Genomic Insights into Probiotic Lactococcus lactis T-21, a Wild Plant-Associated Lactic Acid Bacterium, and Its Preliminary Clinical Safety for Human Application
Masanori Fukao, Keisuke Tagawa, Yosuke Sunada, Kazuya Uehara, Takuya Sugimoto, Takeshi Zendo, Jiro Nakayama, Shuichi Segawa

TL;DR
This paper explores the genome and safety of Lactococcus lactis T-21, a wild plant-associated probiotic bacterium, showing it has unique traits and is safe for human use.
Contribution
The study provides the first detailed genomic and clinical safety analysis of the wild Lactococcus lactis T-21 strain.
Findings
Genomic analysis revealed T-21's ability to metabolize diverse plant carbohydrates and produce exopolysaccharides and nisin.
Preliminary clinical trials showed no adverse effects from consuming T-21-fermented milk in healthy adults.
T-21's wild plant origin contributes to its metabolic versatility and distinct probiotic properties compared to dairy strains.
Abstract
Lactococcus lactis T-21 is a lactic acid bacterium isolated from wild cranberries in Japan that demonstrates significant immunomodulatory properties and has been incorporated into commercial health products. However, probiogenomic analyses specific to T-21 have remained largely unexplored. This study performed a thorough genomic characterisation of T-21 and evaluated its safety in initial clinical trials. Genomic analysis revealed substantial genetic diversity and metabolic capabilities, including enhanced fermentative potential demonstrated by its ability to metabolise a wide range of plant-derived carbohydrates, and genetic determinants associated with exopolysaccharide biosynthesis and nisin production, distinguishing T-21 from domesticated dairy strains. These attributes, reflective of its wild plant origin, may contribute to its metabolic versatility and unique probiotic…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
