a real‐time and in situ manner with both high temporal and high spatial resolutions.
In this study, we aimed to explore the roles of PilU in regulating P. aeruginosa surface behaviors through direct observation of TFP in situ and in real time. Toward this goal, we employed the cysteine‐substitution‐based technique together with bacterial tracking techniques32 to examine TFP dynamics, single‐cell motility, and collective behaviors in wild‐type and ΔpilU mutant strains. We first characterized the TFP morphology and TFP‐driven twitching motility in different mutant strains to show the PilU effect on single‐cell behaviors, then through monitoring the microcolony formation of different mutants on glass surfaces, as well as colony expansions on semi‐solid agar surfaces, the PilU effect on TFP‐based cell–cell interactions and the bacterial collective behavior was studied. Our findings suggest that PilU plays multiple roles in regulating P. aeruginosa surface behaviors beyond its known function in twitching motility.
RESULTS
pilU affects TFP number and locations on a cell surface but not TFP extension and retraction speed
To reveal the role of pilU in P. aeruginosa, we first characterized TFP morphology and activities in three strains, PAO1P_ BAD ‐pilAS99C (WT containing pili visualization plasmid), ∆pilUP BAD _‐pilAS99C, and ∆pilTP* BAD ‐pilAS99C (hereafter, they will be referred as WT^m^, ∆pilU ^ m ^, and ∆pilT ^ m ^, respectively). Figure 1A shows successful TFP staining examples of all three strains. With pili observable under fluorescence microscopy, the distribution, number, length, and retraction/extension speed of TFP were measured, and the results are shown in Figure 1B–E. TFP was observed to appear at cell poles and/or cell body parts. By classifying the distribution of TFP on cell surfaces into three groups, which are poles‐only (including one pole and two poles), body‐only, and poles and body, we found that in all three strains, pili mainly appeared at poles‐only, with a percentage of 72% ± 8% in WT^m^, 84% ± 12% in ∆pilU ^m^, and 97% ± 8% in ∆pilT ^m^ (Figure 1B). By further differentiating whether pili are located at one pole or two poles among these poles‐only events, we found that compared with WT^m^ that has a percentage of 16% ± 4% for two‐poles event, ∆pilU ^m^ displayed a percentage of 31% ± 7%, almost as two times as that of WT^m^, while ∆pilT ^m^ showed a reduced percentage to be 8% ± 2% (Figure 1B), suggesting different roles of PilU and PilT in the control of TFP distribution along cell bodies. Figure 1C shows the measured histograms of TFP numbers per cell. Compared with WT^m^, both ∆pilU ^m^ and ∆pilT ^m^ showed an increased percentage of cells with more TFP appearing (Figure 1C), indicating that the absence of PilU/PilT can lead to hyper‐piliation of cells. But ∆pilT ^m^ cells displayed a larger range of TFP length in 0.5–10 μm (Figure 1D), compared with that of WT^m^ (0.5–6.5 μm) and ∆pilU ^m^ (0.5–4.5 μm), suggesting that PilT is more important than PilU for controlling TFP length. Besides morphology, TFP dynamic activity (i.e., extension and retraction of TFP) was also evaluated. In ∆pilT ^m^, we did not observe TFP retraction events (data not shown), consistent with literature results27. In contrast, ΔpilU* ^m^ cells still exhibited extension‐retraction cycles with an extension speed of 0.26 ± 0.10 μm/s and a retraction speed of 0.22 ± 0.099 μm/s. These speeds showed no statistically significant difference from the corresponding values observed in WT^m^ (0.25 ± 0.10 μm/s for extension and 0.18 ± 0.094 μm/s for retraction) (Figure 1E). This suggests that the absence of pilU does not significantly affect the speed of TFP extension and retraction.
Characterization of type IV pilus (TFP) morphology of WTm, ∆pilU m, ∆pilT m. (A) Representative images of TFP including bright filed, fluorescence staining, and merged. Pili stained with AF‐488‐C5 are shown in green. (B) The distribution of TFP locations on cell surfaces. N (WTm) = 116, N (∆pilU m) = 61, and N (∆pilT m) = 157. (C)The distribution of TFP number per cell. N (WTm) = 131, N (∆pilU m) = 59, and N (∆pilT m) = 71. (D) The distribution of TFP length. For TFP length measurements, pili that were retracting at the start of image recording were not included in the analysis. N (WTm) = 225, N (∆pilU m) = 127, and N (∆pilT m) = 131. (E) The average extension and retraction velocity. For the measurement of TFP extension (retraction) velocity, only cells that performed a single TFP extension (retraction) within a 1‐min window were analyzed. N (WTm) = 73, and N (∆pilU m) = 28. Error bars show standard deviations. Statistical significances were measured using a two‐sample Student's t‐test. ns, not significant. Scale bars, 2 μm.
