Immature Stages and Breeding Behaviour of the Hollyhock Weevil–Rhopalapion longirostre (Olivier 1807) (Coleoptera, Brentidae, Apioninae)
Rafał Gosik

TL;DR
This paper describes the immature stages and breeding behavior of the hollyhock weevil, Rhopalapion longirostre, and its ecological interactions.
Contribution
The first comprehensive description of preimaginal stages and new insights into the species' reproductive behavior and niche competition.
Findings
Two morphological forms of pupae were identified based on body size measurements.
The weevil's range expansion is linked to climate change and global warming.
Competition with Pexicopia malvella in hollyhock fruit limits R. longirostre populations.
Abstract
New observations on the reproductive behaviour of hollyhock weevil–Rhopalapion longirostre (Olivier, 1807) are presented. The morphology of the preimaginal stages of this species is comprehensively described for the first time. The pupal measurements indicate the existence of two morphological forms: a very considerable number of relatively large individuals (body length: 3.00–3.50 mm) and a few very small (1.75, 2.30 mm) individuals. Competition between R. longirostre and the hollyhock seed moth—Pexicopia malvella (Hübner, (1805))—for the development niche is described. New information is given on the distribution of R. longirostre as well as evidence confirming the connection between global warming and its expansion. Since the 1960s, the range of Rhopalapion longirostre has rapidly expanded from central Asia to almost all of Europe; it has also been introduced to North America. This…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
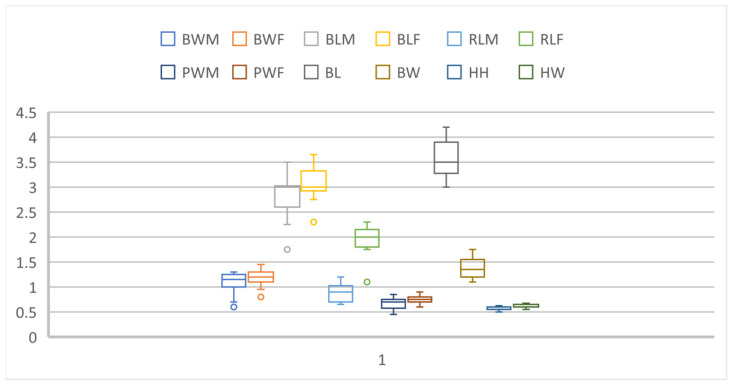 Figure 1
Figure 1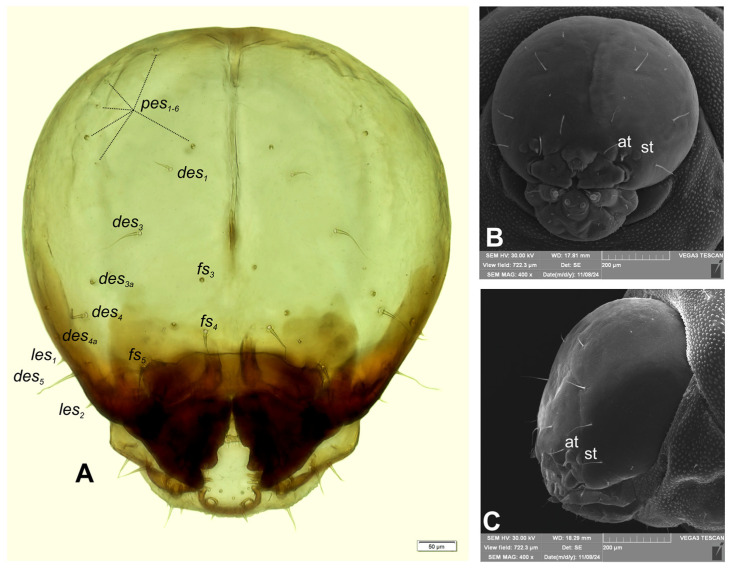 Figure 2
Figure 2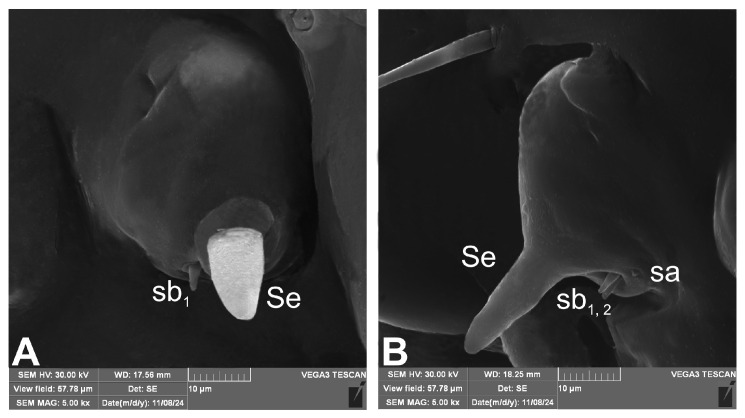 Figure 3
Figure 3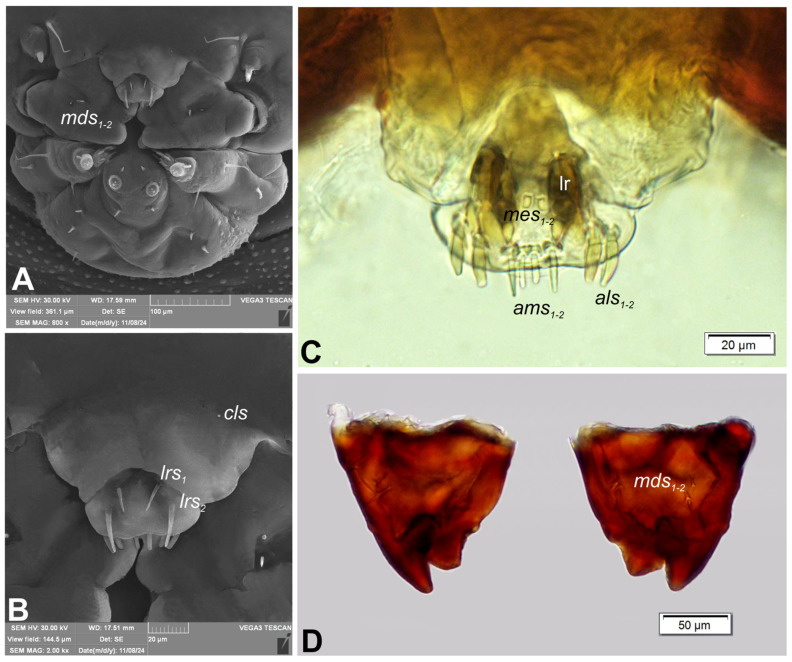 Figure 4
Figure 4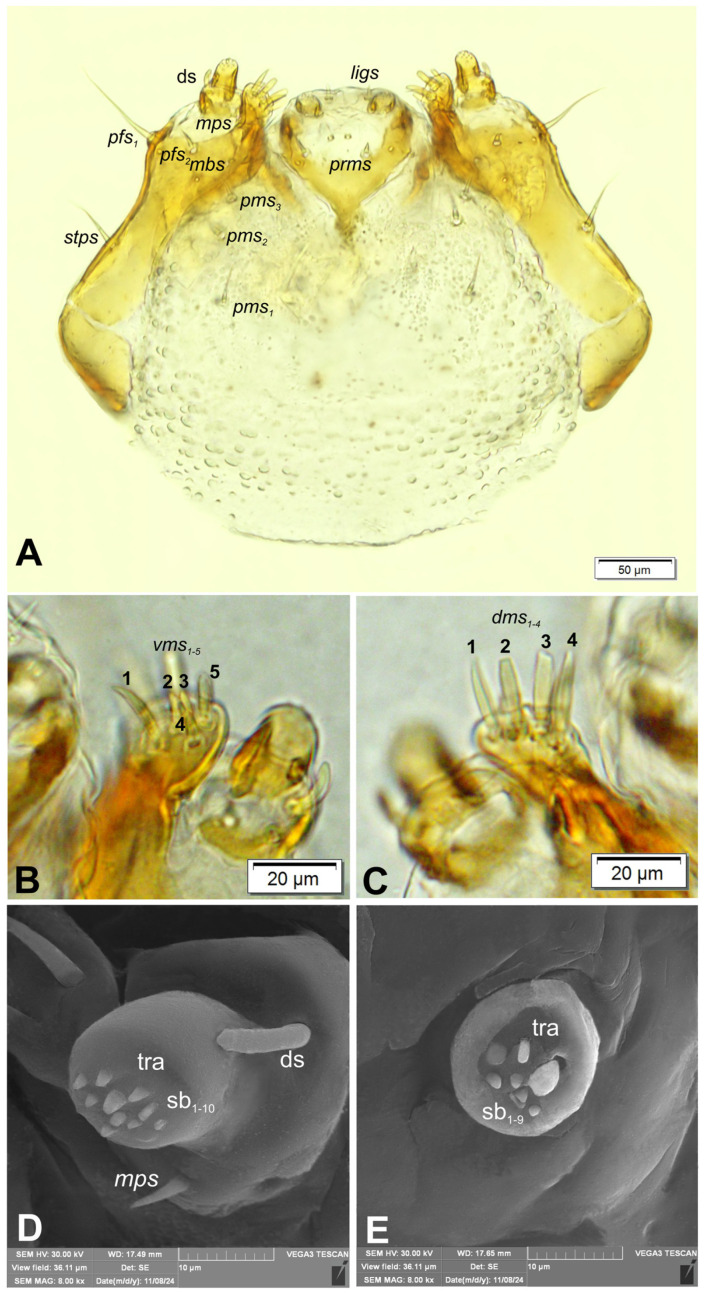 Figure 5
Figure 5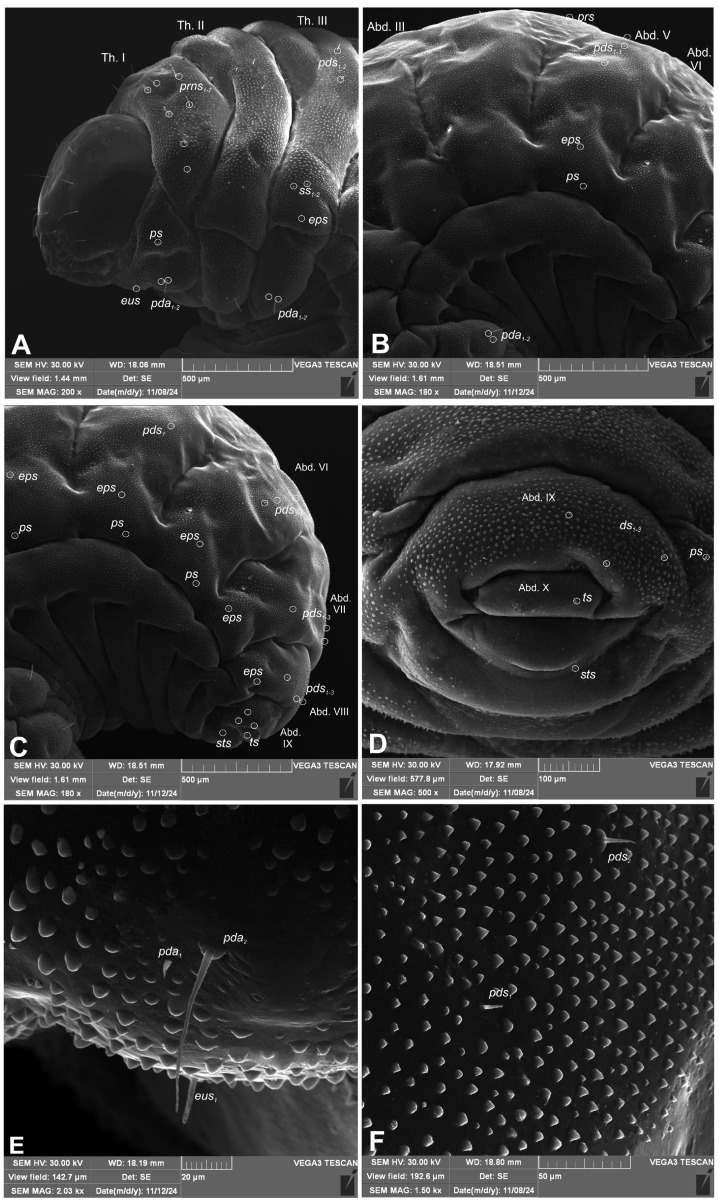 Figure 6
Figure 6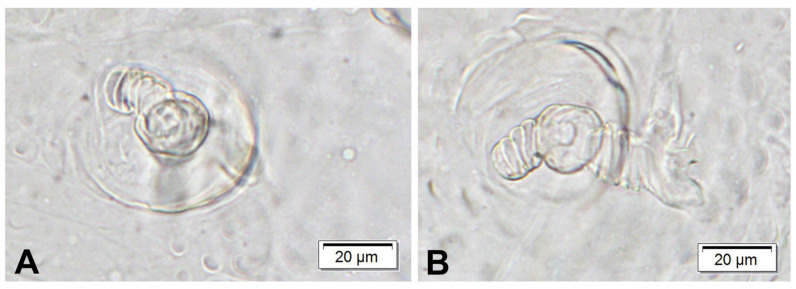 Figure 7
Figure 7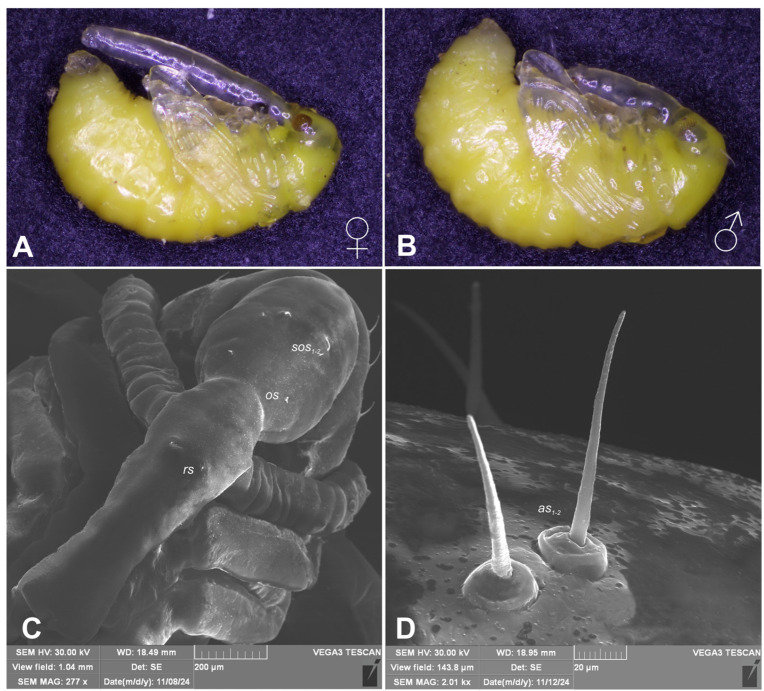 Figure 8
Figure 8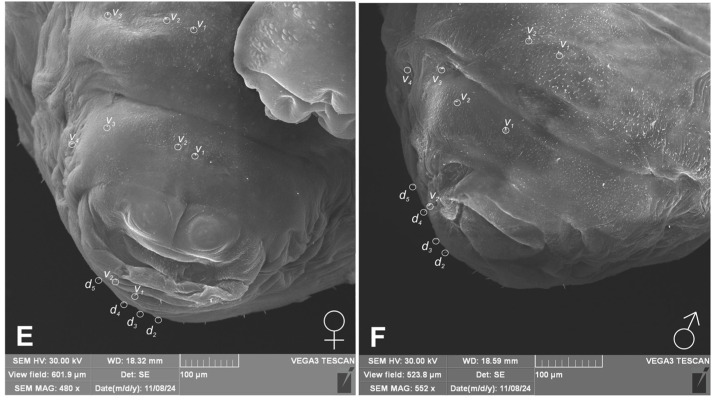 Figure 9
