Toxoplasma gondii Type I TR and ROP16 Synergistically Downregulate IL-12 to Inhibit Host Reactive Oxygen Species Production
Xiaoling Geng, Ruifang Li, Jingying Du, Manyu Zhang, Wei Jiang, Qing Sun, Rongsheng Mi, Shuang Qin, Quan Wang

TL;DR
This study shows that two Toxoplasma gondii proteins, TR and ROP16, work together to reduce inflammation and protect the parasite from immune damage.
Contribution
The study reveals a synergistic mechanism between TR and ROP16 in T. gondii to inhibit host ROS production and immune response.
Findings
Deleting both TR and ROP16 genes reduced T. gondii's growth and virulence.
The double deletion increased ROS levels and lipid oxidation in the parasite and host cells.
TR and ROP16 synergistically suppressed IL-12, reducing host ROS production.
Abstract
Toxoplasma gondii is an obligate intracellular opportunistic protozoan parasite. T. gondii invasion disturbs the balance between reactive oxygen species (ROS) production and antioxidant capacity in the host, triggering the oxidative stress response. Thioredoxin reductase (TR) of T. gondii helps to escape ROS-induced damage in the host, whereas T. gondii rhoptry protein 16 (ROP16) downregulates host innate immunity to suppress excessive inflammation and inhibit ROS production. However, whether TR and ROP16 synergistically promote resistance to ROS-induced damage remains unclear. Here, we used the CRISPR/Cas9 technology to successfully obtain a double TR and ROP16 gene knockout T. gondii strain. The double deletion of TR and ROP16 in T. gondii weakened its growth ability in vitro and decreased its virulence in vivo. Moreover, the double deletion of TR and ROP16 resulted in a lower…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
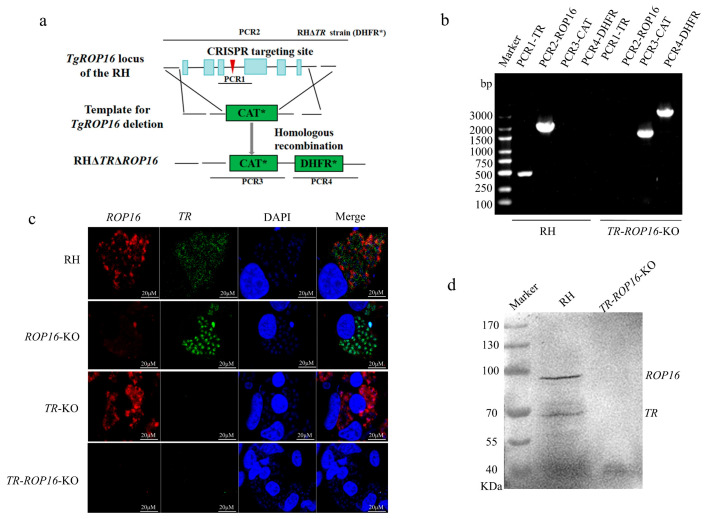 Figure 1
Figure 1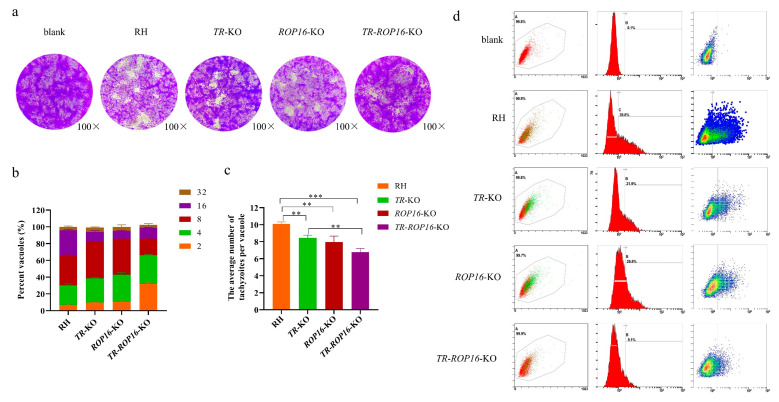 Figure 2
Figure 2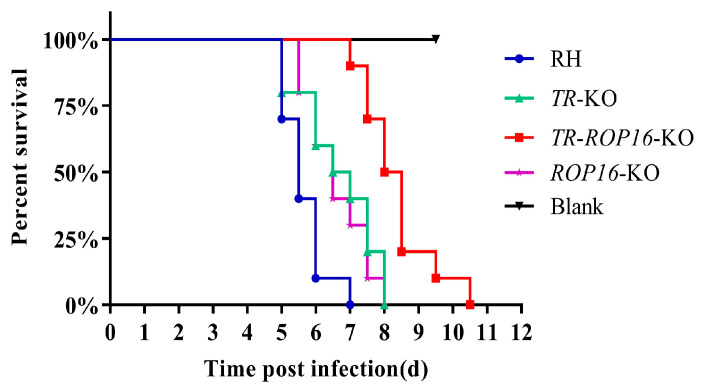 Figure 3
Figure 3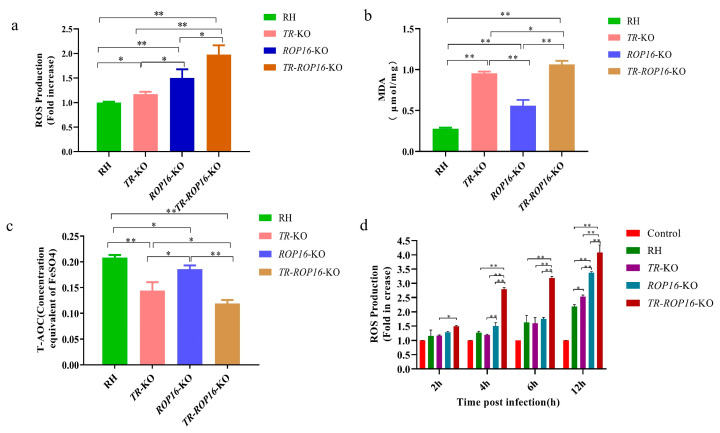 Figure 4
Figure 4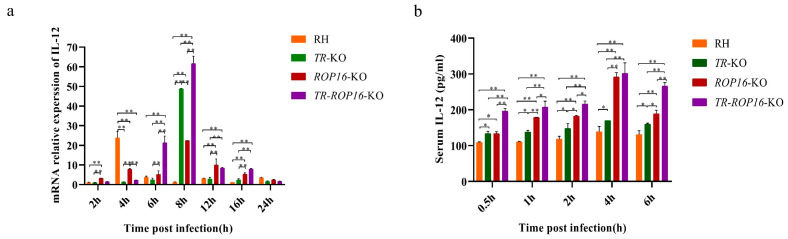 Figure 5
Figure 5- —Natural Science Foundation of Shanghai
- —Shanghai Science and Technology Promotion Agriculture Innovation Program
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsToxoplasma gondii Research Studies · Heme Oxygenase-1 and Carbon Monoxide · Mosquito-borne diseases and control
1. Introduction