Motile ∆pilU
m cells show less twitching but more directional surface motion than WTm
Since TFP in P. aeruginosa is critical for cell surface movement, we next studied the effects of PilU and PilT on the TFP‐driven twitching motility by analyzing a time series of snapshots that recorded the bacterial surface behavior over a certain time period (see examples in Movie S1–S3 in Supporting Information section). Figure 2A are examples showing the obtained bacterial trajectories for WT^m^, ∆pilU ^m^, and ∆pilT ^m^. WT^m^ cells displayed a variety of trajectory patterns as expected. By contrast, ∆pilT ^m^ cells showed only dot‐like short traces. Note that such short traces are artifacts due to the cell elongation through cell growth as their TFP did not retract and cells thus did not twitch. ∆pilU ^m^, on the other hand, showed both long straight trajectories, which were definitely generated by cell movement, and short traces. As ∆pilT ^m^ did not twitch, we focused on the comparison between WT^m^ and ∆pilU ^m^. Although both WT^m^ and ∆pilU ^m^ cells could twitch, the proportion of motile cells was higher in WT^m^ (79% ± 16%) than in ∆pilU ^m^ (46% ± 14%) (Figure S1). The trajectories of motile cells can be quantitatively characterized by mean square displacements (MSDs), which assesses the degree to which cellular movement differs from random diffusion. Figure 2B shows the measured MSDs of WT^m^ and ∆pilU ^m^. The MSD of WT^m^ exhibited a fitting slope of 1.02 ± 0.02, suggesting a random motion of WT^m^ cells. By contrast, the MSD of ∆pilU ^m^ has a fitting slope of 1.21 ± 0.02, indicating a super‐diffusive‐like motion. To further understand the different surface motion shown by MSDs between WT^m^ and ∆pilU ^m^, we calculated twitching speed and the fraction of movement time during the total recording time for each motile cell (Figure 2C,D). Overall, the speed distribution of ∆pilU ^m^ was left‐shifted compared to WT^m^, indicating that the twitching activity of ∆pilU ^m^ is reduced compared to WT^m^ (Figure 2C). This is also consistent with the measured result of the fraction of movement time (Figure 2D), which is 0.59 ± 0.09 for ∆pilU ^m^ (i.e., for a tracked on‐average motile ∆pilU ^m^ cell, there is about 59% of the total tracked time during which the cell moved), but is 0.84 ± 0.08 for WT^m^. As the trajectory pattern is not only cell‐speed‐dependent but also affected by the directional persistence of cell movement, we examined the orientational angle difference of a cell between two consecutive frames, Δθ (Figure 2E). The results showed that the Δθ distribution of ∆pilU ^m^ was narrower than that of WT^m^. The Δθ of WT^m^ ranged from −0.04 to 0.04 rad while that of ∆pilU ^m^ ranged from −0.02 to 0.02 rad. We also counted the distribution of the angle between the cell velocity and the x‐axis (as a reference axis) (Figure S2), but there was no distinguished difference between the two strains, which may be due to relatively large fluctuations in the velocity calculations. Besides Δθ, the proportions of different twitching modes of cells were also measured (Figure 2F), as previous studies have shown that different twitching modes have different MSDs2, 33. The results showed that compared with WT^m^, ∆pilU ^m^ had a higher percentage of crawling‐only mode (70% ± 6% for ∆pilU ^m^ and 56% ± 5% for WT^m^) and a lower percentage of walking‐only and walking and crawling modes. Since walking motility often generates a random diffusive motion, low percentages of walking‐involved modes in ∆pilU ^m^ can help cells to maintain their moving direction. Taken together, these results suggest that compared with motile WT^m^ cells that showed a diffusive‐type surface motion, motile ∆pilU ^m^ cells had a reduced twitching activity, but their more confined Δθ and low percentages of walking‐involved twitching modes resulted in a more directional‐persistent surface motion characterized by a higher slope of MSDs.
*Effects of PilU loss on bacterial surface motility. (A) Examples showing trajectories of Pseudomonas aeruginosa WTm, ∆pilU m, and ∆pilT m. Different colors represent trajectories of different cells. (B) Mean square displacements (MSDs) measured from bacterial trajectories. The solid lines are regression lines of each MSD data set in the log–log plot. (C) Distributions of twitching speed. The twitching speed of each tracked cell at frame n was calculated by the displacement of the cell between nth and (n + 1)th frames divided by the corresponding time interval. N (WTm) = 7701, and N (∆pilU m) = 2708. (D) Fraction of movement time. N (WTm) = 49, and N (∆pilU m) = 33. (E) Distributions of Δθ, which is defined as the orientational angle difference of a cell between two consecutive frames. Curves are the Gaussian‐fitting results. N (WTm) = 81, and N (∆pilU m) = 81. (F) Distributions of twitching modes. A cell is classified in a crawling mode if its aspect ratio is ⩾2 and in a walking mode if its aspect ratio is <2. N (WTm) = 494, and N (∆pilU m) = 494. Error bars show standard deviations. Statistical significances were measured using a two‐sample Student's t‐test. *p < 0.05, **p < 0.01, and **p < 0.001.