Figure 9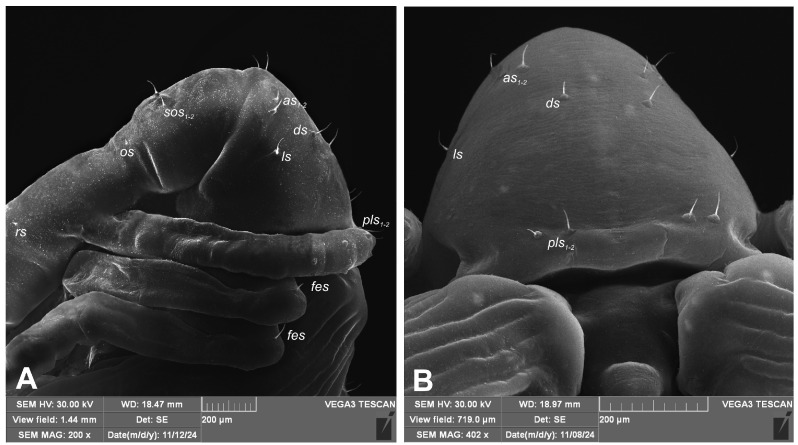 Figure 10
Figure 10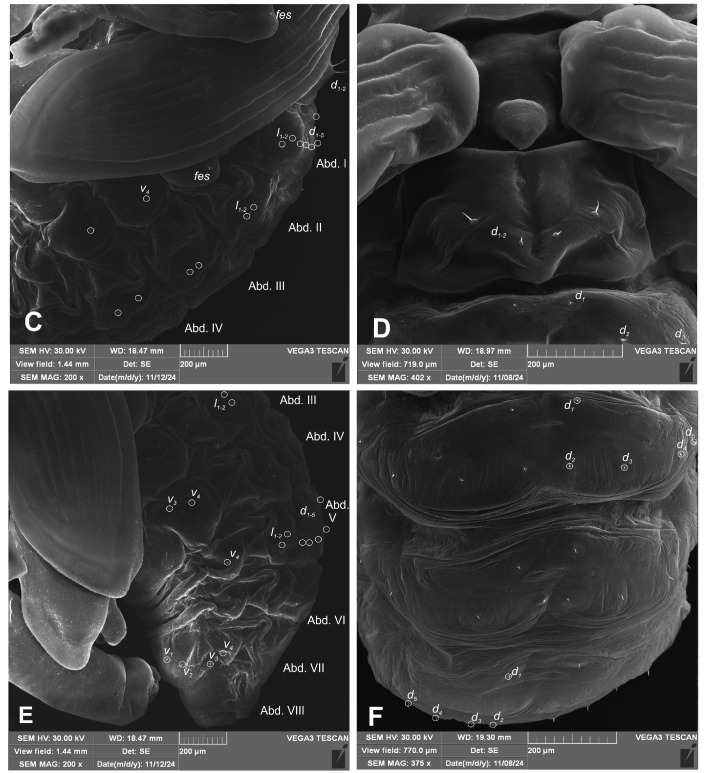 Figure 11
Figure 11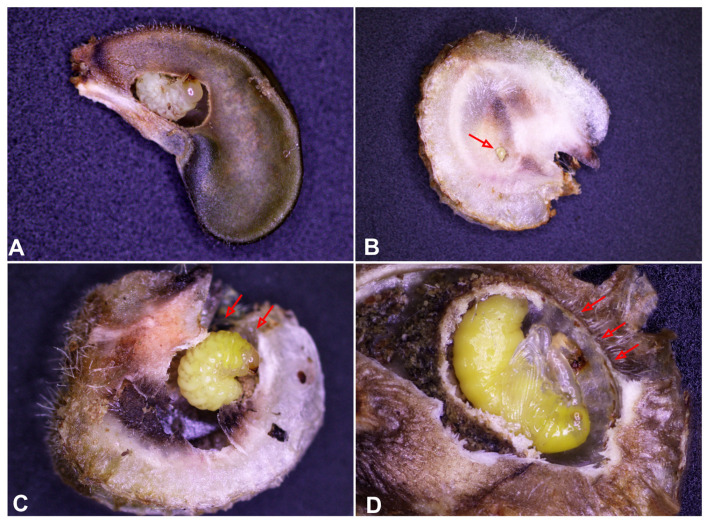 Figure 12
Figure 12 Figure 13
Figure 13Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsForest Insect Ecology and Management · Entomological Studies and Ecology · Insect-Plant Interactions and Control
1. Introduction
The genus Rhopalapion Schilsky, 1906 contains only two species: R. celatum Giusto, 2021a and R. longirostre (Olivier, 1807) [1]. The bilaterally flattened, elongate body, elongate antennal club and elongate rostrum in females make the genus readily distinguishable within the family Brentidae [1,2].