Toxoplasma gondii, an intracellular parasite of the phylum Apicomplexa, class Conoidasida, and order Eucoccidiorida, is a feline intestinal coccidium species. T. gondii shows obligate parasitism in the nucleated cells of the host and causes toxoplasmosis, a common parasitic disease in nearly one-third of the world’s population [1,2]. When a host is infected with T. gondii, it develops systemic clinical symptoms, especially when its immune system is weakened [3]. Typical clinical manifestations of toxoplasmosis include toxoplasmic encephalitis, anterior/posterior uveitis, and neurological, ocular, or systemic damage in neonates [4]. Infection of animals, e.g., sheep and goats, with T. gondii may result in fetal death, mummification, abortion, stillbirths and neonatal deaths, which cause serious economic losses [5]. Therefore, T. gondii causes serious harm to human health and the livestock industry [6].
Innate immunity is the first line of host defense against T. gondii infection. The immune effector cells release reactive oxygen species (ROS) following the activation of the IL-12-IFNγ-STAT1 pathway to eliminate T. gondii infection [7,8,9]. The immune microenvironment of the host is toxic to T. gondii [10]. The virulence factors of T. gondii help to counter ROS-induced damage in different ways [11]. On the one hand, T. gondii has developed an antioxidant defense system, comprising superoxide dismutase, catalase, glutathione peroxidase, and peroxiredoxins, which protect T. gondii cells against ROS [12]. T. gondii also uses the thiol-reduction system, including thioredoxin reductase (TR), glutathione, glutaredoxin, and a specific reductase to resist oxidative damage [12,13]. TR is a NADPH-dependent FAD domain-containing dimeric selenium enzyme belonging to the family of pyridine nucleotide–disulfide oxidoreductases, which includes TrxR1, TrxR2, and TrxR3. The main function of these proteins is to resist oxidative damage brought about by the immune system of the host and maintain the reduced form of thioredoxin (Trx) by using NADPH [14,15,16,17]. On the other hand, T. gondii inhibits the production of ROS in host immune cells by downregulating the IL-12-IFN-γ-STAT signaling pathway. STAT3/STAT6 are activated and phosphorylated by rhoptry protein 16 (ROP16) secreted by type I T. gondii, which ultimately downregulates the activity of the IL-12-IFNγ-STAT1 pathway, thereby reducing the production of ROS and resisting the oxidative stress damage inflicted by the host [18,19,20,21]. However, whether ROP16 and TR secreted by T. gondii have a synergistic effect, promoting resistance to oxidative damage, is currently unknown.