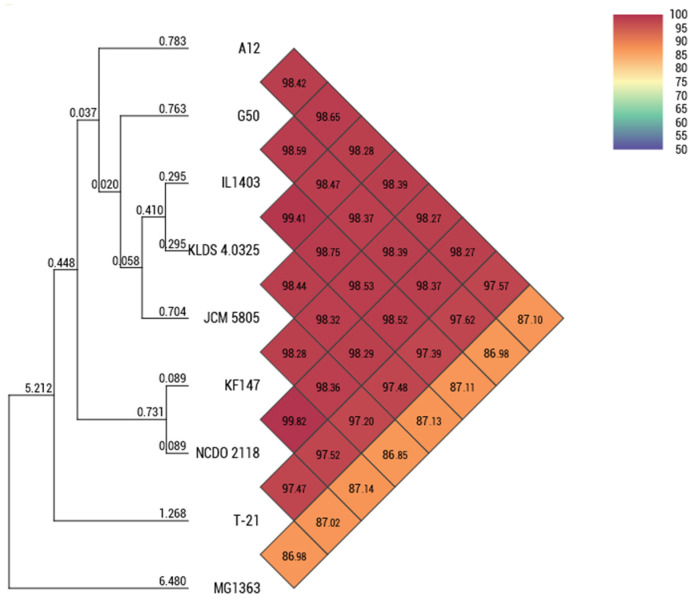 Figure 1
Figure 1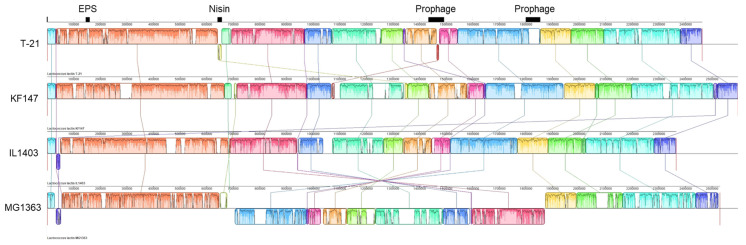 Figure 2
Figure 2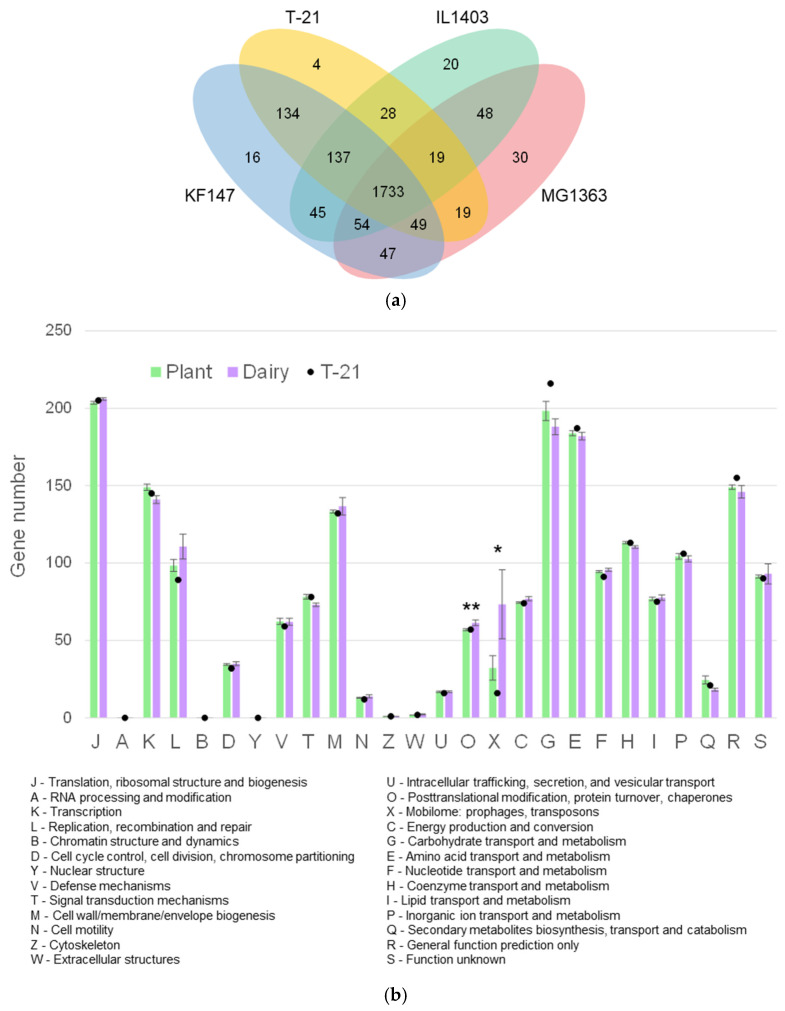 Figure 3
Figure 3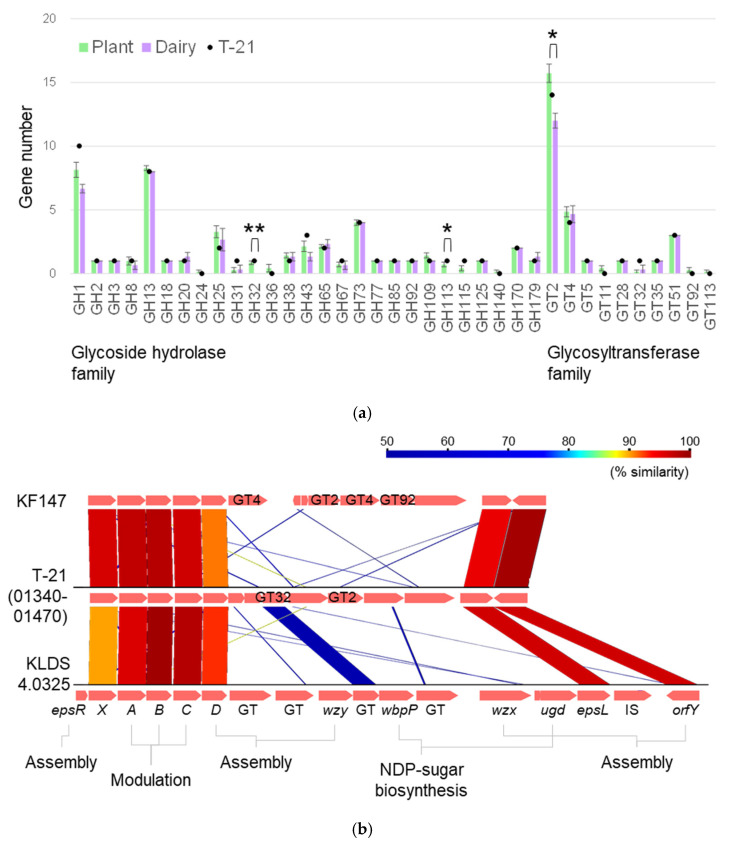 Figure 4
Figure 4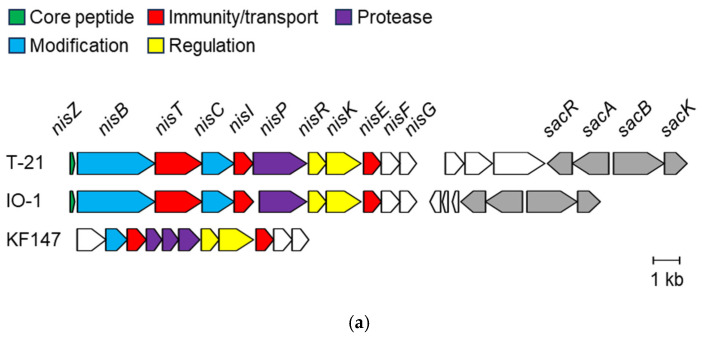 Figure 5
Figure 5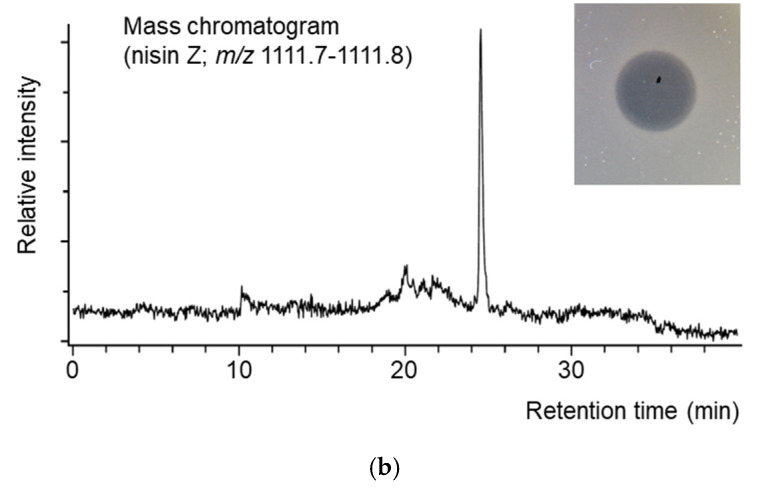 Figure 6
Figure 6| Strain | Origin | Genome | Proteins | Plasmids * | Complete/ | Genbank | Reference |
|---|---|---|---|---|---|---|---|
| subsp. | |||||||
| T-21 | Cranberry | 2.46 | 2344 | ND | Complete | [ | |
| KF147 | Mung bean sprouts | 2.60 | 2534 | 1 | Complete | [ | |
| NCDO 2118 | Frozen peas | 2.55 | 2491 | 1 | Complete | [ | |
| G50 | Napier grass | 2.35 | 2239 | ND | Complete | [ | |
| CAB701 | Cabbage | 2.52 | 2440 | 1 | Complete | [ | |
| A12 | Wheat sourdough | 2.60 | 2677 | 4 | Complete | [ | |
| 14B4 | Almond drupe | 2.58 | 2583 | 1 | Complete | [ | |
| IO-1 | Drain water | 2.42 | 2291 | ND | Complete | [ | |
| JCM 5805 | Dairy starter | 2.53 | 2626 | NA | Draft | [ | |
| IL1403 | Dairy starter | 2.37 | 2404 | ND | Complete | [ | |
| KLDS 4.0325 | Koumiss | 2.59 | 2612 | 6 | Complete | [ | |
| subsp. | |||||||
| MG1363 | Dairy starter | 2.53 | 2647 | ND | Complete | [ |
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsProbiotics and Fermented Foods · Gut microbiota and health · Microbial Metabolites in Food Biotechnology
1. Introduction