Loss of PilU affects the microcolony formation time and morphology through reduced twitching and altered cell–cell collision responses
Biofilm formation is a classical collective behavior of cells after they attach on surfaces, during which TFP plays an important role34. The development of a normal biofilm requires not only the physical pilus itself but also its dynamic activity35. To understand whether and how PilU can affect biofilm development, we studied the dynamics of microcolony formation of the three strains, WT^m^, ΔpilU ^m^, and ΔpilT ^m^ (Figure 3). An earlier study has shown that microcolony formation was closely related to bacterial visit distributions on surfaces through Psl‐guided surface movement36, so we first examined the visit frequency map of cells obtained 4 h after bacterial inoculation (Figure 3A), which showed how often a pixel was visited by bacteria during the 4 h. The color scale in Figure 3A ranges from black to blue, representing the number of bacterial visits from 0 to 100. Among the three strains, WT^m^ had a relatively more uniform distribution while ∆pilT ^m^ had a more localized one and ∆pilU ^m^ was intermediate. Quantitatively, the bacterial visit frequency map can be characterized using a power law distribution36, and a more negative power law exponent would indicate a more uniform distribution. The results in Figure 3B showed that WT^m^, ∆pilU ^m^, and ∆pilT ^m^ had a power law bacterial visit distribution with an exponent of −2.87 ± 0.11, −2.69 ± 0.15, and −2.58 ± 0.11, respectively. Thus, compared to WT^m^, retraction motor mutants ∆pilU ^m^ and ∆pilT ^m^ exhibited a more hierarchical distribution of bacterial visits (Figure 3B). Among them, there was a significant difference between WT and ∆pilT ^m^ (p = 0.029), while there was no significant difference between WT and ∆pilU ^m^ (p = 0.128). Next, we measured the microcolony formation time defined as the time period from the inoculation of bacteria to observing the first microcolony in the field of view. In this study, a microcolony was defined as a cell aggregate consisting of ≥ 30 cells, the same definition used in the literature33. The results are shown in Figure 3C. The microcolony formation time was 3.5 ± 0.2 h for WT^m^, 4.7 ± 0.9 h for ∆pilU ^m^, and 5.1 ± 0.8 h for ∆pilT ^m^. These results indicated that the loss of retraction motors PilU and PilT would slow down the microcolony formation. Particularly, the loss of PilT (∆pilT ^m^) was slowed more than the loss of PilU (∆pilU ^m^). As the three strains showed similar growth curves (Figure S3), such differences in the microcolony formation time are likely caused by their motility differences. To further reveal the role of motility in the microcolony formation, we traced the cell lineages in the course of microcolony formation using the same method as in Zhao et al.36. The results are shown in Figure 3D, where different colors represent different lineages. The microcolony of WT^m^ consisted of cells with about 10 colors (i.e., 10 cell lineages) while the microcolony of ∆pilT ^m^ consisted of cells with only two colors. This is understandable, as WT^m^ cells could twitch and thus could move to join other cells to form microcolonies, while ∆pilT ^m^ cells could not twitch, so their microcolonies were formed from one or two cell lineages through cell multiplication. Similarly, ∆pilU ^m^ cells could twitch but had a reduced twitching activity compared with WT^m^; thus, the microcolony of ∆pilU ^m^ was composed of cells with seven colors. These results indicate that under our test conditions, cell twitching mobility can enrich the composition of microcolonies and slightly speed up the microcolony formation.
*Effects of PilU loss on the formation of microcolonies. (A) Distribution of bacterial visit frequencies measured 4 h after inoculation for the three strains, WTm, ΔpilU m, and ΔpilT m. (B) Power law distribution of bacterial visit frequencies shown in (A). (C) The 30‐cell microcolony formation time. (D) The composition of microcolonies formed by the three strains, WTm, ∆pilU m, and ∆pilT m. Cells in the images of the top row were color‐coded, with each color representing a different cell lineage determined from bacterial tracking. In each image, a microcolony was outlined by a white dotted line. The bottom row depicts more developed microcolonies at the same location 2 h later. Scale bars, 2 μm. Error bars show standard deviations. Statistical significances were measured using a two‐sample Student's t‐test. n.s., not significant; p < 0.05.
The microcolony formation is a multi‐cellular process, during which cell–cell interactions are critical for the final formed structures of microcolony. To explore the TFP‐mediated cell–cell interactions, we took advantage of the visualization of TFP and investigated what motile cells would respond when their TFP detected other cells (i.e., their TFP touched with other cells) along their moving trajectories. The results (Figure 4) showed that when cells collided with each other during their movement, generally they exhibited two types of response behavior, touch‐turn and touch‐upright (see schematic illustrations in Figure 4A). Touch‐turn refers to that a bacterial cell changes its orientation and movement direction (but in a plane parallel to the substrate) after the cell collides with another cell. Touch‐upright refers to that a cell becomes in a stand‐up configuration after it contacts another cell. Figure 4B shows one example for each type of response behavior in WT^m^ and ∆pilU ^m^ (see Movies S4–S7 in Supporting Information section for more details). By counting the percentage of cells that displayed touch‐turn and touch‐upright responses among all observed cell–cell collision events, we found that WT^m^ cells showed more touch‐turn responses than touch‐upright whereas ∆pilU ^m^ cells behave just the opposite (Figure 4C). Such differences in cell–cell collision responses between WT^m^ and ∆pilU ^m^ may contribute to the different morphology of microcolonies formed in these two strains (Figure 3D), where in a WT^m^ microcolony most of the cells lied down and thus the orientation of cells was aligned to a certain degree due to the crowd packing environment inside the microcolony while in a ∆pilU ^m^ microcolony quite a few cells stood up so cells orientated more randomly and thus made the microcolony a more‐disordered looking.
