Re-Emergence of Bacteriophages and Their Products as Antibacterial Agents: An Overview
Vipin Chandra Kalia, Sanjay K. S. Patel, Chunjie Gong, Jung-Kul Lee

TL;DR
This paper reviews how bacteriophages, viruses that infect bacteria, are being reconsidered as a promising alternative to antibiotics for treating bacterial infections.
Contribution
The paper provides a comprehensive overview of the re-emergence of bacteriophages and their antibacterial potential in the context of antibiotic resistance.
Findings
Bacteriophages offer advantages over antibiotics, including high specificity and self-replication.
Phage-antibiotic synergy and the risks of phage resistance are discussed as key considerations.
Quorum sensing is highlighted as a potential mechanism influencing phage therapy effectiveness.
Abstract
Microbes possess diverse genetic and metabolic traits that help them withstand adverse conditions. Microbial pathogens cause significant economic losses and around 7.7 million human deaths annually. While antibiotics have historically been a lifesaving treatment, their effectiveness is declining due to antibiotic-resistant strains, prompting the exploration of bacterial predation as an alternative. Bacteriophages (BPhs) have reemerged as antibacterial agents, offering advantages over antibiotics, such as (i) high specificity, (ii) self-replication, and (iii) strong killing capacity. This review explores BPh- and enzyme-based antibacterial strategies for infectious disease treatment, discussing phage–antibiotic synergy, the risks of BPh resistance, and the role of quorum sensing in BPh therapy.
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
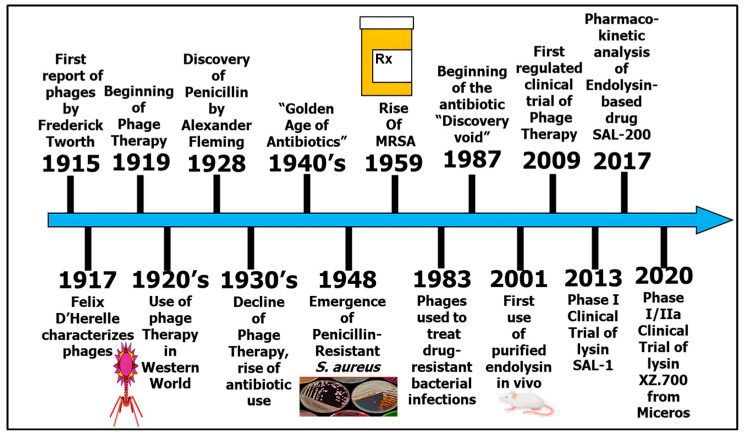 Figure 1
Figure 1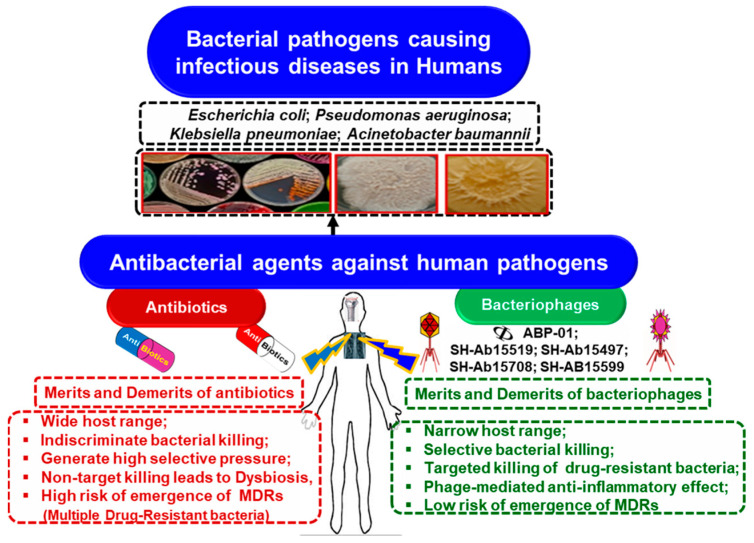 Figure 2
Figure 2| Bacterial Pathogen | Bacteriophage | Antimicrobial Activity | Application/Target | References |
|---|---|---|---|---|
|
| ||||
| ɸAPCEc01, ɸAPCEc02, and ɸAPCEc03 | Complete inhibition of bacterial growth and biofilm formation | Human health | [ | |
| Phage (CA933P) and probiotics (lactobacilli and yeasts) | Cell detachment: 1.2 log CFU; reduced apoptotic cell count by 57.3% | Controlling pathogenic infection and reducing epithelial cell damage | [ | |
| Cocktail (φKMV, φPA2, φPaer4, and φE2005-24-39) | Inhibited biofilm formation and reduced the preformed biofilms | Improving host ranges and using defined cocktails to reduce UTIs | [ | |
| Cocktail of AB-Army1 and AB-Navy1-4 (5 × 109 PFU) | AB-Army1 targeted the capsulated pathogen, and the rest (AB-Navy1-4) lysed the pathogen, preventing the spread of infection and necrosis in a mice wound | Eradicate wound infection in humans | [ | |
| Cocktail of vB_AbaS_D0 and vB_AbaP_D2 (109 PFU/mL) | Improved the therapeutic efficacy and reduced the frequency of phage-resistant bacteria in murine bacteremia (intraperitoneally) | Improved therapeutic efficacy; reduced phage-resistant bacteria | [ | |
|
| vB_SauM_phiIPLA-RODI; vB_SepM_phiIPLA-C1C | 2-log reduction in cell adherence and removed 5 log units of planktonic cells in 8 h | Therapeutic application | [ |
| Phage (108 PFU/mL) | Survival rate of 100% in mice (intravenous) | Public health: anti-drug resistance | [ | |
| Lytic phage EF-P29 (4 × 105 PFU, intraperitoneally) | Protected all mice against bacteremia (2 × 109 PFU/mouse) | For treating life-threatening nosocomial infections and avoiding gut microbiota imbalance | [ | |
| Lytic (EFDG1 and EFLK1) | Rescued mice from severe septic peritonitis (100%) in a mouse model | For treating life-threatening nosocomial infections and avoiding gut microbiota imbalance | [ | |
| SHEF2 | Eradicated biofilms on polystyrene surfaces and on tooth root | Broad-range biocontrol agent against antibiotic-resistant infections | [ | |
| vB_EfaS_PHB08 (105 CFU/cm2) | Reduced bacteria (105 CFU/cm2) on a polystyrene MTP and lettuce as a vegetable model | For treating wounds and UTIs in animals and humans | [ | |
|
| ||||