The range of R. celatum is restricted to central Asia. Since the 1960s, R. longirostre has rapidly expanded its range from central Asia to the Middle East and the Mediterranean region, across the whole of Europe (except Scandinavia) to the British Isles. Moreover, R. longirostre was introduced to North America, where it has since spread from East to West. In mountain regions, it has been recorded at altitudes up to 2150 m a.s.l. [1,2,3,4,5,6]. According to Knutelski and Petryszak [7] and Sprick et al. [8], the north-westward expansion of R. longirostre appears to be a response to climate change. At the same time, its patchy distribution, particularly within residential areas, clearly indicates that its expansion is anthropogenic. It is worth mentioning that these beetles, especially the females, are quite efficient, active fliers [9].
Both Rhopalapion species are heliophilic, xerothermophilous weevils inhabiting pastures, meadows, roadsides, ruderal communities, and gardens, especially in Europe. That is why in some countries the weevil is regarded as a pest of ornamental plants [10,11].
Feeding and development in the genus Rhopalapion is closely associated with plant species like Alcea digitata Alef., A. dissecta (Baker f.) Zohary, A. rosea L. and A. setosa (Boiss.) Alef. Current observations do not endorse historical records regarding its occurrence on other genera in Malvaceae or on many other plant families. Moreover, records of R. longirostre on cotton plants (Gossypium sp.) in Turkey and the USA require confirmation [1].
The phenology of Rhopalapion is well known, having been summarized in detail by Giusto [1]. The adults are active (depending on the latitude) from March to November, but under certain conditions, they may be active all year round. Oviposition takes place from May to July, with females laying eggs in holes bored in the flower buds. After 3–4 days, the first instar larva hatches, migrates to the ovary and enters a single pericarp, where it starts to consume the seed, forming a chamber inside the pericarp. Then, it bores an escape hole in the side of the pericarp, which is subsequently sealed with a protective secretion. Pupation takes place inside the pericarp and lasts 2–6 weeks, depending on the weather conditions. The whole developmental period lasts for 7–10 weeks. Because of the species’ prolonged period of activity, different generations may coexist during a single season. Overwintering takes place under plant remains close to the host plant [12,13,14,15,16,17]. Being the subject of over 300 publications [1], R. longirostre has become one of the most frequently chosen models in morphological [18], ecological and zoogeographical studies [19].
The aims of this study were (1) to provide new information on the biology of this species, in particular with regard to the morphology of the preimaginal stages, and (2) to present new observations of its reproductive and developmental behaviour.
2. Materials and Methods
Larvae: 21 exx, L3, 22 July 2024, Lublin, Poland, garden, from hollyhock seeds.
Pupae: 42 exx: (10♀, 10♂), 24 July 2024, Prawiedniki, Poland, side road, from hollyhock seeds; (11♀, 11♂), 31 July 2024, Lublin, Poland, garden, from hollyhock seeds.
The activity, behaviour, and breeding of this species were observed on many hollyhocks growing in gardens, side roads and ruderal plant communities in Lublin (Poland) and Zemun (Serbia) areas during the 2023 and 2024 seasons. The immature stages for the morphological description were collected in Lublin.
Five to ten schizocarps were collected from each of the 20 hollyhock exemplars. Breeding was carried out according to the method developed by Toševski [20]. Ultimately, 73 larvae and 67 pupae were obtained, from which 21 mature larvae and 42 pupae (21 male and 21 female) were measured and described morphologically.
All the laboratory experiments and breeding took place at the Maria Curie-Skłodowska University in Lublin. Before writing the description, all the specimens were fixed in 75% ethanol and examined under an optical stereomicroscope (Olympus SZ 60 and SZ11) with calibrated oculars. The following measurements of the larva were taken: body length (BL), body width (BW) (at the third thoracic segment), head capsule width (HW) and head capsule height (HH, measured from the apex to the epistoma). The pupal measurements included body length (BL), body width (BW) (at the level of the mid-legs), head width (HW) (at the level of the eyes), rostrum length (RL) and pronotum width (PW). Slide preparation basically followed May [21]. The head of the larva selected for microscopic study was cut off and cleared, after which the mouthparts were separated. The remaining part of the body was cleared in 10% potassium hydroxide (KOH), then rinsed in distilled water and dissected. Thereafter, the head, mouthparts and body (thoracic and abdominal segments) were separated and mounted on permanent microscope slides in Faure-Berlese fluid (50 g gum arabic and 45 g chloral hydrate dissolved in 80 g distilled water and 60 cm^3^ glycerol) [22].