Lactococcus lactis is a principal member of the lactic acid bacteria (LAB) group that is internationally recognised for its fundamental role in the production of fermented dairy products [1]. Subspecies lactis and cremoris of L. lactis have been the subject of extensive research owing to their contributions to dairy manufacturing. However, certain strains can inhabit diverse environments beyond milk, including plant and animal materials. Compelling evidence suggests that these organisms originate from plant niches, having undergone a ‘domestication’ process from their ancestral ‘wild’ forms, characterised by reductive evolution and genome decay in dairy-adapted L. lactis strains [2,3,4,5,6,7].
T-21, isolated from wild cranberries in Japan, represents a unique lineage of L. lactis that is more closely aligned with its wild origin than the domesticated dairy strains. This strain is particularly notable for its significant immunomodulatory properties, demonstrating its ability to mitigate allergic reactions by modulating immune responses upon consumption [8]. Since 2022, T-21 has been commercially incorporated into the postbiotic product by Nissin Food Products. Despite its market introduction, comprehensive probiogenomic analyses specific to the wild characteristics of T-21 have remained largely unexplored, with limited insights into its probiotic application.
In this study, we present the detailed genome of L. lactis T-21 and perform a genome-level comparison between wild strains from various sources and long-domesticated dairy strains. Our study elucidated the phenotypic and genotypic diversity among L. lactis strains from distinct environmental niches and explored the potential of non-dairy sources for use in dairy fermentations as probiotics. Additionally, we initiated a preliminary first-in-human prospective, randomised, double-blind, placebo-controlled, parallel-group trial to assess the safety of live T-21 strains in healthy Japanese adults.