| vB_SepiS-phiIPLA7 (EPS depolymerase Dpo7; 0.15 μM) | Exopolysaccharide matrix degradation (30%); removal of biofilm-attached cells: 90% | Controls hospital-acquired infections and bacteremia | [ | |
|
| Muralytic enzymes P128 from phage K (≥12.5 μg/mL) | Removal of biofilms up to 95.5% | Infectious conditions (chronic rhinosinusitis) | [ |
| Lysin CF-301 (≤0.25 μg/mL) | Eradication of biofilm within 1 h and bacterial killing within 6 h on catheters, surgical mesh, glass, and polystyrene surfaces | Treating staphylococcal infections | [ | |
| Endolysin LysEFm5 (from IME-EFm5; 16 to128 mg/mL) | Lysis of 19/23 pathogenic isolates in actively growing cells | Highly specific against MDR Gram-positive pathogens | [ | |
| Endolysin lys08 (from vB_EfaS_PHB08; 5 µg) | Eradicated the biofilm on a polystyrene MTP | For treating wounds and UTIs in animals and humans | [ | |
| Pathogenic Bacteria | Bacteriophage (BPh) | Disease and Patient Details | Mode of Administration | Outcomes | References |
|---|---|---|---|---|---|
| Cocktail BFC1 (2 BPhs) | Septicemia (acute kidney injury, 61-year-old male) | Intravenous (IV) (109 PFU/mL, every 6 h for 10 days) | Recovery from septicemia achieved after 10 days of treatment | [ | |
|
| OMKO1 | Aortic graft infection (76-year-old male) | IV (107 PFU/mL) + ceftazidime | Pathogen eradication within four weeks, with no recurrence reported after two years | [ |
| Cocktail (2 BPhs) | Bacteremia (2-year-old male child) | IV (3.5 × 105 PFU/mL, every 6 h for 3 days) + antibiotics | Symptoms returned after stopping therapy | [ | |
| Cocktail | Bone metastasis (60-year-old male with lung cancer) | Applied on bone in the cavity (108 PFU/mL) | Positive results lasting 18 months | [ | |
| Cocktail of | Prosthetic joint infection (80-year-old obese woman with type 2 diabetes mellitus and chronic kidney injury) | Intraoperative injection in the joint cavity | Positive results lasting 18 months | [ | |
| Cocktail AB-PA01 (4 lytic BPhs) | Pneumonia in cystic fibrosis (CF, 26-year-old female) | IV (4 × 109 PFU/mL, every 6 h for 8 weeks) | Recovery with no recurrence of pneumonia after 100 days | [ | |
|
| Cocktail AB-PA01 (4 lytic BPhs) | Ventilator-associated pneumonia and empyema (77-year-old female) | IV (109 PFU/mL, every 12 h for 7 days) and nebulized | No adverse effects | [ |
|
| Lytic BPhs (BAP-5phi1, PFU +MTAE-8 phi1, and PFU + MTAE-8 phi3) + antibiotics (meropenem, ceftezidine/avibactam + aztreonam) | HeartMate II driveline infection; recurrent bacteremia | IV (1.5 × 106 PFU + 2.2 × 109 PFU + 2.1 × 107 PFU in 1 mL every 8 h for 6 weeks) | Lytic BPhs allowed the patients to proceed with heart transplantation successfully | [ |
| Cocktail of | Chronic osteomyelitis | Intraoperatively (107 PFU/mL, every 8 h for 7–10 days) | No recurrence or severe side effects up to 16 months | [ | |
| Knee periprosthetic joint infection and chronic osteomyelitis (80-year-old woman with metabolic syndrome, diabetes, etc.) | IV (108 PFU/mL, every 8 h for 5 days) | Patient achieved notable recovery and mobility after 10 months | [ | ||
| Cocktail ϕPC followed by cocktail ϕ IV (AB-Navy1, AB-Navy4, AB-Navy71, and ABNavy97) | Diabetes (68-year-old patient) | IV (109 PFU/mL for 36 h followed by increasing frequency for 2 days for period of 11 weeks) + antibiotics | Rapid recovery over 12 weeks | [ | |
| Cocktail of ɸAbKT21phi3 and ɸKpKT21phi1 | Bone infection (42-year-old male) | IV (5 × 107 PFU/mL, every 8 h for 5 days) | BPhs and antibiotics resulted in avoiding amputation | [ | |
|
| UTI–epididymitis (58-year-old male) | Administered orally and into the bladder through catheter; treatment continued for 12 weeks along with antibiotics | Eradicated a recurrent UTI, showing high efficacy one year later | [ | |
| Cocktail of lytic BPhs (Kp152, Kp154, Kp155, Kp164, Kp6377, and HD001; 5 × 108 PFU/mL of each BPh) + sulfamethoxazole–trimethoprim | Recurrent UTI (63-year-old male) | Intravesical phage administration and oral antibiotics; bladder irrigation every 12 h for 5 days | Remained infection-free at six months | [ | |
