Exploring Aeromonas veronii in Migratory Mute Swans (Cygnus olor): A Debut Report and Genetic Characterization
Zhifeng Peng, Chunyan Gao, Hongxing Qiao, Han Zhang, Huimin Huang, Yamin Sheng, Xiaojie Zhang, Baojun Li, Baoliang Chao, Jingjing Kang, Chuanzhou Bian

TL;DR
This study reports the first isolation of Aeromonas veronii from migratory mute swans in China, showing it can cause disease and is resistant to multiple antibiotics.
Contribution
First documentation of Aeromonas veronii in migratory mute swans, expanding its known host range and highlighting its zoonotic potential.
Findings
A. veronii strain HNZZ-1/2022 was isolated from a dead migratory mute swan and shown to cause disease in goslings.
The strain exhibited multi-drug resistance to meropenem, ampicillin, and enrofloxacin.
It carried six virulence genes and had an LD50 of 3.48 × 10⁸ CFUs/mL in goslings.
Abstract
Aeromonas veronii (A. veronii) can cause serious disease in humans and various animals and is ubiquitous in terrestrial and aquatic milieus. Therefore, A. veronii has recently been considered to be an emerging pathogen worldwide. However, there is no documentation of clinical symptoms and pathological changes in A. veronii causing disease in migratory mute swans. In this study, we isolated and identified an A. veronii strain from migratory mute swans in China for the first time. We further explored its pathogenicity and antimicrobial susceptibility. Our results indicated that the newly identified strain has a strong ability to cause disease and exhibits multi-drug resistance. This is the first report of A. veronii being isolated from a migratory mute swan, which expands its known host spectrum. These findings not only help us to better understand the zoonotic potential of A. veronii but…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
 Figure 1