To date, no protein has been found to have a synergistic effect with TR in resisting ROS damage from host immune cells. In the present study, we used CRISPR/Cas9 technology to knock out ROP16 in the TR knockout strain of T. gondii to obtain the TR-ROP16-KO strain. To investigate the functional contributions of TR and ROP16, we detected and analyzed the factors related to the ROS-scavenging pathway of T. gondii.
2. Materials and Methods
2.1. T. gondii and Cell Cultures
Tachyzoites of the T. gondii strain RH, TR gene deletion strain (TR-KO), and ROP16 gene deletion strain (ROP16-KO) were maintained in vitro by passaging in African green monkey kidney Vero cells or the mouse macrophage cell line RAW264.7. Vero and RAW264.7 cells were grown in T75 cell culture flasks containing Dulbecco’s Modified Eagle Medium (DMEM) supplemented with 10% fetal bovine serum, 2 mM glutamine, 100 kU/L streptomycin, and 400 kU/L penicillin, at 37 °C in the atmosphere of 95% air and 5% CO_2_. The infected Vero cells were lysed through a 5-gauge needle, and the tachyzoites were filtered using a 5 µm pore size Millipore filter (Merck-Millipore, Darmstadt, Germany). The purified parasites were counted using a cell counting chamber under a microscope (Olympus CKX53, Tokyo, Japan).
2.2. Construction of Transgenic Parasite Strains
All plasmids, gRNAs, and primers used in this study are listed in Supplementary Table S1. A double gene knockout strain (TR-ROP16-KO) was constructed using the CRISPR/Cas9-mediated gene targeting technology. In this study, to ensure ROP16 gene knockout, a CRISPR/Cas9 vector expressing dual-guide RNA was constructed. First, full-length sequences of the ROP16 gene were downloaded from the NCBI website (GenBank: GQ249080.1), and gRNA1 and gRNA2 were designed using an optimized CRISPR design algorithm (http://chopchop.cbu.uib.no). Next, guide RNA targeting the ROP16 gene locus was used to replace the single-guide RNA site in the pSAG1::CAS9-U6:sgUPRT plasmid (#54467, Addgene, https://www.addgene.org/) using a ClonExpress MultiS one-step cloning kit (Vazyme Biotech, Nanjing, China) to construct the plasmids pSAG1:CAS9-U6:sgROP16-1 and pSAG1:CAS9-U6:sgROP16-2. Finally, the pSAG1::CAS9-U6:sgROP16 plasmid containing dual gRNAs was obtained by ligating the pSAG1::CAS9-U6::sgROP16-1 plasmid and gRNA2 fragment digested with the KpnI and XhoI enzymes (Takara, Kyoto, Japan) using a T4 DNA Ligation Kit Ver.2.1 (Takara, Kyoto, Japan). To construct the TR-ROP16-KO strain, the pROP16::CAT-D plasmid was constructed by inserting the 3′ and 5′ regions flanking the ROP16 gene amplified from the genomic DNA of the RH strain into both sides of the CAT resistance fragment of the pCas9-CAT plasmid. Chloramphenicol (CAT) formed the CAT* cassette for positive screening, which was cloned from the pCas9-CAT plasmid (#80323, Addgene, https://www.addgene.org/). The pSAG1:CAS9::TgU6:sgROP16 and pROP16::CAT-D plasmids (Supplementary Figure S1) were transferred into the tachyzoites of the TR-KO strain by electroporation, and homologous recombination in the parasites was confirmed by PCR.
2.3. Western Blot Analysis
Freshly purified tachyzoites (1 × 10^8^) were lysed using 200 μL of protein lysate (a mixture of the radioimmunoprecipitation assay buffer and phenylmethylsulfonyl fluoride at a 100:1 ratio) on ice for 3–5 min, and the supernatant of the sample was obtained by centrifugation. Equivalent amounts of the sample supernatant were electrophoresed on 10% SDS polyacrylamide gels and then electroblotted onto 0.45 µm nitrocellulose membranes. Mouse anti-ROP16 (1:1000) and rabbit anti-TR (1:2000), which were prepared and preserved in our laboratory previously, were used as primary antibodies. Horseradish peroxidase-labeled goat anti-mouse IgG and goat anti-rabbit IgG (1:4000) were used as secondary antibodies (Bersee, Beijing, China). Membranes were visualized using a standard enhanced chemiluminescence system (Bio-Rad, Hercules, CA, USA).