| Lytic BPh | Recurrent obstructive nephrolithiasis | Administered orally and via intra-rectal modes over a 3-week cycle | No adverse effects | [ | |
|
| Staphylococcal BPh Sb-1 | Diabetic foot ulcers (44–92 years old, 6 males) | Topical (0.1–0.5 mL of 107–108 PFU/mL) | Effectively treated poorly vascularized toe ulcers in nine patients over seven weeks | [ |
|
| Cocktail AB-SA01 (3 lytic BPhs: Sa87, Sa83, and Sa36) + antibiotics (cefazolin + minocycline) | HeartMate II driveline; sternal osteomyelitis; recurrent bacteremia | IV (3 × 109 PFU in 1 mL every 12 h for 4 weeks) | Allowed the patients to proceed with heart transplantation successfully | [ |
| Cocktail (3 lytic BPhs: Muddy, ZoeJ Δ | CF (15-year-old female) | IV (109 PFU in 5 mL, every 12 h for 32 weeks) | Stabilized post-lung transplantation in cystic fibrosis patients | [ | |
| Antibiotic-resistant bacteria | Cocktails (MS-1 and OPMS-1): | 62 patients, infections of the genitourinary tract, prostatitis, bone, respiratory tract, skin, or soft tissue | Intra-rectal or local (106–109 PFU/mL, every 8–12 h for 12 weeks or more) | 40–55% of patients showed positive responses | [ |
- —Basic Science Research Program through the National Research Foundation of Korea (NRF), funded by the Ministry of Science, ICT & Future Planning
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsBacteriophages and microbial interactions · Plant Virus Research Studies · Monoclonal and Polyclonal Antibodies Research
1. Introduction
The discovery of bacteriophages (BPhs) in the early 20th century sparked interest in their potential as antibacterial agents [1,2,3]. However, inconsistent results and the rise of antibiotics limited their early use [4]. With the emergence of drug-resistant bacteria and declining antibiotic development [5], interest in BPh therapy (BPhT) has resurged (Figure 1) [1]. BPhs offer key advantages, including specificity, self-replication, rapid adaptation, biofilm clearance, high efficacy, and cost effectiveness [2,3,6]. Despite their known potential [7,8], only a few former USSR countries have approved BPh-based treatments for human use [9]. The global demand for new BPhs remains high due to evolving pathogens and regulatory challenges [10].
Bacterial infections significantly contribute to morbidity and cause 7.7 (5.7–10.2) million deaths annually [11,12]. Notably, 65–80% of chronic infections involve biofilm-forming bacteria, making treatment more challenging. Key pathogens include Gram-negative bacteria (e.g., Acinetobacter, Escherichia, Klebsiella, and Pseudomonas), responsible for respiratory and urinary infections, and Gram-positive bacteria (e.g., Enterococcus, Staphylococcus, and Streptococcus), which cause soft tissue infections [13,14,15,16]. Biofilm removal strategies focus on preventing bacterial adhesion, inducing detachment, and disrupting biofilm regulation through quorum sensing (QS) inhibitors (QSIs) and matrix degradation [17,18].
The urgent need for alternative antibacterial approaches has intensified. While biofilm inhibition methods are well documented [19], recent reviews rarely discuss BPhs as tools against infectious pathogens [20]. This review examines BPh-based antibacterial strategies (Figure 2), including phage–antibiotic synergy, the risks of BPh resistance, and the role of QS in BPhT.
A bibliographic analysis was conducted, reviewing research articles and review papers indexed in Scopus, PubMed, and Google Scholar. The search utilized keywords including biocontrol, biofilm, bacteria, bacteriophage, phage enzymes, pathogens, therapy, phage cocktails, antibiotics, antibacterial agents, infectious diseases, phage resistance, quorum sensing, horizontal gene transfer, and endolysins. Approximately 800 studies, primarily from the last decade, were critically evaluated to synthesize the key findings and advancements in the field.