The photographs were taken usings an Olympus BX63 microscope and processed with Olympus cellSens Dimension software (version 1.18). The larvae selected for SEM (scanning electron microscope) imaging were first dried in absolute ethanol (99.8%), then rinsed in acetone, treated by CPD (Critical Point Drying) and finally gold-plated. TESCAN Vega 3 SEM was used to examine selected structures.
The general terminology and chaetotaxy follow Anderson [23], May [21], Marvaldi [24,25] and Trnka et al. [26]; the terminology for the antennae follows Zacharuk [27].
3. Results
3.1. Morphology
3.1.1. Description of the Mature Larva
All the measurements are given in Table 1 and Figure 1.
Mean values: male: BL–3.59, BW–1.36, HH–0.57, HW–0.62.
Head capsule (Figure 2A–C) perfectly rounded; endocarina reaches half-length of the frons; frontal sutures distinct along entire length up to antennae; stemmata (st) conspicuous, each placed to the side of the antennae. Setae of head hair–like, various in length: from elongate and medium to minute. Cranial setae: des_1_ short, placed close to sutura coronalis; des_2_ absent, des_3_ elongate, placed above frontal suture; des_4_ elongate, placed laterally; des_5_ elongate, placed anterolaterally. Frontal setae: fs_1_ and fs_2_ absent; fs_3_ minute, placed medially; fs_4_ elongate, placed anteromedially; fs_5_ elongate, placed anterolaterally, close to antenna; both les_1_ and les_2_ of equal length, very short; ventral setae absent; postepicranial area with 6 min pes. Pores present: one between des_3_ and des_4_, one between des_4_ and des_5_, and two between fs_4_ and fs_5_.
Antennae (Figure 3A,B) situated on each side of anterior margin of head; membranous basal segment convex, semi-spherical, bearing conical, elongate sensorium and two basiconica (sb) and single ampullaceum (sa).
Clypeus (Figure 4A,B) approximately 2.6× wider than long, with single, minute cls, placed posterolaterally. Anterior margin of clypeus strongly rounded to inside. Labrum (Figure 4B) approximately 2.5× wider than long, anterior margin slightly sinuate; lrs_1_ medium, placed anteromedially; lrs_2_ elongate, placed anterolaterally; lrs_3_ absent. Epipharynx (Figure 4C) with 2 als, various in length; two digitate ams, equal in size; two digitate mes (first medium, second very short). Labral rods (lr) elongate, well sclerotized, parallel. Mandibles (Figure 4D) symmetrical, each with two apical teeth of unequal height, inner one very robust, outer tooth curved and much higher than inner one. Molar area with small, medially placed, conical protuberance. Setae: mds_1_ short, mds_2_ minute, both placed medially, in elongated depression. Maxillolabial complex: (Figure 5A) stipes with one medium stps, pfs_1_ elongate, pfs_2_ short and single minute mbs; mala with row of four digitate, almost equally sized dms, five vms: first elongate, second and third short, fourth medium, fifth elongate (Figure 5B,C); maxillary palpi two segmented; basal palpomere much wider than distal one; length ratio of basal and distal palpomeres almost 1:1; basal palpomere with minute mps, one pore and one robust digitiform sensillum (ds), distal palpomere (Figure 5D) with group of 10 apical sensilla (all basiconicae) on terminal receptive area (tra); labium with cup-shaped prementum, with one short prms placed medially; ligula concave, with one medium ligs; premental sclerite Y-shaped; postmentum rounded, with three pms of various length: short pms_1_, placed posteriorly, medium pms_2_, placed mediolaterally, and short pms_3_ placed medially (Figure 5A). Labial palpi (Figure 5E) one segmented; each palpus with single pore, and group of nine basiconicae apical sensilla of various size on terminal receptive area; surface of labium densely covered with knobby asperities.
Live larva yellowish, with light yellow head capsule. Except for pronotal shield, whole body densely covered with knobby or conical cuticular processes. Body thick, strongly curved, rounded in cross section. Prothorax distinct, pronotal shield weakly separated, smooth. Mesothorax smaller than metathorax; each divided dorsally into two lobes (prodorsal and postdorsal lobes almost equal in size). Pedal lobes of thoracic segments weakly isolated, smooth. First abdominal segment as big as metathorax, segments II–V of similar size, much bigger than first abdominal segment; segments VI–IX tapering towards posterior end of body. Abdominal segments I–VII divided into two lobes of equal size. Abdominal segments VIII and IX dorsally undivided. Epipleural, laterosternal and eusternal lobes of segments I–VIII conical, well isolated. Abdominal segment X divided into two horizontally, external lobe slightly wider than inner one. Anus situated terminally. Except for pronotum, pedal area and terminal part of the abdomen almost invisible. Only a few thoracic setae, hair-like, moderately elongate, all other setae minute, hardly distinguishable from cuticular processes, sometimes absent. Thorax (Figure 6A): prothorax with seven medium-length, equally sized prns (three placed on premental shield, the others below), one ps and one eus, both short. Meso- and metathorax each with two medium pds, two min ss and one min eps. Pedal areas of thoracic segments each with two pda (first minute, second medium, both placed on isolated area) (Figure 6E). Abdomen (Figure 6B–D): segments I–VIII with one min prs, three pds (first minute, second and third short) (Figure 6F), one min eps and one min ps (segments VII and VIII without ps); segment IX with three short ds, one min ps and one min sts; external lobe of segment X with one min ts seta (Figure 6D). Spiracles (Figure 7A,B) on thorax unicameral, placed laterally between pro- and mesothorax; abdominal spiracles unicameral, placed anterolaterally on segments I–VIII.