Figure 1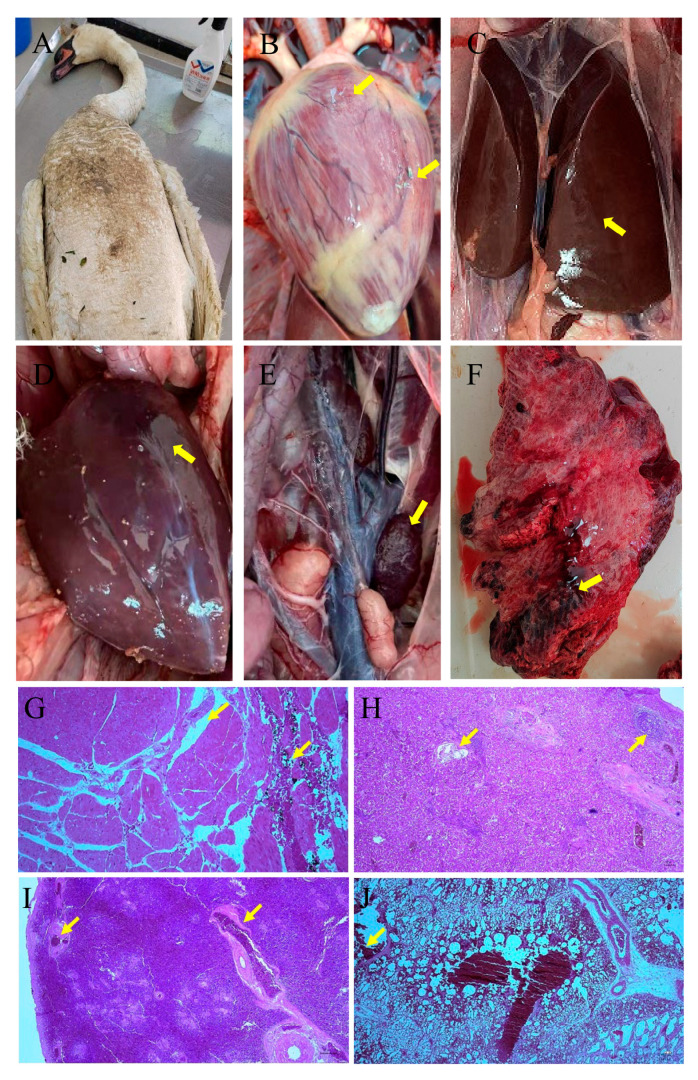 Figure 2
Figure 2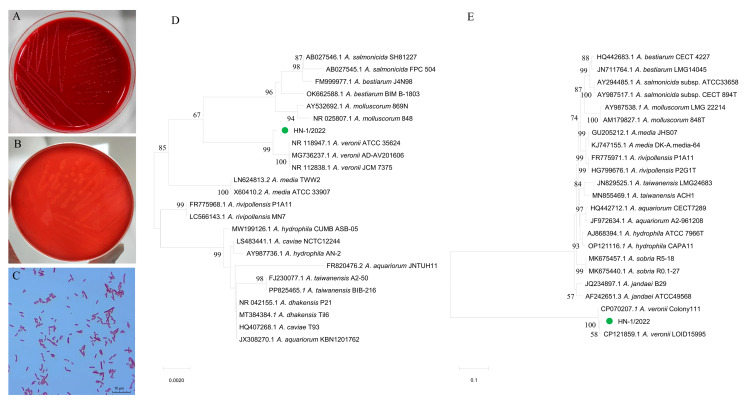 Figure 3
Figure 3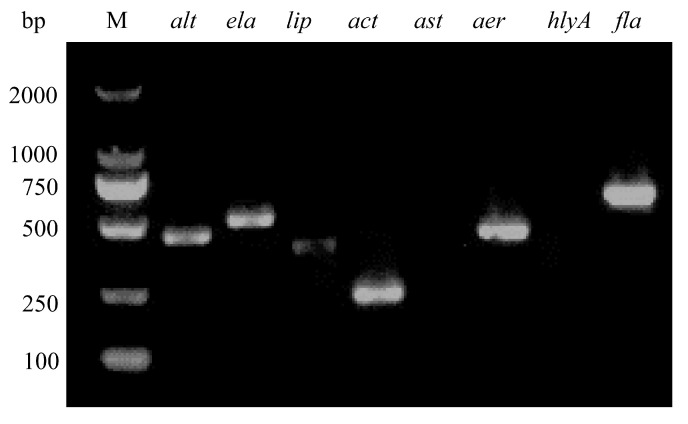 Figure 4
Figure 4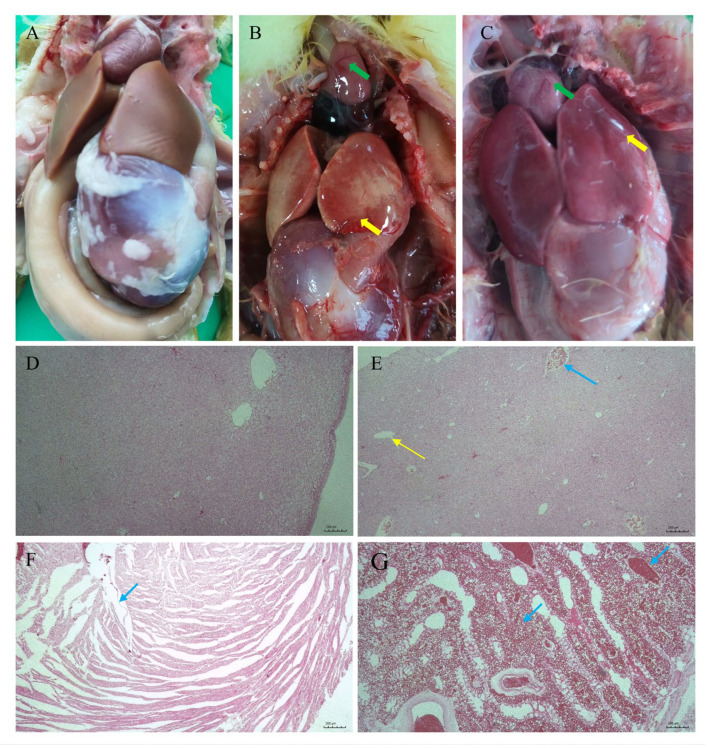 Figure 5
Figure 5- —Key Discipline of Veterinary Medicine of Henan Province
- —Key Discipline of Preventive Veterinary Medicine of Henan University of Animal Husbandry and Economy
- —Henan Province key research and development project
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsAquaculture disease management and microbiota · Fish Ecology and Management Studies · Aquaculture Nutrition and Growth
1. Introduction
The 36 species of the genus Aeromonas are Gram-negative, facultatively anaerobic, non-spore-forming, and catalase-positive bacilli, which are widely distributed in municipal and hospital wastewater [1,2], lake water [3], river water, oceanic water, irrigation water, and regenerated wastewater [4,5]. While some species are opportunistic pathogens of humans [6,7], they also affect a variety of animals, including reptiles, amphibians, fish, and aquatic animals [8,9]. Aeromonas species can cause hepatobiliary tract, urinary tract, soft tissue, and skin infections, as well as pneumonias, peritonitis, and severe septicemia [6,7,9]. The pathogenic potential of Aeromonas is associated with several virulence factors, including aerolysin (aerA), hemolysin A (hlyA), heat-stable cytotonic toxins (ast), heat-labile cytotonic enterotoxin (alt), and aerolysin-related cytotoxic enterotoxin (act) [10,11]. In addition, the lipase (lip), lateral elastase (ela), polar flagellum (fla), flagella (laf), and type III secretion system (TTSS) contribute to the virulence of the genus Aeromonas [12,13]. The above-mentioned virulence factors allow Aeromonas bacteria to adhere, invade, and escape the host’s immune response [14,15].