2. Bacteriophages as Antibacterial Agents
Bacteriophages (BPhs) infect bacteria through lytic and lysogenic cycles [21]. In the lytic cycle, phages hijack bacterial machinery, replicate, and lyse the host cell, making them ideal for therapeutic use. In contrast, the lysogenic cycle involves phage DNA integrating into the bacterial genome, remaining dormant. While lysogenic phages pose risks due to horizontal gene transfer (HGT) of resistance or toxin genes, they hold potential for bacterial genetic engineering. Strictly lytic phages are essential for clinical applications to ensure safety and effectiveness. Prophages can also provide immunity against superinfections [21]. Lytic BPhs interact with biofilms at different stages [22,23] using three main antibacterial strategies: (i) whole phages, (ii) bacteriophage-derived enzymes (e.g., exopolysaccharide polymerases and peptidoglycan hydrolases), and (iii) phage–antibiotic combinations.
Early research on monophage therapy showed promising results (Table 1). A few examples are that (i) the BPh EF-P29 prevented bacteremia and alleviated gut dysbiosis in a vancomycin-resistant Enterococcus faecium (VREF) murine model [24], (ii) the BPh SHEF2 eradicated Enterococcus faecalis biofilms and improved survival in zebrafish infected with E. faecalis OS16 [25], and (iii) lytic phages significantly reduced bacterial loads, with phage ph0034 decreasing counts by 7.5 log CFU/mL and ph0031 by 5.1 log CFU/mL within 24 h [26]. Monophage therapy also showed efficacy against Staphylococcus aureus infections: (i) In a mouse S. aureus lung infection model, lytic BPhs achieved 100% survival within 24 h, compared to 62% with clindamycin alone and 75% with combination therapy [27]. (ii) The BPh SLPW reduced inflammation in methicillin-resistant S. aureus (MRSA)-infected mice [28]. (iii) Georgian Staphylococcus BPhs have successfully treated diabetic foot ulcers and S. aureus infections [29].
Phage cocktails enhance treatment consistency by preventing biofilm formation and reducing BPh-resistant mutants [30] (Table 1). Successful applications include the following: (i) reducing epithelial cell damage and apoptosis in enterohemorrhagic Escherichia coli EHEC O157 infection by 57.3% [31], (ii) reducing E. coli ST131-H30R gut colonization [32], (iii) inhibiting biofilm formation in levofloxacin-resistant Pseudomonas aeruginosa [33], (iv) treating Acinetobacter baumannii wound infections and preventing necrosis [34], (v) rescuing mice from acute and chronic bloodstream infections [35], (vi) enhancing efficacy against A. baumannii with specific BPs [36]; (vii) reducing inflammation in VREF-induced septic peritonitis [37]; (viii) phages targeting E. faecalis and E. coli improved gut microbiota in murine colitis models [38], and (ix) the Kayvirus phages SAM1 and SAM2 in the Fersisi cocktail effectively infected MRSA strains, although host gene expression changes require further study [39]. Despite their advantages, phage therapy faces challenges, including phage resistance, HGT risks, dysbiosis, cross-resistance, and high production costs [40].
3. Bacteriophage Enzymes as Antibacterial Agents
Phage enzymes target bacterial surface structures, such as lipopolysaccharides, exopolysaccharides (EPSs), and capsular polysaccharides [45]. Examples include (i) Pseudomonas putida phage 815 tail spike proteins binding and degrading bacterial capsules [46,47], (ii) phage polymerases reducing biofilms by 37% within 24 h at an MOI of 10^6^ [48], (iii) S. epidermidis phage phiPLA7 depolymerase degrading 30% of the EPSs in S. aureus biofilms [49], and (iv) phage ϕAB6 tail proteins degrading A. baumannii EPSs, demonstrating therapeutic potential [50]. Peptidoglycan-degrading enzymes offer high specificity and efficacy, independent of bacterial resistance mechanisms [51]. These include glycosidases, lysozymes, amidases, and endopeptidases [52]. A few examples are as follows: (i) murein hydrolase P128 and lysostaphin from BPh K disrupted 95.5% of S. aureus sinus biofilms at ≥12.5 μg/mL [42], (ii) endolysin LysH5, CHAPk peptidase, and lysin ClyH effectively degraded S. aureus biofilms [53,54,55], (iii) endolysin Lys68 combined with a membrane permeabilizer reduced Salmonella Typhimurium by 1 log unit [56], (iv) the BPh lysin CF-301 eradicated 90% of S. aureus biofilms within one hour at ≤0.25 μg/mL [43], (v) LysEFm5 from the BPh IME-EFm5 showed activity against vancomycin-resistant E. faecium without requiring Zn ions [44], and (vi) BPh vB_EfaS_PHB08 endolysin reduced E. faecalis by 10^5^ CFU, showing promise for wound and UTI treatments [41].