2.4. Immunofluorescence Assay
Freshly purified tachyzoites (1 × 10^5^) were used to infect Vero cells seeded on coverslips for 4 h. Vero cells were washed with phosphate-buffered saline (PBS) to remove parasites that failed to invade the cells. Subsequently, the samples were further cultured for 20 h at 37 °C in an atmosphere of 95% air and 5% CO_2_. Cell samples were fixed with 4% paraformaldehyde and permeabilized with 0.5% Triton X-100 (Licheng Biology, Nanjing, China). Subsequently, the samples were incubated with anti-ROP16 (1:1000) and rabbit anti-TR (1:2000) primary antibodies for 1 h at 37 °C, and then Alexa Fluor 488- and Alexa Fluor 594-labeled IgG secondary antibodies (Merck, Rahwa, NJ, USA) for 1 h at 37 °C. The nucleus was visualized by incubation with DAPI for 15 min at room temperature. The cells were observed under a fluorescence microscope (LSM 900, Zeiss, Oberkochen, Germany).
2.5. Invasion and Proliferation Assays
For the invasion assays, freshly purified tachyzoites (1 × 10^6^) were labeled using a CDFA-SE cell proliferation kit (Beyotime, Shanghai, China). The labeled tachyzoites were inoculated into each well of 6-well plates containing Vero cells at 90% confluency and cultured for 12 h at 37 °C in an atmosphere of 95% air and 5% CO_2_. The wells were washed with blank DMEM to remove parasites that did not invade Vero cells. Then, the cells were completely digested by trypsin, resuspended in 500 μL of DMEM supplemented with 10% fetal bovine serum, and analyzed with a Cytomics FC 500 flow cytometer (Beckman Coulter, Brea, CA, USA). The percentage of cells infected with T. gondii was calculated using FlowJo software version 7.6.1 (Tree Star Inc., Ashland, OR, USA).
For proliferation assays, freshly purified tachyzoites (1 × 10^5^) were inoculated into each well of 6-well plates containing Vero cells at 80% confluency and cultured for 4 h at 37 °C. The wells were washed with PBS to remove parasites that failed to invade Vero cells. Then, the cells were cultured at 37 °C in an atmosphere of 95% air and 5% CO_2_ for 24 h. One hundred vacuoles of T. gondii were randomly counted, and the number of tachyzoites in each vacuole was recorded. The results were expressed as the percentage of vacuoles containing different numbers of tachyzoites.
2.6. Plaque Assay
A monolayer of Vero cells grown in 6-well plates was infected with 200 freshly purified tachyzoites per well for 6 days. The monolayers were then fixed with 4% paraformaldehyde (Labgic Technology Co., Ltd., Beijing, China) at 4 °C for 20 min, stained with crystal violet for 15 min, and gently washed thrice with PBS. Plaques formed by the growing parasites were counted using an inverted microscope (Olympus CKX53, Tokyo, Japan). All strains were tested thrice, each with three technical replicates.
2.7. Virulence Assay
Freshly purified tachyzoites (1 × 10^3^) were inoculated into the abdominal cavity of 7-week-old female KunMing mice (JieSiJie, Shanghai, China), and animals injected with the same volume of sterile saline were used as the blank group. The time to death of the mice inoculated with different strains of T. gondii was monitored. The experimental results were visualized by plotting the survival curves using Prism 8.0 (GraphPad Software Inc., La Jolla, CA, USA).
2.8. Determination of Malondialdehyde (MDA), Total Antioxidant Capacity (T-AOC), and Reactive Oxygen Species (ROS) in Tachyzoites
MDA, T-AOC, and ROS levels were determined in freshly purified tachyzoites of T. gondii RH, TR-KO, ROP16-KO, and TR-ROP16-KO strains. To measure MDA levels, tachyzoites (1 × 10^8^) were lysed with 200 μL of IP (Beyotime, Shanghai, China) cell lysate, and after the supernatant was collected, MDA levels were determined at 535 nm with an MDA assay kit (Beyotime, Shanghai, China) and expressed in MDA amount per milligram of protein (μmol/mg). To measure T-AOC levels, tachyzoites (1 × 10^8^) were resuspended in 2 mL of PBS. The mixed solution was sonicated on ice, the supernatant was collected, and T-AOC levels were measured at 593 nm using a T-AOC assay kit (Beyotime, Shanghai, China). The total antioxidant capacity results were represented as the concentration of the FeSO4 standard solution (mM). To determine the ROS levels, tachyzoites (1 × 10^7^) were labeled with 2′,7′-dichlorofluorescein diacetate (Beyotime, Shanghai, China) at 37 °C for 20 min and resuspended in 200 μL of DMEM. The resuspended solution was added to a 96-well plate, and the intracellular fluorescence intensity was determined using a BioTek Synergy 2 luminescence microplate reader (BioTek, Winooski, VT, USA). Intracellular ROS levels were expressed as multiples of those in the RH strain.
