Genetic Profiling of MC3T3-E1 Cells in Different Media: Implications for In Vitro Screening Development
Makoto Izumiya, Hidehiko Nobuoka, Hono Endo, Rintaro Ueno, Masaki Mimura, Naoto Saito, Hisao Haniu

TL;DR
This study shows how different culture media affect gene expression in osteoblast cells, which could improve in vitro screening for bone formation in clinical applications.
Contribution
The study reveals how culture media influence gene expression and calcification in osteoblasts, offering insights for better in vitro screening methods.
Findings
MC3T3-E1 cells showed distinct gene expression profiles depending on the culture medium.
Primary osteoblasts had minimal gene expression differences between media, except for Alpl.
DMEM-cultured primary osteoblasts showed calcification differences compared to αMEM(−) conditions.
Abstract
Background/Objectives: The translation of in vitro biomaterial evaluations into successful clinical applications often fails due to discrepancies with in vivo results. Previously, we demonstrated that differences in culture medium conditions influence the bone formation process. This study aimed to investigate the influence of culture media on gene expression during calcification induction in osteoblasts. Methods: Using MC3T3-E1 cells cultured in α Minimum Essential Medium without L-ascorbic acid (αMEM(−)) and Dulbecco’s Modified Eagle Medium (DMEM), we screened gene expression profiles through microarray analysis