Engineered chimeric BPhs are being developed to comply with regulatory requirements, easing the approval process for phage cocktails. Key strategies to enhance BPh efficacy include (i) avoiding identical receptor targets, (ii) using diverse phage species, and (iii) introducing novel BPhs to counteract resistance [9,57]. A low resistance rate to phage lytic proteins has been observed, supporting their potential as future therapeutic agents [58,59].
4. Complementing Phages with Antibiotics
BPhs enhance antibiotic efficacy through synergistic effects, improving outcomes using the BPh Henu2 [60], against MRSA [61], and in dental infections [62]. Examples include the following: (i) a Burkholderia cepacia phage with low-dose meropenem improved Galleria mellonella survival [63], (ii) phage–ciprofloxacin reduced P. aeruginosa load by 10,000-fold in endocarditis-infected rats [64], (iii) ciprofloxacin post-BPh infection eliminated E. coli more effectively than antibiotics or phages alone [65], (iv) the lytic S. aureus phage Sb-1 with oxacillin exhibited synergy [61], (v) daptomycin combined with E. faecium phage cocktail showed limited efficacy [66], (vi) phages 6 and 45 with gentamicin reduced P. aeruginosa by 3 logs in 12 h [60], (vii) phage MRM57 with cefotaxime showed synergy against Citrobacter amalonaticus [67], and (viii) flucloxacillin enhanced S. aureus biofilm suppression in rats (2.15 log CFU/g reduction) [68]. Although PAS (phage–antibiotic synergy) is generally beneficial, some antagonistic interactions occur [69,70].
PAS is regulated through multiple mechanisms. Certain antibiotics stimulate phage replication, increasing progeny release. Quinolones and β-lactams induce bacterial elongation, enhancing susceptibility to phage lysis enzymes in Yersinia enterocolitica and E. coli [71]. Ceftriaxone combined with a Siphoviridae phage inhibited P. aeruginosa cell wall synthesis by triggering sulA gene activation, leading to filamentation and increased phage assembly [72]. Some antibiotics enhance the plaque and burst size, e.g., the T4 phage burst size increased 5-log at low cefotaxime levels, reducing its latent period [73]. PAS reduces resistant mutants, as seen with ciprofloxacin/daptomycin and S. aureus phage Sb-1 [64,74]. Phage–antibiotic interactions enhance antibiotic susceptibility by depleting lysogens [75], re-sensitizing pathogens [76], and reducing resistance [77].
4.1. PAS Against Bacterial Biofilms
Phage–antibiotic combinations effectively eradicate biofilms: (i) a T4 phage with cefotaxime lowered the minimum eradication concentration for E. coli biofilms [73], (ii) T4 and PB-1 phages with antibiotics eliminated 60–99% of biofilm biomass and reduced phage-resistant cells by 39–99% in E. coli and P. aeruginosa infections [78,79], (iii) ciprofloxacin-resistant E. coli biofilms were cleared using the phage ɸWL-3 and fosfomycin [80], (iv) the phage vB PmiS-TH with ampicillin significantly reduced P. mirabilis biofilms, with greater effects at higher antibiotic doses and phage MOI [81], and (v) phage cocktails showed effectiveness against MDR A. baumannii biofilms in a murine model [82]. These findings underscore PAS’s potential in eradicating bacterial biofilms and combating antibiotic resistance.
4.2. In Animal Models
Phage–antibiotic synergy (PAS) has shown efficacy in animal infection models:
- In rat osteomyelitis, the P. aeruginosa PAT14 and S. aureus Sb-1 phages combined with antibiotics significantly reduced biofilm formation [83].
- In a mouse post-arthroplasty model, the S. aureus MR-5 phage (10^9^ PFU/mL) and linezolid (5%) lowered the bacterial load by day 10 [84].
- For K. pneumoniae pneumonia, the phage P-KP2 with gentamicin increased survival to 70% in mice [85].
- In neutropenic mice with P. aeruginosa lung infections, the PEV20 phage (10^6^ PFU/mg) and ciprofloxacin (0.33 mg/mg) led to a 5.9-log bacterial reduction [86].
However, PAS was less effective in some cases: (i) in a rat MRSA model, linezolid monotherapy reduced mortality by 38%, while phage treatment had variable survival rates [87]; (ii) a S. aureus phage cocktail with daptomycin in MRSA pneumonia showed limited efficacy (50–55% survival) [88]. PAS outcomes vary based on the antimicrobial type, dosage, and treatment timing.
4.3. In Humans
4.3.1. Case Reports
BPhT has been applied to treat diverse bacterial infections (Table 2):
-
A 2-year-old child with P. aeruginosa sepsis and congenital heart disease improved with a phage–antibiotic combination, although their symptoms recurred post-therapy [89].
-
A post-aortic aneurysm repair patient with ciprofloxacin-resistant P. aeruginosa was treated with the phage OMKO1 (10^7^ PFU/mL) and ceftazidime, eradicating the pathogen within four weeks, with no recurrence for two years [90,91].
-
In seven cardiothoracic patients (ages 13–66) with MDR infections, phage therapy was effective and well tolerated [92].
