snz/SNX25 at the crossroad of endocytosis and lipid handling in autophagy
Annie Lauzier, Steve Jean

TL;DR
This paper identifies SNX25 as a key regulator of autophagy and lipid handling in both flies and human cells.
Contribution
The study reveals SNX25's novel role in autophagy and its distinct domains for endocytosis and lipid regulation.
Findings
SNX25 is essential for autophagy in Drosophila and mammalian cells.
SNX25 regulates VAMP8 endocytosis and lipid handling through separate domains.
The sorting nexin family's role now includes autophagy regulation.
Abstract
Macroautophagy/autophagy is coupled to a myriad of intracellular processes, among which vesicular trafficking is an important contributor of membranes and proteins required at various stages, from autophagosome formation to degradation. Hence, understanding how membrane trafficking is coupled to autophagy induction and how cells rewire trafficking upon high autophagic needs is instrumental to our understanding of autophagy. In our recent manuscript, we tested a known class of endosomal sorting regulators, the sorting nexin family, for their involvement in autophagy in Drosophila. We identified snz (snazarus) as an important regulator of autophagy in Drosophila as well as in mammalian cells, by demonstrating a role for the snz human ortholog SNX25 in HeLa cells. Using knockout rescue experiments, we observed that SNX25 loss affects many cellular processes, namely VAMP8 endocytosis and…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
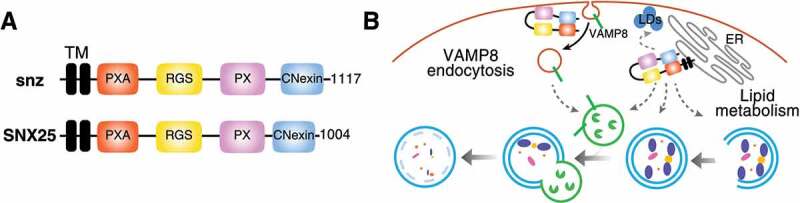 Figure 1
Figure 1 Figure 2
Figure 2- —CIHR (Canadian Institutes of Health Research
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsAutophagy in Disease and Therapy · Cellular transport and secretion · Toxoplasma gondii Research Studies
Autophagy relies on membranes and lipid influxes from a variety of organelles to feed phagophore expansion, leading to autophagosome formation. The phagophore and autophagosome lipid composition is also dynamically regulated during growth, maturation, and autophagosome-lysosome fusion. In addition to phagophores and autophagosomes, other vesicular compartments must account for and adapt to variations in autophagic needs. As such, vesicular trafficking of autophagic regulators must be tuned to demand, and mechanisms regulating this coupling are just being uncovered.
An important class of vesicular trafficking regulators is the sorting nexin (SNX) family. Through the phox homology (PX) domain, SNXs bind various phosphoinositide species, allowing their recruitment to intracellular membranes where they modulate endocytic and sorting events. Given their central roles in trafficking, different members affect autophagosome generation, but very few were identified as regulating terminal steps of autophagy. To test for SNX involvement in late stages of autophagy, we screened all Drosophila SNXs for their requirement in autolysosome formation [1]. From this focused RNAi screen, we identified the gene snz (snazarus) as an important regulator of autolysosome formation. Indeed, decreased expression of snz by RNAi or snz deletion in flies leads to ref(2)P and autophagosome accumulation. Together, these results demonstrate defective autophagy and decreased autophagosome clearance upon snz loss.
We then tested the four snz human paralogs, namely SNX13, SNX14, SNX19 and SNX25, in HeLa cells. We found that SNX14 and SNX25 loss, through RNAi or CRISPR-induced gene inactivation, result in autophagosome accumulation, indicating functional conservation. Because autophagosome buildup can be caused by defective endolysosomal compartments, we assessed endolysosome functions and overall endosome distribution in flies and HeLa cells upon snz or SNX25 loss, respectively. Endolysosomes are mostly unaffected, except for a slight increase in lysosome numbers in snz-depleted flies. This led us to test the localization of the Drosophila R-SNARE Vamp7 in flies and its ortholog VAMP8 in mammals, because Vamp7/VAMP8 trafficking is modulated by starvation and is important for autophagosome-lysosome fusion. Interestingly, we observed Vamp7/VAMP8 accumulation close to, or at, the plasma membrane in snz/SNX25-depleted cells and detected proximity between snz-Vamp7 and SNX25-VAMP8, suggesting a functional link.