2. Materials and Methods
2.1. Genomic Sequencing and Comparative Genomic Analysis
T-21 was isolated from wild cranberries in Shiga Highland, Nagano Prefecture, Japan [8]. This strain was obtained from the microbial repository of Nissin Foods Holdings. T-21 was routinely cultured at 30 °C in MRS broth (Oxoid, Hampshire, UK) without agitation. Genomic DNA was extracted from late-logarithmic phase T-21 cells using standard genomic DNA affinity columns as described previously [9]. Genomic sequencing was conducted using the DNBSEQ-G400 platform (MGI Tech, Shenzhen, China), achieving 200 bp paired-end reads at the Bioengineering Laboratory (Kanagawa, Japan). This yielded a total of 5,152,310 paired-end reads, culminating in a genome coverage of approximately 837×. The sequencing data underwent de novo assembly via the Unicycler v.4.0.7 [10] with scaffolding refined using MUMmer v.3.23 [11] in conjunction with in-house scripts. This enabled alignment with the reference genome of L. lactis CBA3619 (CP042408.1), which was facilitated by Hokkaido System Science (Sapporo, Japan). Genome annotation was conducted using DFAST v.1.6.0 [12] and the RAST toolkit v.1.3.0 [13]. Genomic sequences of the additional L. lactis strains referenced in this study were obtained from GenBank (Table 1). To ensure uniformity, all genomes were re-annotated using the same pipeline, mitigating discrepancies in gene calling and functional annotation owing to the use of various gene-calling software or annotation frameworks.
To ascertain genetic homogeneity across strains, the average nucleotide identity (ANI) was calculated using OrthoANI v.0.93.1 [14]. Phylogenomic relationships were elucidated through pairwise genome comparisons using the genome BLAST distance phylogenetic method [15]. Whole-genome alignments at the nucleotide level, analyses of genomic synteny, and the identification of potential integration sites were conducted using the Mauve alignment tool v.2.4.0 [16], with the resultant visualisations created using GenomeMatcher v.3.0 [17]. Orthologous gene clusters were compared and annotated utilising OrthoVenn3 (January 2024) [18]. Further analysis of the genome-encoded functionalities entailed categorising the protein complement based on the Cluster of Orthologous Groups of Proteins (COG) (January 2024) [19] and the Carbohydrate-Active Enzymes Database (CAZy) (January 2024) [20] assignments. The metabolic pathways were predicted and delineated using the KEGG (Kyoto Encyclopaedia of Genes and Genomes) v.109.0 [21,22] database. PHASTEST (January 2024) [23] was used to detect potential prophages in the genome.
2.2. Phenotypic and Metabolic Characterisation
The capacity for sugar catabolism was evaluated employing the API^®^50 CHL kit (bioMérieux, Marcy l’Etoile, France), which comprises strips infused with 49 distinct carbon sources. To ascertain the production of the bacteriocin by T-21, bacteriocin was isolated from the culture supernatant and subjected to liquid chromatography–mass spectrometry (LC-MS) and bacteriocin activity assays [34]. LC-MS analysis was performed using an Agilent 1100 HPLC system (Agilent Technologies, Santa Clara, CA, USA) coupled with a JMS-T100LC ESI-TOF MS (JEOL, Tokyo, Japan). Chromatographic separation was carried out on an Atlantis T3 reverse-phase column (5 µm, 2.1 mm × 150 mm; Waters, Milford, MA, USA) at 30 °C. The analytical conditions were based on a previously described method [34], with modifications to the elution programme as follows: 20% (v/v) acetonitrile containing 0.05% (v/v) trifluoroacetic acid (TFA) for 10 min, followed by a linear gradient from 20% to 80% (v/v) acetonitrile containing 0.05% (v/v) TFA over 25 min, and a final step with 80% (v/v) acetonitrile containing 0.05% (v/v) TFA for 5 min. To detect and identify the bacteriocin from T-21, the total ion chromatogram was recorded in the range of m/z 500 to 3000. Data acquisition was performed using the JEOL Mass Center program (January 2024). The bacteriocin activity of the culture supernatant was evaluated using the spot-on-lawn method, with Listeria innocua ATCC 33090^T^, a nisin Z-sensitive organism, serving as the indicator strain, as described previously [35]. Following overnight incubation, inhibition zones were measured to assess antimicrobial activity.
2.3. Safety and Clinical Trial Design
The Virulence Factor Database (VFDB) [36] was queried to elucidate the presence of virulence factors and toxin genes within T-21. Furthermore, genetic elements linked to antimicrobial resistance were meticulously examined using the Comprehensive Antibiotic Resistance Database (CARD) v.4.0.0 and the Resistance Gene Identifier (RGI) v.6.0.3 [37] and ResFinder v.4.6.0 [38] tools, employing predetermined criteria of greater than 85% identity and 80% coverage, in agreement with previous studies [9,39,40].