As an important species of the genus Aermonas, A. veronii is pathogenic to various aquatic animals and terrestrial species. This bacterium has been isolated from sheep exhibiting respiratory symptoms [16], dead Yangtze finless porpoise in China [17], Macrobrachium nipponense with red gill disease in China [18], diseased Trionyx sinensis in China [19], diseased Lateolabrax maculatus in China [20], various freshwater fish species in Malaysia [21] and Thailand [22], marketed Tegillarca granosa in Korea [23], channel catfish in the United States [24], and farmed Channa micropeltes in Thailand [25]. More importantly, A. veronii infections have caused serious disease in humans, particularly in the elderly and children, causing sepsis, gastroenteritis, and other conditions [26,27]. It is a dominant bacterial pathogen in freshwater fish worldwide [21,28], and A. veronii is the most prevalent Aeromonas species within the genus Aeromonas in food products in Shanghai, China [29]. Therefore, A. veronii has become a dominant pathogenic species within the genus Aermonas in recent years. Additionally, humans and animals are mainly infected with A. veronii through the ingestion of contaminated water or food, exposure to fecal contamination, and direct contact [30,31]. All of the aforementioned studies highlight that A. veronii is involved not only in animal health and environmental health, but also human health and food safety, posing a serious threat to the One Health approach [27,28,31,32]. Consequently, A. veronii has been considered an emerging zoonotic bacterial pathogen worldwide [32].
The migratory mute swan is the most abundant of the swan species, with a total of seven populations distributed worldwide, comprising over 600,000 individuals [33]. They help regulate vegetation growth by foraging on aquatic vegetation, which affects water quality and provides habitats for other species. In addition, their aggressive territorial behavior has led endangered species like terns and black predators to abandon their nests [34]. In East Asia, the migratory mute swan migrates in a timed and directional pattern, stopping at specific stopover sites along three flyways among China, Russia, Mongolia, North Korea, South Korea, and Japan [35]. The presence of multi-drug-resistant zoonotic bacteria of various genus and different kinds of viruses in migratory mute swans enables them to potentially transmit zoonotic pathogens and drug-resistance genes over long distances, thus posing a potential threat to public health [35,36,37].
The prevalence of Aeromonas spp. in several migratory birds in China has been reported [36]. However, there is no documentation of clinical symptoms and pathological changes in A. veronii causing disease in migratory mute swans (Cygnus olor). In this study, A. veronii was first identified as causative agent of multi-organ lesions in migratory mute swan that died in China. Its pathogenicity and antimicrobial resistance were characterized, respectively. Our findings expand the list of hosts for A. veronii to include migratory mute swans, and the possible transmission routes and public health significance of A. veronii are also discussed.
2. Materials and Methods
2.1. Case Presentation and Pathological Examination
Many mute swans and various species of water birds inhabit Sanmenxia Swan Lake National Urban Wetland Park in the city of Sanmenxia, Henan province, China (Figure 1). In April 2022, several mute swans died successively without obvious clinical symptoms. A fresh corpse of mute swan was transported to the Veterinary Diagnostic Laboratory, Henan University of Animal Husbandry and Economy, for post-mortem examination. The mute swan weighed 12.8 kg and showed no wounds on the body surface (Figure 2A). After disinfection of the mute swan with 75% ethanol, a post-mortem examination was conducted, and various internal organs (spleen, lung, liver, heart, and kidney) were sampled aseptically for laboratory examinations. Tissues fixed with neutral 10% formalin were embedded in paraffin and then manually sectioned with a microtome to obtain 4–5 μm thick paraffin sections. Then, dewaxed sections were stained with hematoxylin and eosin (HE) for light microscopy. This study was approved by the Research Ethics Committee of Henan University of Animal Husbandry and Economy (approval number HNUAHE ER2324121).