2.9. Determination of ROS Levels in Macrophages
Freshly purified tachyzoites (1 × 10^6^) were inoculated into a 6-well plate containing RAW 264.7 cells at 80% confluency and cultured for 2, 4, 6, or 12 h at 37 °C in an atmosphere of 95% air and 5% CO_2_. The wells were washed with DMEM to remove T. gondii that failed to invade RAW 264.7 cells. Then, the cells were digested by trypsin, labeled with 2′,7′-dichlorofluorescein diacetate (Beyotime, Shanghai, China) at 37 °C for 20 min, and washed thrice with DMEM, and the ROS levels were measured using a BioTek Synergy 2 luminescence fluorescence microplate reader (BioTek, Winooski, VT, USA). The results were expressed as multiples of the fluorescence intensity relative to that of the blank control group.
2.10. Quantitative Reverse Transcriptase PCR
Total RNA was extracted from macrophages infected with tachyzoites of T. gondii RH, TR-KO, ROP16-KO, and TR-ROP16-KO strains, and cDNA was obtained using a reverse transcription kit (Takara, Kyoto, Japan). Quantitative detection of IL-12 mRNA in macrophages was performed with a SYBR-Green Premix Ex Taq kit (Takara, Kyoto, Japan). The expression level of the β-actin gene was used as a reference. The primers used in RT-qPCR are listed in Supplementary Table S2; relative fluorescence quantitative PCR (Applied Biosystems 7500 Real-Time PCR System, Thermo Fisher, Waltham, MA, USA) was used to detect the mRNA levels of IL-12. Three biological replicates were used in this study. The mRNA transcription level of each cytokine gene was analyzed using the 2^−ΔΔCt^ method.
2.11. Determination of Mouse Serum IL-12 Levels
Tachyzoites (5 × 10^6^) of T. gondii RH, TR-KO, ROP16-KO, and TR-ROP16-KO strains were inoculated into the abdominal cavity of 6-week-old female Kunming mice (JieSiJie, Shanghai, China). Blood was collected at 0.5, 2, 4, and 6 h after the infection with tachyzoites via tail-vein bleed. Samples were centrifuged at 2000 rpm for 8 min, and serum was collected and stored at −80 °C. The serum level of IL-12 was determined by an ELISA kit (Biolegend, San Diego, CA, USA). Three biological and three technical replicates were used in this experiment.
2.12. Statistical Analysis
SPSS (version 17.0; SPSS, Inc., Chicago, IL, USA) was used for all analyses. Data were analyzed using one-way analysis of variance to determine the statistical significance of strain effects on various parameters. The results were expressed as the mean ± standard deviation. Effects were considered as significant if p < 0.05 and extremely significant if p < 0.01.
3. Results
3.1. Construction of the TR and ROP16 Double Gene Knockout Strain Using the CRISPR/Cas9 Method
To investigate the biological roles of TR and ROP16 in the T. gondii type I strain RH, we used the CRISPR/CAS9 technique to disrupt the ROP16 gene in the TR-KO strain by inserting the chloramphenicol resistance fragment (CAT*-Ts) in the sgRNA-targeted coding region, enabling pROP16-CAT-D to be used for the deletion of the entire ROP16-coding sequence (Figure 1a). Single-clone strains were identified using PCR, and the results showed that the CAT coding sequence was successfully inserted into the target position (Figure 1b). To further confirm that the TR and ROP16 genes of T. gondii were both deleted, immunofluorescence and Western blot analyses were used to show the absence of TR and ROP16 protein expression in the TR-ROP16-KO strain (Figure 1c,d). These results were consistent with the PCR results, proving that the TR-ROP16-KO strain of T. gondii was successfully obtained.
3.2. Double Deletion of the TR and ROP16 Genes in T. gondii Decreases Growth Capacity In Vitro
To investigate whether the double gene deletion of TR and ROP16 affected the growth of T. gondii RH strain in vitro, plaque assays were performed on the monolayers of Vero cells in six-well plates, and the number and size of the plaques were analyzed. Compared with the data for the RH or single-gene deletion strains, the growth of the TR-ROP16-KO strain was significantly reduced (Figure 2a). To assess the proliferative ability of the TR-ROP16-KO strain in vitro, we measured the number of tachyzoites in parasitophorous vacuoles at 36 h after infection. The TR-ROP16-KO strain had significantly fewer tachyzoites than other strains (Figure 2b,c). Next, to evaluate the invasive ability of the TR-ROP16-KO strain in vitro, the parasites were labeled with a fluorescent probe, and the flow cytometry results indicated that the invasion rate of the TR-ROP16-KO strain was remarkably decreased (Figure 2d). These results implied that the double deletion of TR and ROP16 affected the growth of T. gondii.