*Effects of PilU loss on cell–cell collision responses. (A) The two types of response behavior of cells when they contact with each other. (B) Examples of touch‐turn and touch‐upright responses in WTm and ∆pilU m. The interested cells are colored using the same color scheme as in the cartoon in (A), and the yellow lines show their trajectories from 0 min up to the current time point. Orange arrows indicate the moving direction of cells. (C) The measured fraction of touch‐turn and touch‐upright events among all observed cell–cell collisions. N (WTm) = 120 and N (∆pilU m) = 51. Error bars show standard deviations. Statistical significances were measured using a two‐sample Student's t‐test. **p < 0.001. Scale bar, 2 μm.
Loss of PilU results in a dense, multilayered colony edge and slows colony expansion
It has been shown that bacteria behave differently on substrates of varying hardness37, 38. As PilU is known to play an important role when cells require significant retraction forces22, 27, cells without PilU may behave differently on semi‐solid surfaces. To test this hypothesis, we studied the colony expansion of ∆pilU, ∆pilA, ∆pilT, and WT on semi‐solid agar surfaces. The results showed that the expansion edges of ∆pilU, ∆pilA, ∆pilT, and WT displayed different morphologies (Figure 5A). As a control, the expansion edges of WT were rough with relatively large interdigitated convex and concave regions. By contrast, the expansion edges of ∆pilU, ∆pilT, and ∆pilA were relatively smooth. On a macroscopic level, the colony edges of WT were uneven, while those of ∆pilU, ∆pilT, and ∆pilA were smooth (Figure S4). A closer examination of bacteria at the front of colony expansion further revealed that ∆pilT, ∆pilA, and WT all had a single layer of cells at the front of expansion while ∆pilU had a multi‐layered cellular structure (Figure 5A).
*Effects of PilU loss on the colony expansion on semi‐solid agar surfaces. (A) Images showing a part of the colony expansion edge of P. aeruginosa WT, ∆pilU, ∆pilT, and ∆pilA after 12 h incubation. Insets are magnified views of specified region at the expansion edge enclosed by yellow squares, in which cells at the edge were marked yellow. The yellow arrow at the bottom left corner of each image indicates the expansion direction. Scale bar: 10 μm. (B) The fraction of radial‐ and tangential‐orientated cells at the expansion edge. The results were obtained from a total of N field of views: N = 10 for WT, and the total count number of cells at the edge of community is 601; N = 14 for ∆pilU, and the total count number of cells is 650; N = 13 for ∆pilT, and the total count number of cells is 556; N = 10 for ∆pilA, and the total count number of cells is 619. (C) The expansion area of WT, ∆pilU, ∆pilT, and ∆pilA. (D) Distribution of fluorescence signals in the expansion colony of mixed strains. Snapshots were taken after 12 h incubation. Lines are guides to the eye, where the solid line indicates the edge of leading front and the dotted line indicates the boundary between the mixing region and the leading front region. The three graphs on the right are magnified views of specified region at the expansion edge enclosed by yellow squares. The yellow arrow at the bottom left corner of each image indicates the expansion direction. Scale bar: 10 μm. (E) Comparison of fluorescence intensity between the near‐boundary area of the mixing region and the leading front region. Statistical significances were measured using one‐way ANOVA. **, p < 0.001.
In addition, the distribution of cell orientations at the colony edge also showed different trend between WT and TFP mutants, as shown in Figure 5B. We can see that compared with WT, TFP mutants including ∆pilU, ∆pilA, and ∆pilT displayed an increase in the percentage of cells that orientated along the tangential direction of the colony edge and a decrease in the percentage of cells that orientated along the radial direction of the colony (i.e., from the colony center toward the periphery). Interestingly, we found that in ∆pilA and ∆pilT, cells at the colony edge were all in a lie‐down style (i.e., ~0% stand‐up cells), whereas in ∆pilU, more stand‐up cells (56% ± 13%) at the colony edge were observed (see examples in the enlarged windows enclosed by yellow rectangles in Figure 5A) (Figure S5). The stand‐up configuration of ∆pilU is very likely related to the touch‐upright responses of cells demonstrated in Figure 4, which may also contribute to the multilayered structure at the colony front in ∆pilU. All these differences in the structure and morphology of expansion front among the tested strains led to different speed of colony expansion with an order of WT >∆pilA >∆pilT >∆pilU (Figure 5C), which is also consistent with macroscopic observations (Figure S4).