-
In a 61-year-old man with E. cloacae peritonitis and MDR P. aeruginosa septicemia, BFC1 improved his renal function, although he later died of K. pneumoniae sepsis [93].
-
A renal transplant patient’s recurrent K. pneumoniae UTI was successfully treated with a personalized phage [94].
-
A 63-year-old man with a UTI from XDR K. pneumoniae was treated with a lytic BPh cocktail, remaining infection-free at six months [95].
-
Another patient with MDR K. pneumoniae gut colonization received an oral and intra-rectal lytic BPh, showing no adverse effects [96].
-
In cystic fibrosis (CF) cases, the following was found:
-
○A 17-year-old infected with B. cepacia and Achromobacter xylosoxidans showed improved lung function (FEV1 from 1.83L to 3.33L) after inhaled phage therapy [97].
-
○A 26-year-old patient with MDR P. aeruginosa recovered following eight-week phage cocktail (AB-PA01) treatment and antibiotics, with no pneumonia recurrence after 100 days [98].
-
○Engineered phage therapy stabilized post-lung transplant CF patients with Mycobacterium abscessus infections [99].
-
A necrotizing pancreatitis patient with A. baumannii pseudocyst infection recovered following IV phage treatment after resistance to initial cocktails [100].
-
Ventilator-associated pneumonia and empyema patients tolerated a four-phage cocktail well [101].
-
Phage Sb-1 effectively treated S. aureus toe ulcers in nine patients over seven weeks [29].
-
BPhT, used alongside systemic antibiotics, successfully treated biofilm infections from ventricular assist devices, enabling heart transplants in MSSA and P. aeruginosa cases, although serum-neutralizing activity and BPh titers require further study [102].
-
In a study with 62 infected patients and 30 healthy volunteers, the sera’s anti-phage activity did not influence outcomes, although 40–55% of patients showed positive responses [103].
-
In critically ill patients with S. aureus infections treated with a BPh (AB-SA01), the inflammatory markers decreased over 90 days. Future studies will use a 12 h dose of 10^9^ PFU/mL to assess the BPhT’s efficacy [104].
-
In an 80-year-old woman with prosthetic joint infections (P. aeruginosa and S. aureus), phage therapy yielded lasting positive results for 18 months [105].
-
For severe musculoskeletal infections, BPh cocktails were administered intraoperatively every eight hours for ten days alongside antibiotics, with no recurrence or severe side effects for up to sixteen months [106].
-
Personalized BPhs as adjuvants to antibiotics successfully treated P. aeruginosa infections in periprosthetic joints, allowing the patient to regain mobility within ten months [107].
-
A 42-year-old patient with bone infections from XDR A. baumannii and MDR K. pneumoniae avoided amputation with phage therapy [108].
A craniectomy patient with A. baumannii infection received a lytic phage cocktail, but rapid phage clearance led to treatment withdrawal; the patient died on day 20 [109].Lung transplant patients with Burkholderia dolosa and P. aeruginosa (MDR) were treated using phages. The P. aeruginosa patients recovered, but those with B. dolosa infections relapsed, leading to mortality [110].A critically ill patient with respiratory infection caused by A. baumannii improved after 35 days of treatment with the phage AbW4878Ø1 (1 × 10^9^ PFU/mL) and broad-spectrum antibiotics [111].
While phage therapy shows promise, factors such as serum-neutralizing activity, dosing, and pathogen susceptibility require further study.
4.3.2. Clinical Trials
The PhagoBurn trial tested a 12-phage cocktail (PP1131) for P. aeruginosa-infected burn wounds. The low-dose treatment (1 × 10^6^ PFU/mL) took 144 h to meet the primary endpoint, compared to 47 h for silver sulfadiazine. The trial was terminated early due to poor efficacy, likely due to inadequate phage dosage (200–2000 PFU vs. the typical 2–3 × 10^7^ PFU/mL) [112].A trial on E. coli-associated diarrhea in children was abandoned due to ineffectiveness. A follow-up study on 79 children tested two oral phage cocktails (M: 1.4 × 10^9^ PFU/mL, T: 3.6 × 10^8^ PFU/mL) over four days, showing no adverse effects but comparable efficacy to the placebo. Likely, the issues included gastric degradation and poor intestinal delivery, highlighting the need for improved formulations for E. coli in Asia [113,114].A trial on MRSA bacteremia and endocarditis evaluated exebacase, an anti-staphylococcal lysin. A single IV dose with standard antibiotics led to superior clinical responses and good tolerability in MRSA patients compared to antibiotics alone [115,116].Of the numerous phase I/II BPhT studies, only two have reached phase III: (i) nebulized pyoBPh complex [117] and (ii) post-transurethral prostate resection intervention for UTI patients [118]. Some recent trials have shown contradictory outcomes [119].A phase I trial on chronic rhinosinusitis tested the intranasal phage cocktail AB-SA01 (up to 3 × 10^9^ PFU/mL) over two weeks in a tertiary center, demonstrating safety and efficacy. These findings suggest phages as potential antibacterial agents for this condition [120].
5. Limitations
BPhs are proposed as an alternative to antibiotics but require efficacy validation before clinical trials. Key challenges include phage resistance, gut microbial dysbiosis, HGT, and cross-resistance.