2.2. Bacteria Culture and Molecular Detection of Virus
The visceral organs were streaked onto brain–heart infusion (BHI) plates with 5% sheep blood (AOBOX, Beijing, China), blood agar plates with 5% sheep blood (Biocell, Zhengzhou, China), MacConkey (MAC) plates (AOBOX, Beijing, China), and Trypticase soy agar (TSA) plates with 5% sheep blood (AOBOX, Beijing, China), respectively. After incubation at 37 °C for 24 h, single-cultured colonies were selected for bacteriological culture, Gram staining, and microscopic examination [38]. Then, the tissue samples were pooled and homogenized in phosphate-buffered saline (PBS, pH 7.2). After being freeze–thawed three times, the homogenized tissue samples were centrifuged at 8000× g for 10 min. Viral DNA/RNA was extracted from the supernatant using a Takara Virus DNA/RNA Kit (Takara, Dalian, China) according to the manufacturer’s instructions. The DNA/RNA samples were used to detect goose reovirus (GRV), goose hemorrhagic polyomavirus (GHPV), goose parvovirus (GPV), Tembusu virus (TMUV) and avian influenza virus (AIV) with specific primers as described previously [39,40,41] (Supplementary Table S1).
2.3. Protein Analysis by MALDI-TOF MS
According to the manufacturer’s instructions, the single-cultured colonies were chosen for species identification using MALDI-TOF MS (Bruker, Billerica, MA, USA) [42]. Approximately 1 μL of each extract was prepared and mixed with α-cyano-4-hydroxycinnamic acid (Bruker Daltonic, Billerica, MA, USA), spotted onto a 96 polished steel targets plate, and dried at room temperature. Mass spectrometry was performed with a MALDI Biotyper system with the microflex^®^LRF software V3.0 (Brucker Daltonics). Spectra ranging from a mass-to-charge ratio (m/z) of 2000 to 20,000 were determined using the MALDI Biotyper system the V4.1 software and library (MALDI Biotyper database, 8468 entries, Bruker Daltonics).
2.4. Sequence Analysis of 16S rRNA and gyrB Genes
The genomic DNA of the newly identified strain was extracted using a genomic DNA purification kit (Tiangen Biotech, Beijing, China) following the manufacturer’s guidelines and then stored at −20 °C. The 16S rRNA was amplified using universal primers 27F (5′-AGAGTTTGATCCTGGCTCAG-3′) and 1492R (5′-GGCTACCTTGTTACGACTT-3′) and a PCR amplification procedure as described previously [43]. The gyrB gene was amplified using the universal primers gyrB-F (5′-GAAGTCATC ATGACCGTTCTGCAYGCNGGNGGNAARTTYGA-3′) and gyrB-R (5′-AGCAGGGTACGGATGTGCGAGCCRTCNACRTCNGCRTCNGTCAT-3′) and a PCR amplification procedure as described previously [44]. The 16S rRNA and gyrB gene sequences were sequenced by SUNYA Biotech (Zhengzhou, China). Further molecular identification of the isolated strains was performed through phylogenetic analysis of the 16S rRNA and gyrB genes. Phylogenetic trees were constructed based on 16S rRNA gene and gyrB gene sequences and analyzed using the MEGA-X software with the neighbor-joining method and 1000 bootstrap replicates, respectively.
2.5. Detection of Virulence-Associated Genes
Total DNA extracted from the newly identified Aeromonas isolate was used as PCR templates for virulence gene detection. Virulence genes aerA, hlyA, fla, alt, ast, act, lip, and ela were selected as virulence markers. The virulence genes were detected using the primers (Supplementary Table S2) and PCR amplification procedure as described previously [38]. Analysis of PCR products by 1% agarose gel electrophoresis was performed using a DL 2000 DNA marker (Takara, Dalian, China).