To further understand the role of TFP‐mediated interactions in colony expansion, we mixed WT containing green fluorescent protein (GFP)‐expressing plasmids (WT‐GFP) with ∆pilU, ∆pilT, and ∆pilA, respectively. Each mixture had a ratio of WT‐GFP to TFP mutant of 1:5. The results are shown in Figure 5D. We can see that after the colony expanded for a certain time, stratification of cells was observed in all tested mixtures, which formed two regions: the leading front region and the mixing region (Figure 5D,E).
In the leading front region, by counting the cells within a 20 μm range from the front edge, we found that all cells were WT‐GFP cells and they essentially formed a monolayer. The dominance of WT cells in the leading front region was also confirmed by the results of a mixture of WT + ΔpilU‐GFP (Figure S6), where the colony expansion results showed that in the leading front region, there were no fluorescent cells observed (i.e., all cells were non‐fluorescent WT cells). These results suggest that cells with malfunctioned TFP expanded slower than WT, which is in consistent with the results shown in Figure 5C. Behind this WT‐dominated leading front region, cells of WT‐GFP and TFP mutants were mixed and formed a multilayered structure with high cell density. In this mixing region, the expansion was mainly due to cell growth and multiplication.
Interestingly, WT‐GFP cells in the mixing region displayed different distributions among different mixtures. In the WT‐GFP + ∆pilU mixture, the fluorescence intensity of the mixing region showed a non‐uniform distribution with much brighter signals near the boundary, indicating an accumulation of WT‐GFP cells at those locations (Figure 5D). Whereas in the WT‐GFP + ∆pilT mixture, the degree of non‐uniformity of the fluorescence intensity distribution was less although near‐boundary areas were still brighter compared with other areas in the mixing region. By contrast, in the WT‐GFP + ∆pilA mixture, the fluorescence intensity was more uniformly distributed, indicating that there was no accumulation of WT‐GFP cells near the boundary. Quantitatively, by normalizing the fluorescence intensity of the near‐boundary area by that of the leading front region, WT‐GFP + ∆pilU had the highest ratio (~2) (Figure 5E). Considering the observation that a colony of pure ∆pilU cells had a multilayered edge (Figure 5A), these results may suggest that ∆pilU cells can block the migration of WT cells toward the leading front region through forming a multilayered edge and thus lead to the accumulation of WT‐GFP cells near the boundary.
We note that the multilayered colony edge phenotype in ∆pilU was not due to the affected twitching motility per se, as such phenomena were not observed in either WT (normal twitching) or ∆pilA/∆pilT (non‐twitching). Together, all these results indicate that beyond affecting the twitching motility, PilU also plays a role in the regulation of cell colony expansions on semi‐solid surfaces.
DISCUSSION
Our study reveals multiple roles of PilU in regulating P. aeruginosa surface behaviors. We found that PilU influenced (1) TFP distribution and quantity, (2) single‐cell motility patterns, (3) cell–cell collision responses, and (4) collective behaviors in microcolony formation and colony expansion, which are summarized in Figure 6.
Summary of the effects of pilU on the surface behaviors of P. aeruginosa.
First, PilU affected cell morphology as the loss of PilU increased the TFP number on each cell. This is consistent with the hyperpiliated phenotype reported in the literature23, 39. Our results revealed that PilU played an important role in affecting the quantity of TFP, while PilT was more important in affecting the length of TFP. Consistent with the different impacts on cell morphology between PilU and PilT, they also resulted in different phenotypes with different cell twitching activities, where ∆pilT ^m^ could not twitch while ∆pilU ^m^ could. Such different twitching behavior suggests that the pilus retraction function of PilU cannot be separated from PilT, which also agrees with a recently established picture in the literature that in P. aeruginosa PilT is the main contractile protein while PilU acts as a supporting PilT‐dependent retraction motor22, 26, 27. We note that this conclusion may not be true in other species, as in D. nodosus it has been shown that mutation of either pilT or pilU eliminated cell ability to twitch24. Although ∆pilU ^m^ of P. aeruginosa could twitch, ∆pilU ^m^ cells exhibited slower and more directional‐persistent surface motion compared to WT^m^.
Second, in terms of collective behavior, our results showed that PilU affected microcolony formation in two ways. On one hand, the loss of PilU would slow down the twitching movement of cells and temporally lead to delayed microcolony formation. We note that this conclusion on the role of twitching motility in the formation of microcolonies does not conflict with the results of a previous study40, where the initial microcolonies (mushroom stalks) were formed by a clonal growth of non‐motile cells. First, the colony defined in this study (a cell cluster with a cell number bigger than 30) is different from the initial microcolonies in Klausen et al.40. The microcolony defined in this study happened earlier than that in Klausen et al.40. Second, for the cell cluster formed by WT cells, cells at the center will become less or even non‐motile due to the surrounding crowded environment, so these non‐motile cells will grow to form the mushroom stalks as shown in Klausen et al.40. A similar observation of PilU loss in delaying the microcolony formation was also reported in N. meningitidis 41. However, unlike the hyperpiliation of ΔpilU in P. aeruginosa, in the study of N. meningitidis 41, the electron microscopy measurements showed no difference in the morphology or piliation levels between ΔpilU and WT strains. Although there were no twitching measurements in this study, the delaying of microcolony formation was also observed in a liquid medium under which the twitching motility did not work41. Together, it seems to suggest that there might be different causes for the delaying of microcolony formation observed in P. aeruginosa and N. meningitidis.