3.1.2. Description of Mature Pupa
All the measurements are given in Table 1 and Figure 1.
Mean walues: male: BW–1.09, BL–2.80, RL–0.90, PW–0.67; female: BW–1.19, BL–3.09, RL–1.93, PW–0.76.
Body yellowish, elongate, slightly curved. Cuticle on head, rostrum and thorax smooth, on abdomen irregularly covered with minute, hair–like asperities. Rostrum robust, longer than the whole body in female, reaching metacoxae in male (Figure 8A,B). Pronotum trapezoidal, slightly elongate. Mesonotum narrower than metanotum. Abdominal segments I–VII of equal length, segment VIII semicircular, segment IX terminal. Gonotheca undivided in male, divided in female, with tubercles on subcontiguous lobes. Urogomphi (posterior processes) absent (Figure 8E,F). Spiracles placed laterally on abdominal segments I–VI, functional on segments I–V, vestigial on segment VI. Clubs sparsely covered with conical protuberances.
Chaetotaxy is easily seen only on head and pronotum, hardly visible on rest of body, discernible only under the highest magnification, sometimes absent. Head with one medium-length, one short sos and one minute os. Rostrum with single, minute rs. Pronotum with two as, one ds, one ls and two pls. All pronotal setae medium-length, equally long, on small protuberances (Figure 8B). Dorsal parts of mesothorax without setae; metathorax with one medium-length and one short setae, situated medially. Apex of femora with single, medium-length fes. Abdominal segments I–VIII with five minute setae dorsally (first placed posteromedially, second to fifth placed anteromedially). Each lateral part of abdominal segments I–VIII with two minute setae. Ventral parts of abdominal segments I–VIII with four minute setae (first and second placed medially, third and fourth lateromedially). Abdominal segment IX with two minute setae placed anteromedially (Figure 9A–F).
4. Discussion
R. longirostre is subject to evolutionary pressure promoting large-bodied individuals. In the case of females, the large body size (long snout) allows them to effectively drill a deep hole in the perianth of the corona and then lay the egg near the ovary [18]. Wilhelm et al. [28] state that only males of sufficient size have any chance of mating with large females. Hence, large-bodied individuals are favoured in both sexes (Figure 1). According to Wilhelm et al. [17], R. longirostre and Alcea sp. are excellent examples of the competition between an animal and its host plant, as described by Toju and Sota [29]. On the other hand, both Pupier [14] and Wilhelm et al. [18] note that approximately 15% of teneral adults fail to escape from the pericarp after pupation and ultimately die, there being more females than males among the individuals stuck inside the pupal cradles. Pupier [14] states that the adult escapes from the pericarp by enlarging the exit hole previously bored by the larva. Paradoxically, the smaller the individual, the greater its chances of successfully exiting the pupal cradle (Figure 10A–D). Since the average widths of the pupal pronotum are 0.70 mm (male) and 0.75 mm (female) and are slightly smaller than the diameter of the seed (on average 0.85 mm at its widest point), in practice, only the smallest specimens are able to rotate freely inside the pupal cradle. In females, the long snout additionally impairs the ability to manoeuvre freely. In practice, the effective lowering of the pericarp depends on how large a hole the larva bored and what position the larva took up before pupation. Tenerals have the best chance of leaving the pericarp when the exit opening is located near the abdomen in relation to the position of the pupa. However, if it is situated on the dorsal side of the pupa or near the head, the chances of its enlargement and the subsequent exit of the teneral beetle are small.
Wilhelm et al. [18] give an accurate description of the selection mechanisms among pairs for copulation in R. longirostre. It is worth adding, however, that in the life cycle of R. longirostre, there is quite a long period between the onset of activity (early March) and the period of breeding and oviposition (May). During this time, males can be observed adopting a reverse position upon the female (Figure 11A,B). This allows them to “occupy” her before she is ready to mate and at the same time enables them to actively repel other males without the risk of being pushed away.
The fieldwork yielded no parasitoids or pathogens that eliminated R. longirostre specimens during the preimaginal period. Although Wilhelm et al. [18] mention that larvae and pupae of R. longirostre are attacked by fungi or mites, it seems that their presence is secondary and occurs only after the death of the insects. However, one pathogenic factor limiting the number of R. longirostre is the sympatric development of caterpillars of a moth Pexicopia malvella (Hübner, (1805)) in mallow fruits.