2.6. Pathogenicity Test on Goslings
Twenty-five healthy, one-day-old goslings were obtained from Yuanyang Yunbo goose industry company. Goose reovirus (GRV), goose hemorrhagic polyomavirus (GHPV), goose parvovirus (GPV), Tembusu virus (TMUV) and avian influenza virus (AIV) were excluded using specific primers as described previously [40,41]. To acclimate the laboratory environment, all goslings were housed in a pathogen-free environment for three days before the experiment initiation. Four groups of goslings (five goslings per group) were inoculated intraperitoneally with the newly identified strain at doses of 1.1 × 10^9^, 1.1 × 10^8^, 1.1 × 10^7^, and 1.1 × 10^6^ CFU/gosling (0.2 mL/gosling), respectively. One control group was inoculated with 0.2 mL sterile PBS per gosling. The mortality and symptoms of the goslings were recorded daily for 7 days. The LD_50_ value was calculated by the Reed and Muench method. Three dead goslings were randomly selected for bacteriological detection and pathological examination. The soft tissues fixed with neutral 10% formalin were embedded in paraffin and then manually sectioned with a microtome to obtain 4–5 μm thick paraffin sections. Then, dewaxed sections were stained with HE for light microscopy. Re-isolation and identification of the bacteria from the hearts, livers, and spleens of dead goslings were carried out.
2.7. Antimicrobial Susceptibility Testing
The antimicrobial susceptibility was evaluated using the microbroth dilution method according to the Clinical and Laboratory Standards Institute guidelines (CLSI, 2020). Briefly, a single colony of the newly identified Aeromonas isolate was inoculated onto Mueller–Hinton (MH) agar and incubated at 35 ± 1 °C for 24 h. A 0.5 McFarland turbidity standard (5 × 10^5^ CFU/mL) bacterial suspension was prepared with MH broth. Then, 50 µL of the bacterial suspension was added to each well of the microplate. Two wells containing only bacterial suspension were used as a positive control, and two wells containing only sterile cation-adjusted MH broth were used as a negative control. The microplates were incubated at 35 ± 1 °C for 24 ± 2 h. Escherichia coli ATCC 25922 was used as a quality control strain in all tests. The following antibiotics (Solarbio, Beijing, China) were tested: meropenem (0.25–128 µg/mL), linezolid (0.25–128 µg/mL), ampicillin (0.25–128 µg/mL), doxycycline (0.0625–32 µg/mL), tigecycline (0.25–128 µg/mL), colistin (0.25–128 µg/mL), gentamicin (0.25–128 µg/mL), florfenicol (0.125–64 µg/mL), enrofloxacin (0.3125–160 µg/mL), cefoxitin (0.25–128 µg/mL), ceftazidime (0.25–128 µg/mL), tiamulin (1–512 µg/mL), spectinomycin (0.25–128 µg/mL), and Fosfomycin (0.25–128 µg/mL).
3. Results
3.1. The Dead Mute Swan Exhibited Acute Lesions in Multiple Organs
After necropsy, multiple organs of the dead mute swan exhibited acute lesions, including degeneration and hemorrhage in the myocardium (Figure 2B), swelling in the liver (Figure 2C), hemorrhage in the spleen (Figure 2D), hemorrhage and swelling in the kidneys (Figure 2E), and congestion and necrosis in the lungs (Figure 2F). Based on hemorrhage in multiple organs, the cause of death of the mute swan was initially diagnosed as bacteremia and septicemia. The histological examination of the dead mute swan showed inflammatory cell infiltration, vacuolar degeneration, and extensive hemorrhage in multiple organs. The myocardial extracellular spaces were widened, with a large number of inflammatory cells infiltrating (Figure 2G). The structure of hepatic cords was unclear, and local necrosis was infiltrated by inflammatory cells (Figure 2H). The spleen exhibited capillary congestion (Figure 2I). The alveolar interstitium was widened, with a large number of inflammatory cells infiltrating, and a great quantity of red blood cells was found in the alveolar interstitium (Figure 2J).
3.2. The Isolated Strain HNZZ-1/2022 Was a Gram-Negative, Short Rod with Blunt Ends
The aseptically collected tissues were inoculated on TSA plates, MAC plates, BHI plates, and sheep blood plates, respectively, for bacterial isolation. The results showed that the dominate bacteria, termed HNZZ-1/2022, was cultured from plates inoculated with the heart blood, spleen, lung, and liver tissue. The colonies of HNZZ-1/2022 were observed to be smooth, round, and pale grey in color, with a diameter of 2 mm (Figure 3A). The newly isolated strain HNZZ-1/2022 exhibited β-hemolysis on sheep blood plates (Figure 3B), and it was Gram-negative, arranged singly or doubly, and was a short rod with blunt ends (Figure 3C).