Third, the loss of PilU also changed the TFP‐based cell–cell collision responses from touch‐turn dominance in WT to touch‐upright dominance in ΔpilU, which then resulted in different morphologies of microcolonies. The altered cell–cell interactions in ΔpilU changed significantly the colony expansion pattern of bacteria on semi‐solid agar surfaces, as illustrated in Figure 5, where ΔpilU cells formed a three‐dimensional multilayerd “fence”‐like edge that greatly reduced the expansion rate, whereas ∆pilT, ∆pilA, and WT all had a single layer of cells at the front of colony expansion. Similar differences in the colony edges were also found in a stab‐inoculated sub‐surface twitching assay, where pilU mutant was found to form a thick subsurface colony with a distinctive fringe whereas the pilA and pilT mutants formed colonies with smooth edges24. These results suggest that the “fence”‐like edge is likely due to the intrinsic properties of ΔpilU rather than substrate variations.
As ΔpilU cells were hyperpiliated and were also able to perform extension and retraction, we would like to know the passive role of TFP (i.e., physical filament itself) and the active role of TFP (i.e., through TFP extending and retracting activities) in these colony expansions. ΔpilT mutants were hyperpiliated but unable to retract TFP, so we could deduce the passive role of TFP from the measurements of ΔpilT which showed that TFP filaments can indeed slow down the colony expansion. This is consistent with the passive role of TFP found in the swarming behavior of P. aeruginosa 39. The mechanism of the passive role of TFP presumably arises from the adhesive function of TFP, which has been shown to play roles in surface attachment and cell aggregations1, 26, 34, 42. However, the passive role of TFP cannot explain the observation that the edge morphology of colony expansion of ΔpilT was more similar to that of both ΔpilA (no pili) and WT than that of ΔpilU (i.e., forming a monolayer of cells in ΔpilT, ΔpilA, and WT, but not in ΔpilU), suggesting that the active role of TFP in ΔpilU is more important for the formation of multilayered “fence”‐like edge. The increased touch‐upright responses in ΔpilU can contribute to the formation of “fence”‐like edge, as more upright cells would facilitate the formation of an “interception network” to prevent cells from passing through easily and thus more cells would accumulate near the edge, leading to a thick “fence”‐like edge.
However, the mechanism underlying the increased touch‐upright responses in ΔpilU is not clear. Our measurements of TFP distribution on cell surfaces showed that compared with WT^m^, ∆pilU ^m^ displayed a higher percentage of two‐poles events while ∆pilT ^m^ showed a reduced percentage. These results seem in agreement with Chiang's observations that PilT was found to be localized to both poles while PilU was localized at the piliated pole43. Thus, one possible scenario to increase the touch‐upright responses might be that loss of PilU would facilitate more cells having TFP located at both poles, which can create a tug‐of‐war situation when TFP at both poles retract simultaneously. Such a tug‐of‐war situation could be interrupted by cell collision, which leads to an imbalance in TFP retraction forces, causing cells to pivot into an upright position upon collision. We note that it is very likely that other factors would also be involved in TFP‐polarization, such as PilG and PilH, which have been shown to control the polarization of P. aeruginosa during mechanotaxis44. Future studies using high‐resolution microscopy techniques to visualize TFP dynamics during cell–cell interactions could help elucidate the precise mechanism underlying this touch‐upright phenotype. In addition, such different cell–cell interactions in ΔpilU may suggest that PilU can have an environmental sensing role that is independent of PilT and may function through cellular surface sensing systems such as the Pil‐Chp chemosensory system45. But more work is needed to test these hypotheses.
In conclusion, our results expanded our current understanding of the role of PilU in P. aeruginosa. Beyond that PilU acts as a PilT‐dependent retraction motor and is required for generating high retraction forces such as twitching in a typical agar stab assay23, 46, we further demonstrated the roles of PilU in cell surface behaviors. We showed that loss of PilU reduced cell twitching activity, delayed microcolony formation, altered cell response behavior after collision, and slowed down the colony expansion by forming a distinctive “fence”‐like colony edge. While our study provides new insight into PilU's roles beyond twitching, several questions remain. Future research should investigate the molecular mechanisms by which PilU influences TFP distribution and cell–cell interactions. Additionally, examining the impact of PilU on biofilm formation and virulence in animal models would further our understanding of its physiological importance.