3.3. Double Deletion of the TR and ROP16 Genes Changes T. gondii Virulence in Mice
To evaluate the role of TR and ROP16 in the virulence of T. gondii, 1000 tachyzoites of the TR-ROP16-KO, TR-KO, ROP16-KO, and RH strains were injected intraperitoneally into Kunming mice. The mice infected with the RH strain died within 7 days and those infected with the TR-KO or ROP16-KO strain died within 8 days. However, mice infected with the TR-ROP16-KO strain survived for significantly longer and died within 10.5 days (Figure 3). These results indicate that the double deletion of TR and ROP16 further reduces the virulence of type I T. gondii.
3.4. Double Deletion of the TR and ROP16 Genes in T. gondii Increases Intracellular Oxidative Stress Response
To test whether the double gene deletion of TR and ROP16 affected the production of intracellular ROS, the levels of oxidative stress in the parasites were determined by measuring MDA, T-AOC, and ROS. In parasites of the TR-ROP16-KO strain, we observed an increased accumulation of ROS (Figure 4a), and the MDA level was significantly higher (Figure 4b), whereas the T-AOC level was significantly lower compared with those in the TR-KO, ROP16-KO, and RH strains (Figure 4c). In addition, to explore whether the deletion of the TR and ROP16 genes in T. gondii affected the oxidative stress response of host cells, we evaluated ROS levels in mouse macrophages infected with different T. gondii strains. T. gondii infection elevated reactive ROS levels in macrophages in a steep, time-dependent fashion. Intriguingly, the rate of increase in ROS levels in cells infected with the ROP16-KO strain was higher than that in cells infected with the TR-KO strain. Moreover, ROS levels in cells infected with the TR-ROP16-KO strain were significantly higher than those in cells infected with the single-gene deletion strains (Figure 4d). These studies suggest that the double deletion of the TR and ROP16 genes in T. gondii synergistically leads to an increase in oxidative stress levels in host cells.
3.5. Cytokine Changes in the Host Are Affected by the Deletion of the TR and ROP16 Genes in T. gondii
Cytokines play an important role in the body reaction against T. gondii infections. To address whether the double gene deletion of TR and ROP16 affected the production of cytokines in host cells, RAW264.7, cells were infected with tachyzoites of the RH, TR-KO, ROP16-KO, or TR-ROP16-KO strain for at 2, 4, 6, 8, 12, and 24 h. Cytokine expression levels in mouse macrophages at different time points of infection were detected by RT-qPCR. We observed that T. gondii infection changed IL-12 mRNA levels in host cells. Notably, IL-12 mRNA levels in RAW264.7 cells inoculated with the TR-KO strain were significantly higher than those in the cells inoculated with the ROP16-KO strain at 8 h after infection (Figure 5a). Interestingly, IL-12 mRNA levels in RAW264.7 cells infected with the TR-ROP16-KO strain were significantly increased at 6 to 8 h after infection, compared with IL-12 expression levels in cells inoculated with other strains (Figure 5a). In addition, to determine whether the double deletion of the TR and ROP16 genes affected IL-12 production in the host, the mice were infected with tachyzoites of the RH, TR-KO, ROP16-KO, or TR-ROP16-KO strains, and serum samples were obtained at 0.5, 1, 2, 4, and 6 h post infection for IL-12 detection by ELISA. We found that at 0.5–6 h after infection, IL-12 levels in the mice infected with the TR-KO, ROP16-KO, and TR-ROP16-KO tachyzoites were consistently higher than those in the control group at every detection time point. Furthermore, serum levels of IL-12 in mice infected with TR-ROP16-KO tachyzoites were significantly higher than those in mice from the TR-KO- and ROP16-KO-infected groups (Figure 5b). Surprisingly, during the infection period, IL-12 levels in mice infected with the TR-ROP16-KO strain were increased not only due to the absence of ROP16, but also because of the TR deficiency.