In addition to genomic safety evaluations, a clinical trial was conducted to evaluate the safety of T-21 consumption, facilitated by the AMC Nishi-Umeda Clinic, Osaka, Japan. This trial was approved by the Institutional Review Board of the AMC Nishi-Umeda Clinic and adhered to the principles of the Declaration of Helsinki and the Ethical Guidelines for Medical and Health Research (UMIN ID: UMIN000049824). The study included male and female participants aged 20–65 years. Healthy male and female volunteers aged 20–65 years were recruited for the study. Eligibility criteria included the absence of chronic illnesses, no regular use of medications affecting gut health, and no history of severe allergies to food or beverages. Applicants who met these criteria were enrolled as participants. Of the applicants, 64 met the eligibility criteria and were subsequently randomised into two groups: a T-21 group (n = 32), receiving a daily dose of ≥1.0 × 10^11^ colony-forming units (cfu), and a placebo group (n = 32) over an eight-week period (Table 2). To ensure the consistency and quality of the test beverage, it was freshly prepared each week during the trial. The viable count of T-21 was guaranteed at ≥1.0 × 10^11^ cfu per dose by plating appropriate dilutions of the beverage on BCP agar (Eiken, Tokyo, Japan) and incubating at 30 °C for 48 h under aerobic conditions. The test beverage was stored at 0 to 10 °C until consumed, as described in the footnotes of Table 2. The participants were asked to document their health status, medication intake, and the frequency of consumption of the test beverages. Clinical evaluations were scheduled at weeks 4 and 8 of the intake duration. The precision and reliability of the trial were ensured by the involvement of M&I Science, Osaka, Japan, as a contracted research organisation.
3. Results and Discussion
3.1. Genomic Architecture and Strain Diversity
T-21 contained a 2.46 Mb circular chromosome without plasmids (Table 1). The ANI values between T-21 and other strains of L. lactis subsp. lactis showed similarity percentages within 97.2–97.6% (Figure 1). The ANI leverages whole-genome sequences and serves as a critical instrument for species delineation, adhering to the 95–96% threshold for species definition [41]. This confirmed T-21’s classification within L. lactis subsp. lactis. The generated phylogenetic supertree illustrated a bifurcation indicative of niche-specific adaptation (Figure 1), with isolates of dairy and plant origin forming distinct clusters, supporting the identification of T-21 as an environmentally friendly wild strain, which suggests greater genetic diversity than its domesticated or dairy counterparts. This study provides an in-depth examination of the distinct genomic and phenotypic characteristics of T-21 by comparing its genome with those of prototypical strains: IL1403 [31], a model for domesticated strains; KF147 [4], representative of wild strains; and subsp. cremoris MG1363 [33]. Comparative analysis of orthologous gene clusters indicated that T-21 comprised 2123 gene clusters, of which 1733 were shared with the four reference strains and 202 singletons, demonstrating a significant degree of conservation within subsp. lactis with strain-specific genomic regions (Figure 2 and Figure 3a). Genomic insertions in T-21 (especially those associated with prophage, exopolysaccharide (EPS) biosynthesis, and nisin production) are shown in Figure 2.
3.2. Genomic Insights into Metabolic Capabilities and Phenotypic Divergence
3.2.1. Functional Genomic Overview
Furthermore, genes unique to T-21 (particularly those enabling the metabolism of plant-derived carbohydrates) underscore the distinctive capabilities of the strain. This extensive genomic comparison highlights the diversity and evolutionary dynamics within L. lactis subsp. lactis, shedding light on the adaptive strategies and functional capacities of environmental wild strains, such as T-21. Moreover, in assessing their metabolic capabilities, COG groupings were compared. These groupings are crucial to understanding niche adaptation and provide insights into the metabolic versatility of a strain. In T-21, a significant proportion of genes fall under ‘metabolism’, with ‘carbohydrate transport and metabolism (G)’ and ‘amino acid transport and metabolism (E)’ being the dominant categories (403 genes) (Figure 3b). The pronounced abundance of ‘carbohydrate transport and metabolism (G)’ in T-21 suggested an adaptive response to diverse nutritional sources, a trait presumably inherited from plants. Additionally, the ‘Mobilome: prophages, transposons (X)’ category shows notable variability between plant- and dairy-derived strains, reflecting differences in their genomic architecture. Unlike their dairy counterparts, plant-derived strains do not exhibit gene reduction, potentially indicating the genomic instability of plant-associated genomes [7]. In this aspect, T-21 has a lower prevalence.