MATERIALS AND METHODS
Bacterial strains and growth conditions
All strains and plasmids used in this study are listed in Table S1. Escherichia coli DH5α strain was used as a host for plasmid construction and amplification, see the literature32. P. aeruginosa strains were cultured on Luria‐Bertani (LB) agar plate at 37°C for 12 h, and appropriate antibiotics (200 μg/ml carbenicillin) were added if necessary. The culture was incubated by shaking in a FAB medium containing 30 mM glutamic acid (Sigma‐Aldrich) and selected concentrations of arabinose at 220 rpm and 37°C for about 6 h, until the OD_600_ of approximately 0.3. The culture was then diluted to OD_600_ of approximately 0.01 in FAB medium containing 0.6 mM glutamic acid and arabinose for injection into the flow chamber. For strains WT^m^, ∆pilU ^m^, and ∆pilT ^m^, 0.2% arabinose (Sigma‐Aldrich) was added to the medium to control the production of mutant PilA. A fully automatic inverted microscope is used to observe the movement of bacteria on glass surfaces.
Flow cell
Before inoculating bacteria into the flow tank, the FAB medium containing 0.6 mM glutamic acid was rinsed with a microinjection pump (Harvard Apparatus) at a flow rate of 30 ml/h for 5 min. The flow of the medium was subsequently halted, and a 1 ml syringe fitted with a needle was used to inject 1 ml of the bacterial suspension (with an OD_600_ of approximately 0.01) directly into the flow chamber. After inoculation, an incubation period of 5 min was allowed for the cells to attach to the surface, followed by 5 min of flushing at a high flow rate of 30 ml/h to wash away the floating cells. After that, the flow rate was set to 3 ml/h and the image recording was started. In this work, the flow cell experiment was carried out at 30°C.
Pili fluorescent staining
Pilin labeling was achieved using Alexa Fluor^TM^ F488 C5‐maleimide (Thermo Fisher Scientific, A10254). First, the injection pump was stopped, and 300 μl AF488‐mal solution with a concentration of 0.5 mg/ml was injected into the chamber through the injection port and immediately incubated in the dark for 15 min. The injection pump was turned on, and the fresh medium was rinsed at 30 ml/h for 5 min to remove the bound dye in the flow cell. The cell bodies and labeled TFP were then imaged using fluorescence microscopy on a Leica DMi8 microscope.
Data collection
Single‐cell tracking: An EMCCD camera (Andor iXon) was used to capture images on a Leica DMi8 microscope equipped with a zero‐drift autofocus system. The image size is 66.5 μm × 66.5 μm (1024 × 1024 pixels). These images were taken with a 100× oil lens (plus a 2× magnifying glass).
TFP phenotype and expansion rate data acquisition (fluorescence shooting): The images were recorded every 8 s. In order to reduce the influence of fluorescence irradiation on the physiological state of bacteria, the observation field of each sample was changed for 1 min of shooting, and the sample was changed for about 30 min of shooting.
Twitching movement data acquisition: Leica software was used to integrate equipment to record bright‐field images every 3 s, and the total recording time was about 12 h.
Group expansion tracking: The Nikon Ti2 inverted fluorescence microscope equipped with zero‐drift autofocus system and infrared camera Prime95B were used for bright‐field and fluorescent dual‐channel data acquisition. A 60× objective lens (plus 2× magnification) and NIS‐Elements Viewer software were used. The bright‐field images were recorded with one image every 3 s. The fluorescence images were recorded every 30 min, and the total recording time was about 12 h.
Single‐cell tracking image and data analysis
In this study, bacterial tracking was performed in the same way as in previous studies32, 43. Through bacterial tracking, a variety of bacterial information can be obtained, including cell centroid coordinates, cell orientations, cell instantaneous velocity, and MSDs of cells. The analysis of bacterial visit frequency was conducted employing the method in reference36.
When comparing the motility data of WT^m^ with ∆pilU ^m^ cells, it is necessary to first determine whether the cells are in a state of motion and then perform statistical analysis on the motile cells of the two strains. The threshold for determining cell movement or non‐movement was determined using the result of ∆pilT cells, as their speed is influenced by cell growth. ∆pilT cells have a measured speed distribution (from 100 cells) with an average speed of 0.41 ± 0.04 μm/min and a peak value of 0.32 μm/min, so a threshold of 0.45 μm/min was selected.
The determination of motile cells: A cell is defined as a motile cell if its movement speed is greater than 0.45 μm/min at any given moment during the time it appears in the field of view. If its speed remains below 0.45 μm/min throughout, it is defined as a non‐motile cell.
To determine the fraction of movement time of a cell, we first selected data within the first 3 h after cell adhesion to the surface and before the formation of biofilm microcolonies. Then, a specific cell was chosen, and if its instantaneous movement speed exceeded 0.45 μm/min, it was classified as motile; otherwise, it was classified as non‐motile. The ratio of the duration of cell movement to the total time the cell appears in the field of view is the proportion of time during which the cell is in motion. This ratio can be used to describe the level of cellular activity during the observation period.
For pili length measurements, pili that had retracted when image recording began were excluded from the analysis. For the measurement of pili elongation (contraction) speed, only cells that performed a single pilin elongation (contraction) within a 1‐min time window were analyzed.
The determination of cell touch‐turn or touch‐upright: When the pili of a cell touch another cell (cell body or pili), the cell changes its orientation and movement direction, but in a plane parallel to the substrate, then this phenomenon is defined as touch‐turn; when the pili of a cell touch another cell (cell body or pili), the cell becomes in stand‐up configuration, then this phenomenon is defined as touch‐upright.