4. Discussion
The effector factors ROP16 and TR of T. gondii play pivotal roles in the protection against host innate immunity and directly affect the consequences of infection and disease development [17,22]. During invasion into host cells, ROP16 is secreted from the rhoptries of T. gondii and rapidly localizes to the host cell nucleus. [23]. Previous studies found that although ROP16 is polymorphic and virulence-dependent [24], deleting the ROP16 gene in the type I strain affected the virulence of the parasite in mice [25], which was consistent with the results of our previous studies. Moreover, preliminary studies in our laboratory also showed that the invasion and proliferation abilities of the ROP16-KO type I strain were weaker than those of the type I RH strain. Furthermore, TR is a key secretory factor protecting the type I strain against host oxidative damage, as it reduces superoxide radicals through a direct redox reaction [26,27]. Deletion of the TR gene in the type I strain likely inevitably dysregulated the redox system. Preliminary studies in our laboratory showed that the invasion, proliferation, and virulence of the type I TR-KO strain were weaker than those of the RH and TR gene-complementary strains (TR-CO) [17]. However, the biological characteristics of the TR-ROP16-KO type I T. gondii strain devoid of both ROP16 and TR remained unknown. Therefore, we examined the invasiveness, proliferation, and virulence of the TR-ROP16-KO strain using the same method. By using flow cytometry and microscopy, we established that the invasive and proliferative abilities of the TR-ROP16-KO strain were significantly lower than those of the TR-KO, ROP16-KO, and RH strains. The data on the survival of infected mice showed that the virulence of the TR-ROP16-KO strain was significantly lower than that of either of the single-gene deletion strains. These results indicate that the TR and ROP16 genes in T. gondii are crucial for maintaining the biological activity of the parasite.
Oxidative stress is mainly caused by an imbalance between ROS production and antioxidant capacity of host cells [28]. Animal cells produce lipid oxides, such as MDA, which is a marker of oxidative stress [29,30]. In addition, cells have various antioxidants that eliminate various ROS produced in the body to protect against oxidative stress [31]. Therefore, the level of oxidative stress can be assessed by measuring the levels of MDA, T-AOC, and ROS in parasite cells. Our previous research showed that the oxidative stress level in the TR-KO strain of T. gondii was significantly higher than that in the wild and TR-CO strains, indicating that TR gene deletion affected the antioxidant capacity of T. gondii [17]. Our present results showed that the extent of oxidative stress in the TR-ROP16-KO strain was significantly higher than those of the wild RH strain as well as of the TR-KO and ROP16-KO strains. Specifically, the TR-ROP16-KO strain had the lowest antioxidant capacity, highest degree of lipid oxidation, and highest ROS level caused by parasites. This suggests that the TR and ROP16 genes of T. gondii may play a synergistic role in resisting ROS damage brought about by host immune cells.
ROS, including H_2_O_2_, superoxide anion, and hydroxyl free radicals, are the byproducts of cellular oxygen metabolism [32,33]. Host immune cells (neutrophils, eosinophils, and macrophages) resist microbial infections by releasing ROS [34,35]. In addition, ROS produced by host innate immune response inhibit the activity of T. gondii in monocytes and production of IFN-γ in macrophages of infected mice [36]. In this study, ROS levels in macrophages inoculated with the TR-ROP16-KO, TR-KO, ROP16-KO, and RH strains were significantly higher than those in the control group, and the highest levels were observed in the cells infected by the TR-ROP16-KO strain between 4 and 12 h post infection. Therefore, we speculated that the high ROS levels observed in macrophages infected with T. gondii strains developed on the wild RH strain background may be caused by the TLR-induced MYD88-IL-12 pathway activation [37]. However, the dramatically increased ROS generation in macrophages inoculated with the TR-ROP16-KO strain may be owing to the deletion of the TR gene of T. gondii, which affected the release of antioxidant substances and conferred an inability to completely eliminate all types of ROS in cells. In addition, the deletion of the ROP16 gene in T. gondii resulted in the accumulation of IL-12 in the host innate immune IL-12-IFN-γ-STAT1 signaling pathway, which indirectly affected ROS levels.
Cytokines play an important role in resistance to T. gondii infection, and immune effector cells trigger the release of ROS through the IL-12-IFNγ-STAT1 pathway to clear the infection of T. gondii [38,39,40]. Previous reports have indicated that ROP16 in T. gondii interferes with IL-12 levels in that signaling pathway to achieve parasitic reproduction in the host [41]. In this study, we found that IL-12 mRNA expression in RAW264.7 cells infected with the ROP16-KO strain was significantly increased at 6 to 8 h after infection, compared with that in cells infected with the RH strain. Moreover, ELISA showed that the IL-12 level in mice infected with ROP16-KO T. gondii was significantly higher than that in the RH strain. These results are consistent with those of the previous studies conducted in our laboratory. Similarly, TR helps resist the action of ROS produced by innate immunity [41]. TR is a component of the antioxidant system of T. gondii that catalyzes the conversion of Trx into its reduced form by consuming NADPH [42], which maintains the Trx redox state and resists free radical damage inflicted by host immune cells [43]. Surprisingly, compared with the effects of the RH strain, we found that IL-12 mRNA expression level in RAW264.7 cells infected with the TR-KO strain was significantly increased at 8 h after infection, and the serum level of IL-12 in mice infected with the TR-KO strain was also significantly elevated. To explain this finding, it is useful to remember that Trx (NADPH-dependent) in host cells takes part in the cytokine-induced denitrosylation and activation of NF-κB [44,45]. Therefore, we speculate that the deletion of the TR gene reduced the consumption of the NADPH substrate in host cells. In RAW264.7 cells infected with the TR-KO strain, a relatively sufficient level of NADPH maintained the activity of NF-κB and activated the production of IL-12, suggesting that TR downregulates IL-12 levels by inhibiting the NF-κB pathway. In addition, we found that IL-12 mRNA expression level in RAW264.7 cells infected with the TR-ROP16-KO strain was significantly increased at 6 to 8 h after infection, compared with the level in the cells infected by the RH, TR-KO, or ROP16-KO strains. Moreover, ELISA showed that IL-12 levels in mice infected with the TR-ROP16-KO strain were significantly higher than those in mice infected with other strains tested. These results suggest that the inflammatory response induced by the simultaneous deletion of TR and ROP16 reached the highest degree, because ROP16 downregulated IL-12 levels through the phosphorylation of STAT3 or STAT6, and TR downregulated IL-12 levels through the inhibition of the NF-κB pathway. Regardless of whether T. gondii TR inhibits NF-κB by consuming host cell NADPH or not, based on the obtained results, we conclude that the deletion of the TR and ROP16 genes of T. gondii synergistically increased IL-12 levels and induced host cells to produce more ROS through the IL12-IFNγ-STAT1 signaling pathway.