3.2.2. Genetic Determinants of EPS Biosynthesis
In the realm of polysaccharide production among LAB, L. lactis boasts two chromosomal loci dedicated to the synthesis of cell wall polysaccharides, known as the rhamnose-glucose polysaccharide (RGP) and EPS clusters, alongside a gene cluster for teichoic acid biosynthesis [42]. The notable variability in gene order and composition within these regions among L. lactis strains suggests that these organisms produce a broad spectrum of EPS structures. This genetic diversity implied that T-21 also possessed the genetic machinery required for EPS biosynthesis. Our genomic analysis of T-21 focused on carbohydrate-active enzymes, sugar transport systems, and nucleotide sugar biosynthesis pathways, revealing a predominance of genes related to the glycoside hydrolase (GH) and glycosyltransferase (GT) families in the CAZy (Figure 4a). Specifically, GH1 and GH13 were the most represented GH families, suggesting adaptation to a carbohydrate-rich environment. The GH1 family primarily encodes enzymes, such as beta-glucosidase, beta-galactosidase, and lactase, whereas GH13 includes pullulanase, maltotriose-forming alpha-amylase, and amylosucrase, which play pivotal roles in biological processes, including sugar biosynthesis and cellular metabolism [43]. Glucosidases encoded by the GH1 and GH13 families are instrumental in hydrolysing glucosidic bonds to produce monosaccharides, which serve as precursors for EPS biosynthesis [44]. This enzymatic activity underscores T-21’s ability to efficiently utilise a variety of carbon sources, facilitating the synthesis of nucleotide sugar bases essential for EPS production.
Furthermore, our annotation identified seven classes of GT family genes in T-21 (Figure 4a), indicating a high capacity for sugar biosynthesis. GT2 and GT4 are the predominant families, and these genes are mostly associated with EPS biosynthesis gene clusters in LAB [45,46]. The diversity of GT within T-21, especially those encoded within the EPS gene cluster, suggests a significant polymorphism in GT that is intricately linked to the molecular diversity of EPS [7,42,46]. The EPS biosynthesis cluster in T-21 comprised a series of genes essential for EPS synthesis and export (Figure 4b). Among these, epsA and epsB are crucial for determining EPS length and are indispensable for its biosynthesis. The epsC gene is not essential for biosynthesis but plays a regulatory role, whereas epsD acts as a priming glycotransferase that is crucial for EPS production [47]. This comprehensive gene cluster underscores T-21’s sophisticated EPS biosynthesis capabilities. Comparative analyses of eps gene clusters across various LAB strains have highlighted significant variability, which likely reflects the diverse environmental origins of these strains [7,48]. LAB strains with genes associated with EPS production exhibit a pronounced capacity for biofilm formation and adherence to mucosal interfaces. Furthermore, it contributes to immunomodulatory functions [49,50,51,52]. Importantly, the EPS biosynthesis potential of T-21 aligns with previous findings that postbiotics derived from T-21 exhibit immunomodulatory properties [8]. However, the specific structures and functionalities of the EPS produced by T-21 remain unclear and warrant further investigation. Such studies could provide insights into the adaptive mechanisms and therapeutic applications of this strain in biotechnology and health sciences.
3.2.3. Enhanced Fermentative Potential and Phenotypic Adaptation
T-21 was characterised by an abundance of GH1 and GH13 in terms of fermentative capabilities (Figure 4a). The genomic composition of T-21 encompassed genetic sequences indicative of a broader variance in carbohydrate metabolism compared to the domesticated dairy strain IL1403. The presence of several additional T-21-specific GHs suggests the potential for novel activities that are unique to T-21. GH catalyses the hydrolysis of glycosidic bonds between two or more carbohydrates or between a carbohydrate and a non-carbohydrate moiety [20]. The functionality of these genes was assessed through phenotypic analysis involving the testing of 49 carbon sources with varying complexities. Growth assays on a diverse array of mono- and oligosaccharides demonstrated that plant-derived isolates (including T-21) grew on a more extensive range of sugar substrates than the strains IL1403 or JCM 5805 (Table 3). The utilisation of carbon sources presents the most significant phenotypic divergence between strains, with plant isolates uniquely metabolising L-arabinose, D-xylose, mannitol, sucrose, and gluconate—carbohydrates predominantly derived from plant sources. A comparative analysis of the genetic and phenotypic profiles of these strains revealed specific traits of T-21 that are potentially related to their survival in plant environments. The extensive repertoire of strain-specific genes underscores their considerable adaptive potential within complex ecosystems. The observed differences are consistent with the hypothesis that dairy isolates of L. lactis evolved from plant isolates, as evidenced by genome reduction in dairy strains [2,3,4,5,6,7]. This is particularly highlighted by the diminished complement of chromosomal genes encoding GH, which is involved in carbohydrate metabolism and many of which are implicated in plant polysaccharide metabolism. For instance, a notable aspect of this genomic region is the presence of genes involved in arabinose metabolism, including GH43 alpha-N-arabinofuranosidase [27,48]. The organisation of the ara cluster in T-21 was consistent with that observed in strain KF147. Several phenotypic distinctions were correlated with genetic variances between the strains, thereby elucidating the underlying genetic basis for the observed phenotypes.