3.3. The Strain HNZZ-1/2022 Was Identified as A. veronii
In the protein analysis by MALDI-TOF MS, the strain HNZZ-1/2022 best matched with A. veronii with a score value of 2.255. The 16S rRNA gene of the isolated bacterial strain HNZZ-1/2022 was 1459 bp in length (Supplementary Figure S1; GenBank accession number: OQ860819). It was 99.6–100.0% identical to A. veronii ATCC 35624 (NR_118947.1), A. veronii AD-AV201606 (MG736237.1), and A. veronii JCM 7375 (NR_112838.1). The gyrB gene was 1191 bp in length (Supplementary Figure S1; GenBank accession number: PP798204), and it was 98.3–99.4% identical to A. veronii LOID15995 (CP121859.1) and A. veronii Colony111 (CP070207.1). In the phylogenetic trees constructed based on the 16S rRNA sequence (Figure 3D) and gyrB sequence (Figure 3E), the isolated strain HNZZ-1/2022 was clearly grouped with a cluster of known A. veronii strains. Based on the morphology, MALDI-TOF testing, and analysis of the 16S rRNA and gyrB gene sequences, the isolated strain HNZZ-1/2022 was identified as A. veronii.
3.4. Strain HNZZ-1/2022 Contained Six Virulence-Associated Genes
To evaluate the virulence of A. veronii strain HNZZ-1/2022, eight virulence-associated genes were detected by PCR. The results indicated that A. veronii HNZZ-1/2022 contained six virulence-associated genes, including genes encoding for heat-labile cytotoxic enterotoxin (alt), elastase (ela), lipase (lip), cytotoxic enterotoxin (act), aerolysin (aerA), and polar flagella (fla) (Figure 4).
3.5. A. veronii Strain HNZZ-1/2022 Caused Multiple-Organ Pathological Changes in Goslings
Four groups of goslings (five goslings per group) were intraperitoneally inoculated with the newly identified strain with doses of 1.1 × 10^9^, 1.1 × 10^8^, 1.1 × 10^7^, and 1.1 × 10^6^ CFU/gosling (0.2 mL/gosling), respectively. The goslings exhibited various symptoms, such as lethargy, tremor, and tachypnea, within 12 h post inoculation, which were similar to those in the dead mute swan. The goslings began to die 24 h post inoculation, and all the goslings in the two highest-dose inoculation groups died within 2 days. The mortality rates of the other three groups inoculated with doses of 1.1 × 10^7^ and 1.1 × 10^6^ CFU per gosling within 7 days were 0% and 0%, respectively. The LD_50_ value of A. veronii strain HNZZ-1/2022 was estimated to be 3.48 × 10^8^ CFU/mL for goslings (Table 1). No obvious pathological lesions were observed in the negative control group (Figure 5A,D). Pathological lesions in the heart, liver, and lung were similar to those in the dead mute swan (Figure 5B,C,E–G). Additionally, according to the sequence analysis of the 16S rRNA gene and gyrB gene, the recovered isolate from the infected goslings was identified as A. veronii.
3.6. Strain HNZZ-1/2022 Is Multi-Drug-Resistant
The antibiotic susceptibility test of the A. veronii HNZZ-1/2022 strain using the microbroth dilution method using 14 antibiotics showed that HNZZ-1/2022 was sensitive to cefoxitin (8 µg/mL) and gentamicin (2.5 µg/mL) and resistant to meropenem (32 µg/mL), ampicillin (32 µg/mL), and enrofloxacin (0.625 µg/mL) (Supplementary Table S3).
4. Discussion
To date, A. veronii has been reported as a causative pathogen in various species of animals, including freshwater fish, amphibians, birds, aquatic animals, and reptiles [8,9,45,46]. A. veronii has caused various diseases in humans worldwide, and it has become the dominant Aeromonas species among clinical isolates [21,24,28,47,48,49]. Additionally, it is involved in food safety and public health concerns [28,29,32]. Given these facts, A. veronii has become an increasingly recognized zoonotic pathogen due to its isolation from various clinical and environmental samples worldwide [3,32,50]. In the present study, A. veronii was isolated as the causative agent from a dead migratory mute swan exhibiting acute lesions in multiple organs, such as hemorrhage and necrosis. To our knowledge, this is the first report of a fatal A. veronii infection in migratory mute swans, thus expanding the known host spectrum of A. veronii, which was traditionally limited to aquatic animals and humans [12,26,27].