5.1. High Specificity and Microbial Documentation
BPhs target specific bacterial strains, minimizing microbiota disruption but limiting broad therapeutic application [50,51]. Effective treatment requires precise pathogen identification using advanced diagnostics like MALDI-TOF, whole-genome sequencing, or culture-based methods [121,122]. Pre-treatment phage testing (e.g., plaque assays and biofilm disruption studies) ensures infectivity and lytic activity against clinical isolates. Customized phage cocktails may be needed for polymicrobial infections or rapidly evolving pathogens [123].
BPh infections can alter bacterial metabolism, as seen in S. aureus biofilms treated with phiIPLA-RODI, which showed reduced peptidoglycan biosynthesis, potentially affecting virulence and resistance [124,125]. The gut microbiome, containing ~10^12^ viruses (mostly BPhs), facilitates HGT, potentially spreading antibiotic resistance and virulence factors [126,127]. While temperate phages contribute to HGT, their role in human health remains unclear [128,129]. Gut microbiome alterations have been linked to obesity, diabetes, immune disorders, and cardiovascular diseases [130,131,132,133].
BPh safety is evaluated by its impact on gut microbiota and host tissues. While BPhs can eliminate antibiotic-resistant bacteria, endotoxin release may trigger immune responses. No severe adverse effects have been reported [134]. The phage vB_KpnM_GF effectively controlled a K. pneumoniae outbreak, suggesting potential for decolonizing multidrug-resistant pathogens [135].
5.2. Dose, Interval, and Administration Optimization
Challenges in BPh therapy (BPhT) include formulation, propagation, and stability. There is insufficient knowledge on maintaining phage viability at room temperature, affecting their shelf life and usability. Delivery methods like inhalation (for respiratory infections) and oral administration (for gastrointestinal infections) require formulations that protect phages from degradation in physiological conditions [136,137].
To optimize BPhT, the focus has shifted to purified phage enzymes like endolysins and cell wall hydrolases, which offer enhanced bacterial targeting and reduced gene transfer risks [48]. Regulatory clarity on BPhT practices is essential for clinical application [138,139].
5.3. Regulatory and Safety Considerations
Phages, being “living” entities, complicate regulatory approval by agencies like the FDA and EMA [140]. While phage therapy is used in parts of Europe, standardized guidelines are lacking [141,142]. Pharmacokinetic data on endolysins are limited, and while BPh-related side effects (e.g., skin allergies) are rare, further study is needed [143].
Lytic BPhs are preferred over temperate ones to prevent virulence and resistance gene transfer. Phage cocktails with EPS polymerases improve biofilm targeting [144]. The immune system’s response, particularly anti-phage antibodies, can affect therapy outcomes, as seen in phage MS-1 treatment, where 23% of patients with pre-existing antibodies showed reduced efficacy [145,146].
Endolysins degrade bacterial cell walls via glycosidases, amidases, or endopeptidases, leading to osmotic lysis. Unlike BPhs, they degrade quickly, reducing gene transfer risks. Studies on PaI and CpI-1 lysins showed no immune-related side effects, stable catalytic activity, and no toxicity [147,148]. Recombinant phage proteins, such as thermostable endolysins, hold promise but require further safety and immunogenicity studies [149].
5.4. Emergence of BPh-Resistant Bacteria
Like antibiotics, BPhs can drive bacterial resistance. Viral fitness factors may enhance bacterial survival, promoting resistance to both antibiotics and phages [150,151]. In P. aeruginosa, mutations in hmgA and galU affected O-antigen expression, preventing phage adsorption [152]. In L. monocytogenes, a loss of cell wall teichoic acid residues due to phage A511 infection reduced bacterial fitness [153,154]. However, resistant mutants could aid in developing live attenuated vaccines [155].
CRISPR/Cas-mediated immunity also enables bacteria to recognize and clear phages, although this reduces bacterial fitness at low CRISPR array scales [156,157,158,159]. Biofilms hinder phage adsorption by creating physical barriers, while competitive inhibitors may block phage receptors [160]. S. aureus infection by phiIPLA-RODI induced biofilm formation through altered peptidoglycan biosynthesis, conferring resistance. Thus, lytic phages that release plasmid DNA should be avoided in BPhT [124,161].
6. Emerging Role of Bacterial QS in Bacteriophage Infection
Quorum sensing (QS), a key bacterial communication system, regulates biofilm formation, phage adsorption, the lytic–lysogenic cycle, genetic exchange, and phage–host coevolution [162,163]. QS exerts dual effects on phage infections, either inhibiting or promoting them. For example, in Vibrio anguillarum, QS inhibits phage infection by reducing phage receptor expression. A ∆vanT mutant exhibits increased OmpK receptor expression at low cell densities, promoting biofilm formation and phage resistance. Conversely, at high cell densities, the ∆vanO mutant suppresses prophage induction and enhances proteolysis, preventing biofilm formation [164,165].