Even though the weevil larvae are not a food resource for the caterpillars (no weevil larvae being eaten by this moth’s caterpillars have been observed), both species compete for the same ecological niche. Whereas one hollyhock seed provides sufficient resources for the development of one weevil, the moth caterpillar eats all the seeds in a schizocarp one after another. The caterpillar gnaws through all the seeds one by one, and on encountering weevil larvae or pupae, it either destroys them mechanically or bites a hole in the pericarp and expels the immature weevils (Figure 11C,D). Then, the exit hole is sealed with thread. In this way, one caterpillar can effectively prevent the development of 30 and more weevils. The hollyhock seed moth begins oviposition in mid-June [30]. Because the weevil starts laying eggs earlier than the moth, the latter has no opportunity to choose unoccupied fruit.
To date, within the subtribe Malvapiina Alonso-Zarazaga, 1990, larvae of only one species other than R. longirostre have been described, i.e., Malvapion malvae (Fabricius, 1775) [31], and no other pupae. Hence, isolating the characteristics of the group is impossible at this stage of research. Additionally, the development of Malvapion malva (Fabricius, 1775) on hollyhock requires confirmation. Unlike R. longirostre, each larva of other Malvapiina empties two to three seeds during its growth. This is probably why they never form populations as large as R. longirostre and are generally considered rare [32,33,34].
Wang et al. [35] first drew attention to the primary asymmetry of the mandibles in the larvae of Pseudoaspidapion botanicum Alonso–Zarazaga and Wang, 2011. Diplapion confluens (Kirby, 1808) also has asymmetrical mandibles, a fact that was omitted in the original description [36].
Chaika and Tomkovich [37] noted that polyphagous larvae have a greater diversity of sensilla than monophagous larvae. The fact that the monophagous larva of R. longirostre is equipped almost exclusively with basiconic sensilla fully endorses this observation.
In addition to several other weevils that develop on hollyhocks, e.g., Aspidapion aeneum (Fabricius, 1775) or Alocentron curvirostre (Gyllenhal, 1833), R. longirostre is another species which is rapidly expanding northwards [38,39]. In some countries, this gives it the status of an alien, invasive or even pest species [40]. The main factors enabling the expansion of R. longirostre towards the north-west is climate change [11,41]. In the case of Poland, this has been confirmed by climatological research. For example, compared to 1961–1990, the average annual air temperature in 2011–2020 increased from 7.5 °C to 9.1 °C, while the average winter temperature increased from –1.9 °C to −0.2 °C [42]. Another factor undoubtedly assisting the spread of this species in Poland is that roadside verges are being mown less often. This allows hollyhocks to grow to their full extent, flower and bear fruit, creating opportunities for these insects to complete their development.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Giusto C. Taxonomic revision of the genus Rhopalapion Schilsky, 1906 (Coleoptera, Apionidae) with description of Rhopalapion celatum n. sp. from the Turanian Region Zootaxa 20214908416510.11646/zootaxa.4908.1.333756632 · doi ↗ · pubmed ↗
- 2Alonso–Zarazaga M.A. Barrios H. Borovec R. Bouchard P. Caldara R. Colonnelli E. Gültekin L. HlaváčP. Korotyaev B. Lyal C.H.C. Cooperative Catalogue of Palaearctic Coleoptera Curculionoidea 2nd ed.Sociedad Entomológica Aragonesa; Monografias electrónicas SEA 2023 Volume 14780 Available online: http://sea-entomologia.org/Me SEA 14_2023.pdf(accessed on 15 December 2024)
- 3Osella G. Primo contributo alla conoscenza dei Curculionidi delle oasi xerotermiche Mem. Mus. Civ. Stor. Nat. Verona 196816213233
- 4Kozłowski M. Knutelski S. First evidence of an occurrence of Rhopalapion longirostre Olivier, 1807 (Coleoptera: Curculionoidea: Apionidae) in Poland Snudebille 20056181184
- 5Szypuła J. Wanat M. Nowe stwierdzenia Rhopalapion longirostre (Olivier, 1807) (Coleoptera: Apionidae) w Polsce Wiad. Entomol.200726128
- 6Ugarte San Vicente I. Salgueira Cerezo F. Nuevos registros de Rhopalapion longirostre (Olivier, 1807) para la Península Ibérica (Coleoptera: Curculionoidea: Apionidae)Heteropterus Rev. Entomol.20088117120
- 7Knutelski S. Petryszak B. Nouvelles données sur la repartition en Europe de Rhopalapion longirostre Olivier, 1807 (Coleoptera: Apionidae)L’Entomologiste 1997535153
- 8Sprick P. Winkelmann H. Behne L. Rhopalapion longirostre (Olivier, 1807) (Coleoptera, Apionidae): Anmerkungen zur Biologie und zur aktuellen Ausbreitung in Deutschland Weevil News 2002918