Previous studies have shown that the virulence phenotype is a cumulative effect of multiple pathogenic factors [51]. In this study, A. veronii HNZZ-1/2022 contained six virulence genes, including genes encoding for heat-labile cytotoxic enterotoxin (alt), elastase (ela), lipase (lip), cytotoxic enterotoxin (act), aerolysin (aerA), and polar flagella (fla). These virulence genes encode proteins and are considered to be major factors affecting the pathogenicity of the genus Aeromonas. The presence of polar flagella (fla) in the genus Aeromonas confers rapid mobility, allowing the bacteria move on solid surfaces and form biofilms [10]. Additionally, the cytotoxic enterotoxin (act) restrains the phagocytosis activity of host cells and produces hemolysis [52]. The aerolysin encoded by the aerA gene is the prototype hemolysin of the genus Aeromonas, which can form pores in the target cell membrane and lead to osmotic lysis [10]. In this study, the LD_50_ value of the A. veronii strain HNZZ-1/2022 was estimated to be 3.48 × 10^8^ CFU/mL for goslings. These findings indicate that HNZZ-1/2022 is highly pathogenic.
With the increasing reports of multi-drug-resistant A. veronii, antimicrobial resistance has become an increasingly concern in humans as well as animals [24,29,50]. Aeromonas species isolated from aquatic organisms, insects, chicken, and humans in India show similar antimicrobial resistance profiles [53]. Previous studies indicated that most A. veronii strains are resistant to amoxicillin, ampicillin, and penicillin, regardless of whether isolated from humans or animals [8,54]. In the current study, A. veronii HNZZ-1/2022 isolated from migratory mute swan exhibited resistance to meropenem, ampicillin, and enrofloxacin. Carbapenems are considered critically important antimicrobials for human medicine by the World Health Organization. Notably, carbapenemase-producing A. veronii has been reported in the environment of an equine veterinary hospital in the USA [50]. Additionally, recent studies indicated that A. veronii strains isolated from the urban-impacted Akaki river in Ethiopia were resistance to imipenem (75.5%) and meropenem (63.3%), with 94.4% of carbapenem-resistant A. veronii strains being carbapenemase producers [55]. Furthermore, almost all A. veroni strains isolated from municipal and untreated hospital wastewater were also resistant to imipenem and meropenem [2]. In the current study, the newly identified A. veronii HNZZ-1/2022 isolated from migratory mute swans exhibited resistance to meropenem. The emergence of bacterial resistance to carbapenem antibiotics poses a significant challenge to public health, as carbapenems are a last-resort treatment option for life-threatening infections. Resistance to these drugs would reduce the effectiveness of infection treatment and increase medical costs. Moreover, A. veronii HNZZ-1/2022 showed penicillin resistance, which is consistent with previous reports that Aeromonads are intrinsically resistant to β-lactams [22].
Although we cannot confirm the sources of the bacteria in this case, it is possible that A. veronii was horizontally transmitted through contaminated water or food. There is a possibility that the mute swans were exposed to the bacteria via contaminated water or fish raised in the lake. Additionally, tourists sometimes feed mute swan sweet potatoes and other foods, which may also serve as a source of infection. Alternatively, the mute swan may have been infected through ingestion of food contaminated with A. veronii. The data obtained in the current study suggest that the migratory mute swan is a natural reservoir of A. veronii, implying mute swans can transmit the pathogen over long distances. In East Asia, migratory mute swans migrate along three routes connecting China, Russia, Mongolia, North Korea, South Korea, and Japan. During migration, mute swans stop at various parks, rivers, and freshwater lakes such as Qinghai Lake, Valley of the Lakes, Hulun Lake, Yellow River, Selenga River, Har Us Nuur National Park, and Khogno Khan National Park [35]. These findings suggest that A. veronii can be transmitted to many areas containing bodies of water by migratory mute swans, leading to water contamination [36]. Humans can be infected with A. veronii through direct and indirect contact with the migratory mute swans or through drinking A. veronii-contaminated water. Meanwhile, fish obtained from contaminated water can cause foodborne transmission of A. veronii in humans [29,32]. Furthermore, farmed waterfowl can be directly and indirectly infected with carbapenem-resistant A. veronii through drinking contaminated water, potentially transmitting the bacteria to humans. Therefore, foodborne and waterborne transmission of A. veronii is a public health concern that should receive more attention. This calls for continuous monitoring of A. veronii from various sources for One Health mitigation as well as improved environmental sanitation for farmed waterfowl to prevent contamination of water and human infections. It also highlights the need for caution when using carbapenems in the clinical treatment of A. veronii infections.