5. Conclusions
In conclusion, we found that ROP16 and TR are key virulence factors crucial for the growth of T. gondii in vitro and infectivity in vivo. The double deletion of TR and ROP16 in T. gondii weakened the biological phenotype of the parasite. Interestingly, we found that the double deletion of TR and ROP16 in T. gondii resulted in the lowest antioxidant capacity, highest degree of lipid oxidation, and highest ROS levels in the parasite and host cells. Importantly, we found that TR and ROP16 synergistically downregulated IL-12 levels, likely due to the facilitation of STAT3 or STAT6 phosphorylation by ROP16 and inhibition of the NF-κB pathway by TR. Therefore, the deletion of TR and ROP16 genes of T. gondii synergistically increased IL-12 levels and augmented ROS production through the activation of the IL-12-IFNγ-STAT1 signaling pathway. This study clarifies the mechanism of the reduced virulence of the TR-ROP16-KO strain, providing a theoretical basis for the subsequent development of T. gondii vaccines as well as the prevention and control of toxoplasmosis.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Elmore S.A. Jones J.L. Conrad P.A. Patton S. Lindsay D.S. Dubey J.P. Toxoplasma gondii: Epidemiology, feline clinical aspects, and prevention Trends Parasitol.20102619019610.1016/j.pt.2010.01.00920202907 · doi ↗ · pubmed ↗
- 2Mirza Alizadeh A. Jazaeri S. Shemshadi B. Hashempour-Baltork F. Sarlak Z. Pilevar Z. Hosseini H. A review on inactivation methods of Toxoplasma gondii in foods Pathog. Glob. Health 201811230631910.1080/20477724.2018.151413730346249 PMC 6381540 · doi ↗ · pubmed ↗
- 3Montoya J.G. Liesenfeld O. Toxoplasmosis Lancet 20043631965197610.1016/S 0140-6736(04)16412-X 15194258 · doi ↗ · pubmed ↗
- 4Sanchez S.G. Besteiro S. The pathogenicity and virulence of Toxoplasma gondii Virulence 2021123095311410.1080/21505594.2021.201234634895084 PMC 8667916 · doi ↗ · pubmed ↗
- 5Tonouhewa A.B.N. Akpo Y. Sherasiya A. Sessou P. Adinci J.M. Aplogan G.L. Youssao I. Assogba M.N. Farougou S. A serological survey of Toxoplasma gondii infection in sheep and goat from Benin, West-Africa J. Parasit. Dis.20194334334910.1007/s 12639-018-01076-131406398 PMC 6667587 · doi ↗ · pubmed ↗
- 6Pan M. Lyu C. Zhao J. Shen B. Sixty Years (1957–2017) of Research on Toxoplasmosis in China—An Overview Front. Microbiol.20178182510.3389/fmicb.2017.0182528993763 PMC 5622193 · doi ↗ · pubmed ↗
- 7Lima T.S. Lodoen M.B. Mechanisms of Human Innate Immune Evasion by Toxoplasma gondii Front. Cell Infect. Microbiol.2019910310.3389/fcimb.2019.0010331041194 PMC 6476913 · doi ↗ · pubmed ↗
- 8Hunter C.A. Sibley L.D. Modulation of innate immunity by Toxoplasma gondii virulence effectors Nat. Rev. Microbiol.20121076677810.1038/nrmicro 285823070557 PMC 3689224 · doi ↗ · pubmed ↗