QS activity is quantified using (1) chemical assays (e.g., HPLC, GC-MS, fluorescence/absorbance assays, and bioassays), (2) gene expression analysis (e.g., reporter gene systems, qRT-PCR, and RNA-Seq), (3) phenotypic assays (e.g., biofilm quantification via crystal violet staining and motility assays), and (4) mathematical modeling to determine QS dynamics [166].
QS molecules such as N-acyl-homoserine lactone (AHL), CAI-1, and AI-2 modulate phage resistance by downregulating lipopolysaccharide (LPS) O-antigens, reducing Vibrio cholerae susceptibility to phages [167]. Phage infection can also activate QS genes, enhancing biofilm formation in P. aeruginosa and E. coli [168]. Indole, a QS signal, inhibits phage infection in P. aeruginosa by downregulating the genes essential for type IV pilus (T4P) assembly, reducing phage adsorption [169]. Similarly, in Shewanella baltica, QS decreases phage adsorption via LuxR-mediated LPS receptor downregulation [170].
Phage–QS interactions remain underexplored. Phages influence QS-regulated virulence throughout infection [171]. In P. aeruginosa, QS upregulates bci, impacting motility, biofilm formation, and pyocyanin production, critical for infections in CF patients [172]. Additionally, las-QS in P. aeruginosa PAO1 enhances phage infection by upregulating galU, increasing phage-mediated killing efficiency [173]. In V. cholerae, phages regulate QS-controlled biofilm genes [174] (Table 3).
Cyclic-di-GMP (c-di-GMP) modulates biofilm dynamics, and the phage PB1 disrupts biofilms by producing anti-c-di-GMP peptides [175]. QS inhibitors (QSIs) prevent biofilm formation, increasing bacterial susceptibility to phages. However, when QS promotes phage infection, QSIs may reduce phage therapy efficacy. A sequential strategy using QSIs and phages is advisable when QS does not enhance phage infection, facilitating bacterial eradication at lower antibiotic doses [17].
Future research should critically evaluate QS-mediated biofilm formation, a major barrier to effective phage therapy. Strategies focusing on bacterial adhesion prevention, biofilm matrix degradation, and QS-targeted interventions require extensive investigation to enhance BPhT efficacy.
7. Conclusions
Bacteriophage therapy (BPhT) emerged nearly a century ago but was overshadowed by antibiotics. The rise of multidrug-resistant pathogens and stagnation in antibiotic development have renewed interest in alternatives like QSIs. Although promising against various infections (e.g., respiratory, urinary tract, burn wounds, and endocarditis), BPhT lacks regulatory approval as a standard treatment. Studies highlight the potential of PAS in reducing antibiotic use and limiting resistance, but further research is needed to clarify synergistic and antagonistic effects. Addressing these challenges could integrate BPhT into clinical practice as a vital tool in the post-antibiotic era.
A critical research focus should include QS and QSI strategies targeting bacterial adhesion, polysaccharide production, and biofilm degradation. Combining QS-targeted interventions with phage therapy can significantly improve treatment efficacy and clinical outcomes.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Murray E. Draper L.A. Ross R.P. Hill C. The advantages and challenges of using endolysins in a clinical setting Viruses 20211368010.3390/v 1304068033920965 PMC 8071259 · doi ↗ · pubmed ↗
- 2Opperman C.J. Wojno J.M. Brink A.J. Treating bacterial infections with bacteriophages in the 21st century S. Afr. J. Infect. Dis.20223734610.4102/sajid.v 37i 1.34635399556 PMC 8991297 · doi ↗ · pubmed ↗
- 3Shkoporav A.N. Turkington C.J. Hill C. Mutualistic interplay between bacteriophages and bacteria in the human gut Nat. Rev. Microbiol.20222073774910.1038/s 41579-022-00755-435773472 · doi ↗ · pubmed ↗
- 4Wu S. Zachary E. Wells K. Loc-Carrillo C. Phage therapy, future inquiries Postdoc J.20131243510.14304/SURYA.JPR.V 1N 6.328286802 PMC 5342839 · doi ↗ · pubmed ↗
- 5Kalia V.C. Rani A. Lal S. Cheema S. Raut C.P. Combing databases reveals potential antibiotic producers Expert Opin. Drug Discov.2007221122410.1517/17460441.2.2.21123496078 · doi ↗ · pubmed ↗
- 6Principi N. Silvestri E. Esposito S. Advantages and limitations of bacteriophages for the treatment of bacterial infections Front. Pharmacol.20191051310.3389/fphar.2019.0051331139086 PMC 6517696 · doi ↗ · pubmed ↗
- 7Kakasis A. Panits A.G. Bacteriophage therapy as an alternative treatment for human infections. A comprehensive review Int. J. Antimicrob. Agents.201953162110.1016/j.ijantimicag.2018.09.00430236954 · doi ↗ · pubmed ↗
- 8Ling H. Lou X. Luo Q. He Z. Sun M. Sun J. Recent advances in bacteriophage-based therapeutics, Insight into the post-antibiotic era Acta Pharm. Sin. B 2022124348436410.1016/j.apsb.2022.05.00736561998 PMC 9764073 · doi ↗ · pubmed ↗