3.2.4. Nisin Production and Antimicrobial Activity
Another notable characteristic of T-21 is that the complete nisin gene cluster nisZBTCIPRKFEG [53] is present on the chromosome of T-21 and is located in the nisin-sucrose transposon, similar to that of IO-1 [29] (Figure 5a). These genes are essential for the biosynthesis, regulation, and immunity of nisin Z. KF147 possesses an incomplete chromosomal nisin gene cluster, rendering it incapable of nisin production, although there are certain immunity genes, specifically nisFEG and/or nisI [48]. Notably, several strains derived from plant or vegetable sources were identified as producers of nisin Z, which are distinct from plant-origin strains [7]. It was hypothesised that nisin Z production in L. lactis functions in microbe–plant interactions (defence or adhesion) or might confer protection against other species encountered during plant fermentation as well as on living plant surfaces. We identified nisin Z production by T-21 using the LC-MS system previously developed for the rapid detection and identification of bacteriocins. Consistent with a previous report [34], nisin Z was detected as [M+3H]^3+^ ions in the partially purified culture supernatant of T-21. The molecular weight of the compound was 3331 Da, which corresponded to that of nisin Z (Figure 5b). Furthermore, antimicrobial activity resembling that of nisin was verified in the culture supernatant. In particular, a nisin-like clear-edged inhibitory zone was observed, confirming the presence of nisin-like antimicrobial activity. The use of L. innocua as an indicator organism was based on previous studies demonstrating its sensitivity to nisin [35]. The verification of antimicrobial activity against other representatives of pathogenic enteroflora, which are more commonly associated with gastrointestinal diseases, remains an important area for future research.
3.3. Prophages: Genomic Features and Considerations
Prophages constitute a significant element since resistance to phages is a principal factor in the selection of strains for industrial culture applications. Additionally, careful attention should be given to the potential for starter culture prophages to mutate and become virulent [54]. We identified two putative prophages in the chromosome of T-21 (Figure 2). Protein analysis indicated that most proteins within these loci exhibited high amino acid identity with known prophages in the dairy starter strain IL1403, categorising them within the same family. Regarding the prophages of IL1403, no reports of virulent conversion during fermentation have been documented, and they are not considered a risk factor in dairy production. Furthermore, it has been reported that IL1403 prophages significantly contribute to diverse aspects of L. lactis cell physiology, many of which are relevant to its industrial and health applications [55]. The presence of these prophages appears to be a natural characteristic of T-21, and their behaviour presumably aligns with that of IL1403. Under certain cultivation conditions, there is the potential for these to be activated as temperate phages and transition into the lytic cycle; however, the risk of prophage activation can be minimised with appropriate starter culture management and controlled cultivation processes. Consequently, the prophages in T-21 do not inherently present technological challenges in terms of standard manufacturing operations.
3.4. Safety Assessment and Clinical Trial Outcomes
Safety assessment of new probiotic candidates requires further evaluation since previous studies have reported infections owing to the consumption of probiotics [56]. Antibiotics can persist for extended periods in soil and water-based wild environments; thus, the widespread use of agricultural antibiotics may select for resistant strains of bacteria in these habitats. Therefore, if non-dairy or wild isolates (such as T-21) are to be used as cultures in food processing, their antibiotic resistance traits must be carefully assessed and should fall within the guidelines set out by the EFSA [57,58]. We found no genes related to transferable antibiotic resistance and virulence factors after conducting a thorough survey of the T-21 genome. Specifically, no genes homologous to virulence factors commonly associated with clinically relevant pathogens, resistance genes, or mutations conferring antibiotic resistance were detected. Furthermore, T-21 lacked plasmids associated with its mobilisation capabilities.