The colony expansion experiments
The colony expansion experiments were performed in a similar way as described in Tylor and Welch47. In these experiments, 0.3% FAB agar–air interface was used to observe the expansion motion. The bacterial solution was cultured overnight to OD_600_ of approximately 1, and 0.5 μl was added to the center of the solidified medium, and then the bacterial solution was hung in the biosafety cabinet for 2 min, so that the surface water of the bacterial solution evaporated to facilitate observation. The device was put on a thin coverglass, and a 60x lens was used for observation. Images were recorded every 3 s. The fluorescence intensity of the near‐boundary area of mixing region refers to the average fluorescence intensity of the area within a range of 100 pixels away from the boundary of the mixing region, while the fluorescence intensity of the leading front region refers to the average fluorescence intensity of the area of the leading front region.
Measuring expansion rate: The bacterial expansion rate was calculated as the ratio of the total area visited by the bacteria divided by the time period during which bacteria expanded. In order to identify edges, the Sobel operator was used to convolve the image, followed by median filtering. Then, a threshold to segment the image was set and the area covered by bacteria was calculated.
For mixed cultures, in order to quantitatively determine the proportion of WT cells in the leading front region, we measured the percentage of WT cells within a 20 μm range from the front edge of the leading front region. For each experimental condition, a total of 10 images were selected and measured, and the counting results were subjected to statistical analysis to determine the WT cell proportion.
AUTHOR CONTRIBUTIONS
Jingchao Zhang: Conceptualization (equal); data curation (lead); formal analysis (equal); methodology (equal); project admin‐istration (equal); resources (lead); software (equal); supervision (equal); validation (lead); visualization (lead); writing‐original draft (equal); writing‐review and editing (equal). Yan Luo: Conceptualization (equal); data curation (lead); formal analysis (equal); methodology (equal); project administration (equal); resources (lead); software (equal); supervision(equal); writing‐original draft (equal). Yiwu Zong: Conceptualization (equal); data curation (equal); resources (equal); software (equal). Shangping Lu: Conceptualization (equal); resources (equal). Yi Shi: Conceptualization (equal); data curation (equal); resources (equal); software (equal); funding acquisition (lead); writing‐review and editing(lead). Fan Jin: Conceptualization (equal); data curation (equal); resources (equal); software (equal); funding acquisition (lead); writing‐review and editing (lead). Kun Zhao: Conceptualization (lead); Data curation (equal); formal analysis (equal); funding acquisition (lead); project administration (lead); writing‐original draft (lead); writing‐review and editing (lead).
ETHICS STATEMENT
This study did not involve any experiments on animals or humans.
CONFLICT OF INTERESTS
The authors declare no conflict of interests.
Supporting information
Supporting information.
Supporting information.
Supporting information.
Supporting information.
Supporting information.
Supporting information.
Supporting information.
Supporting information.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Ellison CK , Whitfield GB , Brun YV . Type IV Pili: dynamic bacterial nanomachines. FEMS Microbiol Rev. 2022;46:fuab 053.34788436 10.1093/femsre/fuab 053 · doi ↗ · pubmed ↗
- 2Gibiansky ML , Conrad JC , Jin F , Gordon VD , Motto DA , Mathewson MA , et al. Bacteria use type IV pili to walk upright and detach from surfaces. Science. 2010;330:197.20929769 10.1126/science.1194238 · doi ↗ · pubmed ↗
- 3Wall D , Kaiser D . Type IV pili and cell motility. Mol Microbiol. 1999;32:01–10.10.1046/j.1365-2958.1999.01339.x 10216854 · doi ↗ · pubmed ↗
- 4Merz AJ , So M , Sheetz MP . Pilus retraction powers bacterial twitching motility. Nature. 2000;407:98–102.10993081 10.1038/35024105 · doi ↗ · pubmed ↗
- 5Skerker JM , Berg HC . Direct observation of extension and retraction of type IV pili. Proc Natl Acad Sci USA. 2001;98:6901–6904.11381130 10.1073/pnas.121171698 PMC 34450 · doi ↗ · pubmed ↗
- 6Eriksson J , Eriksson OS , Maudsdotter L , Palm O , Engman J , Sarkissian T , et al. Characterization of motility and piliation in pathogenic Neisseria . BMC Microbiol. 2015;15:92.25925502 10.1186/s 12866-015-0424-6PMC 4449605 · doi ↗ · pubmed ↗
- 7Nakane D , Nishizaka T . Asymmetric distribution of type IV pili triggered by directional light in unicellular cyanobacteria. Proc Natl Acad Sci USA. 2017;114:6593–6598.28584115 10.1073/pnas.1702395114 PMC 5488941 · doi ↗ · pubmed ↗
- 8Mahmoud KK , Koval SF . Characterization of type IV pili in the life cycle of the predator bacterium Bdellovibrio . Microbiology. 2010;156:1040–1051.20056705 10.1099/mic.0.036137-0 · doi ↗ · pubmed ↗