The multi-domain-containing protein snz and its human orthologs (Fig. 1A) display specific functions linked to lipid droplet formation and lipid stress handling, which are conveyed through independent domains. To test a role for the individual domains, we performed rescue experiments by expressing SNX25 deletion mutants in SNX25 KO HeLa cells and monitored VAMP8 internalization. We focused on VAMP8, given its plasma membrane accumulation and reduced internalization in SNX25 KO cells. Surprisingly, the SNX25 TM domains are dispensable for VAMP8 internalization, whereas the PX domain is required. Mammalian SNX13, SNX14, and SNX25 PX domains have distinct preferences for phosphoinositides, with the snz PX domain more closely resembling the SNX25 PX domain. Considering the VAMP8 uptake defect upon SNX25 deletion, the SNX25 PX domain requirement for VAMP8 internalization and the preferred SNX25 PX domain interaction with di-phosphorylated phosphoinositides, we hypothesized that SNX25 can potentially affect endocytosis more generally. This hypothesis was not corroborated, and no effects are observed on clathrin-dependent and -independent endocytosis upon SNX25 loss. Figure 1.Potential roles of snz and SNX25 in autophagy. (A) Domain organization of snz and SNX25. (B) snz/SNX25 are required for proper Vamp7/VAMP8 internalization independently of endoplasmic reticulum (ER) localization (left part of the model). snz/SNX25 can potentially affect lipid exchange between the ER and endolysosomes or autophagosomes. snz/SNX25 can also modulate lipid droplet biogenesis, potentially affecting autophagy (right part of the panel).
Although VAMP8 uptake is important for autophagic flux, the possibility remained that SNX25 loss could disturb other processes, further affecting autophagy. We thus tested if the SNX25 TM or PX domains are required for autophagic flux. Our data demonstrated that, autophagic flux is rescued by expressing either SNX25 PX- or TM-deleted mutants in SNX25 KO cells. These results indicated that a SNX25 TM mutant unable to rescue VAMP8 endocytosis can still rescue the autophagy defects caused by SNX25 deletion. Thus, additional processes are impaired by SNX25 loss.
The SNX13, SNX14, SNX19 and SNX25 subfamily has well-known roles in lipid droplet formation and metabolism. Moreover, SNX14 deletion affects the relative ratio between phosphatidylcholine and phosphatidylserine. Thus, we performed a rescue experiment with ethanolamine (ETA) to test whether increased intracellular phosphatidylcholine concentrations can rescue SNX25 loss. Using various ETA concentrations, we rescued SNX25 loss of function. These results suggest that SNX25 influences both VAMP8 internalization as well as cellular lipid metabolism (Fig. 1B).
Altogether, our manuscript uncovered a conserved role for snz and SNX25 in the regulation of autophagy through modulation of Vamp7/VAMP8 endocytosis as well as lipid metabolism. Given our ability to rescue SNX25 loss with different SNX25 deletion mutants or by ethanolamine supplementation, we posit that distinct SNX25 domains may partially compensate for SNX25 loss by modifying specific metabolic or trafficking pathways. Accordingly, reintroducing a partially functioning mutant or modulating specific lipid species abundance rescue *SNX25-*deleted cells. How homeostatic conditions or cellular stressors regulate snz and the SNX25 subfamily, Vamp7/VAMP8 endocytosis, and lipid handling, will be a truly interesting area of research.
The reference list from the paper itself. Each links out to its DOI / PubMed record.