In addition to the genome-based safety assessment, a randomised, double-blind, placebo-controlled, parallel-group trial demonstrated the safety of T-21, which included consuming ≥1.0 × 10^11^ cfu daily for eight weeks. All participants who completed the trial were included in the safety analysis. During the trial, adverse events were reported in both the T-21 group (14 events in 8 participants) and the placebo group (10 events in 7 participants), as detailed in Table 4. All reported symptoms were mild and included common issues such as headache, abdominal pain, and nasal congestion. None of the events were deemed serious or associated with the consumption of the test beverages. No abnormal changes were observed in general blood biochemical parameters in either group after eight weeks of daily consumption. Importantly, none of the observed changes were considered clinically relevant. These findings suggest that both test beverages are safe for consumption. While the findings from this study confirm the genomic safety of T-21 and its safe consumption in humans over an eight-week period, there are limitations to consider. The sample size of this clinical trial (n = 64) may not provide sufficient statistical power to detect rare adverse effects or subtle health benefits associated with T-21 consumption. Furthermore, the trial did not evaluate the potential efficacy of T-21 against specific gastrointestinal pathogens or its broader probiotic benefits beyond safety. Future studies should involve larger, more diverse populations and longer trial durations to assess the potential health benefits of T-21 consumption. Additionally, investigations into T-21’s effects on gut microbiota composition and functionality, as well as its potential to alleviate gastrointestinal disorders, are important directions for future research.
4. Conclusions
In conclusion, genomic insights into Lactococcus lactis T-21 revealed a strain with a distinct genetic profile, characterised by its capacity for EPS biosynthesis and nisin production, significantly diverging from wild and domesticated dairy strains. The origin of this strain from a plant-associated niche contributes to its unique metabolic versatility, including the ability to metabolise a wide range of plant-derived carbohydrates. These capabilities reflect its functional diversity and underline its potential for probiotic applications. This preliminary first-in-human clinical trial underscores live T-21’s safety for human consumption, marking a significant step towards harnessing its immunomodulatory properties for probiotic applications. Future investigations will delve deeper into T-21’s functional mechanisms, exploring its full therapeutic potential, and will employ metabolomic and proteomic approaches to further characterise its unique properties. This study enriches our understanding of the genetic diversity among L. lactis strains and lays the groundwork for leveraging wild plant-associated strains in the development of innovative probiotic formulations.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Smid E.J. Kleerebezem M. Production of aroma compounds in lactic fermentations Annu. Rev. Food Sci. Technol.2014531332610.1146/annurev-food-030713-09233924580073 · doi ↗ · pubmed ↗
- 2van Hylckama Vlieg J.E.T. Rademaker J.L.W. Bachmann H. Molenaar D. Kelly W.J. Siezen R.J. Natural diversity and adaptive responses of Lactococcus lactis Curr. Opin. Biotechnol.20061718319010.1016/j.copbio.2006.02.00716517150 · doi ↗ · pubmed ↗
- 3Kelly W.J. Ward L.J.H. Leahy S.C. Chromosomal diversity in Lactococcus lactis and the origin of dairy starter cultures Genome Biol. Evol.2010272974410.1093/gbe/evq 05620847124 PMC 2962554 · doi ↗ · pubmed ↗
- 4Siezen R.J. Bayjanov J. Renckens B. Wels M. van Hijum S.A.F.T. Molenaar D. van Hylckama Vlieg J.E.T. Complete genome sequence of Lactococcus lactis subsp. lactis KF 147, a plant-associated lactic acid bacterium J. Bacteriol.20101922649265010.1128/JB.00276-1020348266 PMC 2863553 · doi ↗ · pubmed ↗
- 5Bachmann H. Starrenburg M.J.C. Molenaar D. Kleerebezem M. van Hylckama Vlieg J.E.T. Microbial domestication signatures of Lactococcus lactis can be reproduced by experimental evolution Genome Res.20122211512410.1101/gr.121285.11122080491 PMC 3246198 · doi ↗ · pubmed ↗
- 6Kelleher P. Bottacini F. Mahony J. Kilcawley K.N. van Sinderen D. Comparative and functional genomics of the Lactococcus lactis taxon; insights into evolution and niche adaptation BMC Genom.20171826710.1186/s 12864-017-3650-528356072 PMC 5372332 · doi ↗ · pubmed ↗
- 7Wels M. Siezen R. van Hijum S. Kelly W.J. Bachmann H. Comparative genome analysis of Lactococcus lactis indicates niche adaptation and resolves genotype/phenotype disparity Front. Microbiol.201910410.3389/fmicb.2019.0000430766512 PMC 6365430 · doi ↗ · pubmed ↗
- 8Uehara K. Sunada Y. Kondo S. Matsuo S. Effects of food containing Lactococcus lactis strain T 21 on the improvement of skin condition: A randomized, double-blind, placebo-controlled, parallel-group study Biosci. Microbiota Food Health 20244338139010.12938/bmfh.2023-08639364129 PMC 11444856 · doi ↗ · pubmed ↗