5. Conclusions
To the best of our knowledge, this is the first report of A. veronii being isolated from a migratory mute swan in China, further expanding its known host spectrum. A gosling pathogenicity test showed that A. veronii HNZZ-1/2022 could cause goslings to die. Moreover, it exhibits resistance to meropenem, ampicillin, and enrofloxacin. These findings suggest its strong zoonotic potential. However, in the current study, only one A. veronii strain isolated from a migratory mute swan in Sanmenxia Swan Lake National Urban Wetland Park was studied, and the mechanism of its antibiotic resistance was not studied deeply. Hence, much more effort will be focused on the active surveillance of A. veronii, its antibiotic resistance, and its transmission mechanism to better understand the epidemiological characteristics of A. veronii and guide the management of future human and animal infections.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Yang C. Guo M. Yang H. Wen Y. Zhu Z. Wang T. Zhu J. Chen L. bla(KPC-24)-Harboring Aeromonas veronii from the Hospital Sewage Samples in China Microbiol. Spectr.202210 e 005552210.1128/spectrum.00555-2235546572 PMC 9241592 · doi ↗ · pubmed ↗
- 2Drk S. Puljko A. Dželalija M. Udiković-KolićN. Characterization of Third Generation Cephalosporin- and Carbapenem-Resistant Aeromonas Isolates from Municipal and Hospital Wastewater Antibiotics 20231251310.3390/antibiotics 1203051336978380 PMC 10044312 · doi ↗ · pubmed ↗
- 3Xu X. Fu H. Wan G. Huang J. Zhou Z. Rao Y. Liu L. Wen C. Prevalence and genetic diversity of Aeromonas veronii isolated from aquaculture systems in the Poyang Lake area, China Front. Microbiol.202213104200710.3389/fmicb.2022.104200736578578 PMC 9791064 · doi ↗ · pubmed ↗
- 4Guerra R.M. Maleno F.D. Figueras M.J. Pujol-Bajador I. Fernández-Bravo A. Potential Pathogenicity of Aeromonas spp. Recovered in River Water, Soil, and Vegetation from a Natural Recreational Area Pathogens 202211138210.3390/pathogens 1111138236422633 PMC 9696040 · doi ↗ · pubmed ↗
- 5Rusiñol M. Hundesa A. Cárdenas-Youngs Y. Fernández-Bravo A. Pérez-Cataluña A. Moreno-Mesonero L. Moreno Y. Calvo M. Alonso J.L. Figueras M.J. Microbiological contamination of conventional and reclaimed irrigation water: Evaluation and management measures Sci. Total Environ.202071013629810.1016/j.scitotenv.2019.13629831923670 · doi ↗ · pubmed ↗
- 6Bhowmick U.D. Bhattacharjee S. Bacteriological, Clinical and Virulence Aspects of Aeromonas-associated Diseases in Humans Pol. J. Microbiol.20186713714910.21307/pjm-2018-02030015452 PMC 7256846 · doi ↗ · pubmed ↗
- 7Chang H.L. Chen P.L. Lin S.Y. Chen T.C. Chang K. Ko W.C. Lu P.L. Chen Y.H. Two fatal cases of Aeromonas dhakensis bacteremia and necrotizing fasciitis in severe dengue patients J. Microbiol. Immunol. Infect.20185169269410.1016/j.jmii.2018.03.00329859782 · doi ↗ · pubmed ↗
- 8Hossain S. De Silva B.C.J. Dahanayake P.S. Heo G.J. Characterization of virulence properties and multi-drug resistance profiles in motile Aeromonas spp. isolated from zebrafish (Danio rerio)Lett. Appl. Microbiol.20186759860510.1111/lam.1307530229985 · doi ↗ · pubmed ↗