and validated key findings with quantitative PCR. Additionally, we compared these gene expression patterns with those in primary osteoblasts (POBs) cultured under the same medium conditions. Results: The results revealed distinct gene expression profiles in MC3T3-E1 cells…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
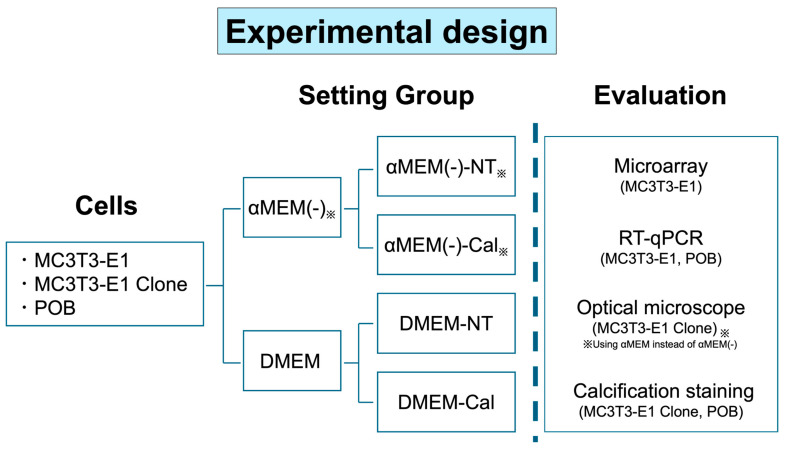 Figure 1
Figure 1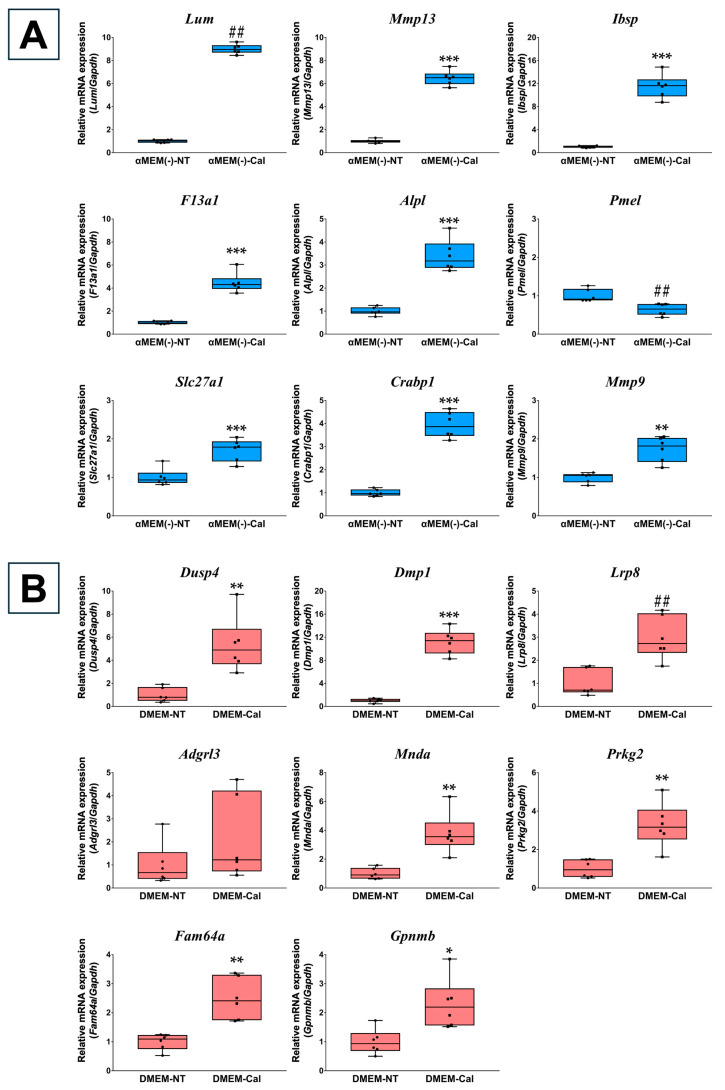 Figure 2
Figure 2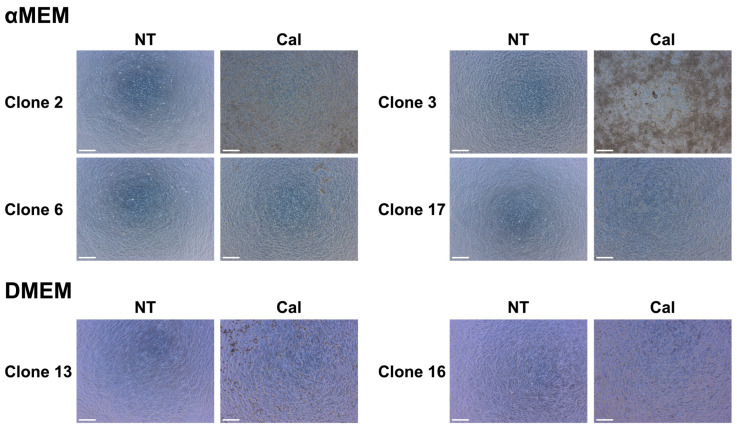 Figure 3
Figure 3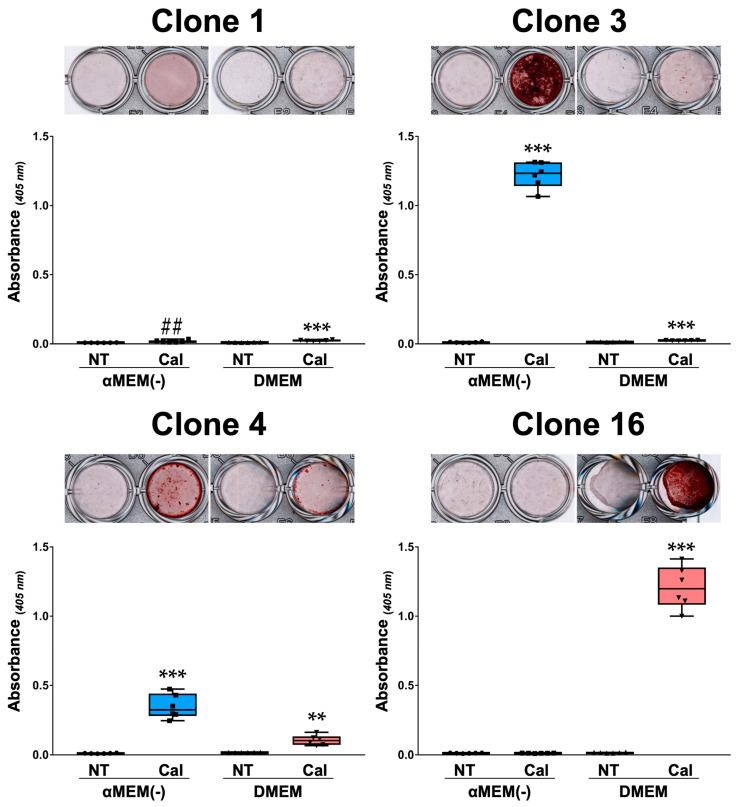 Figure 4
Figure 4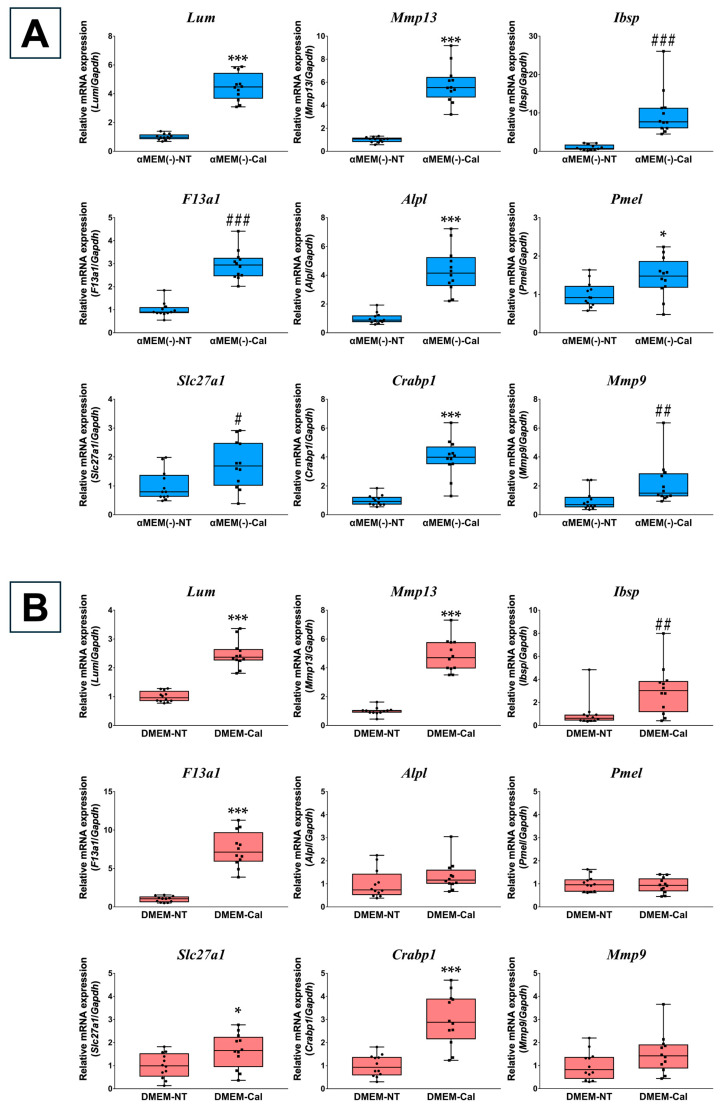 Figure 5
Figure 5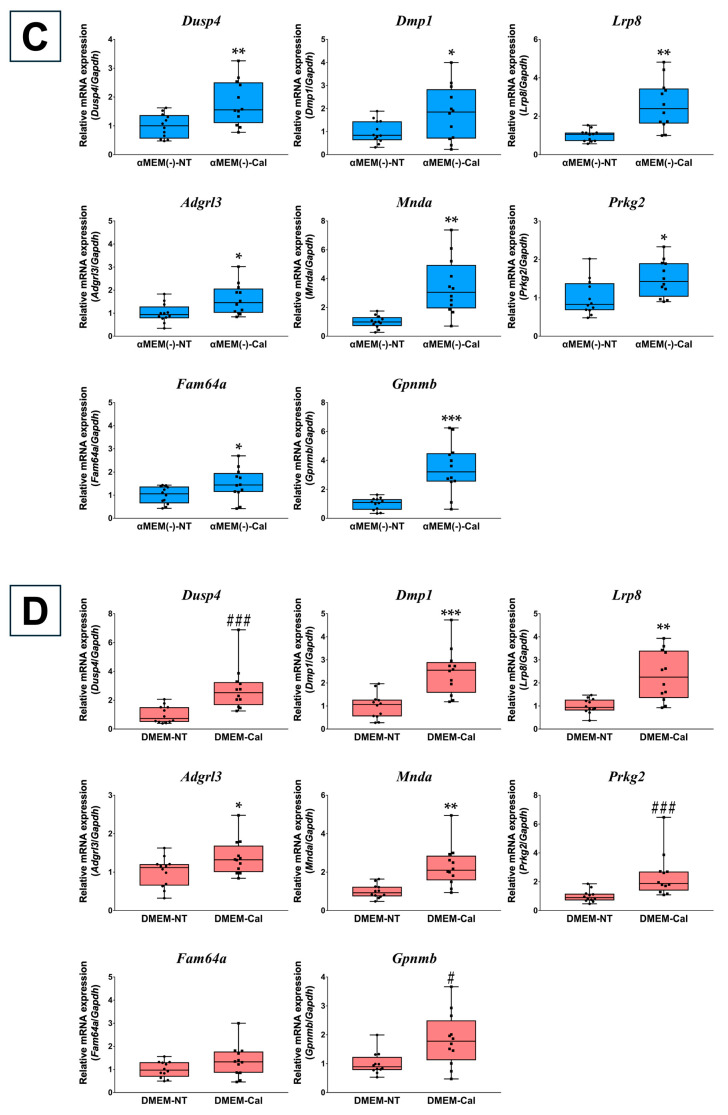 Figure 6
Figure 6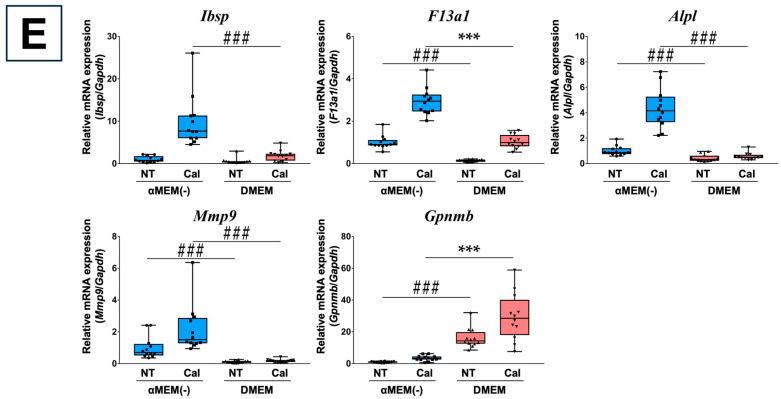 Figure 7
Figure 7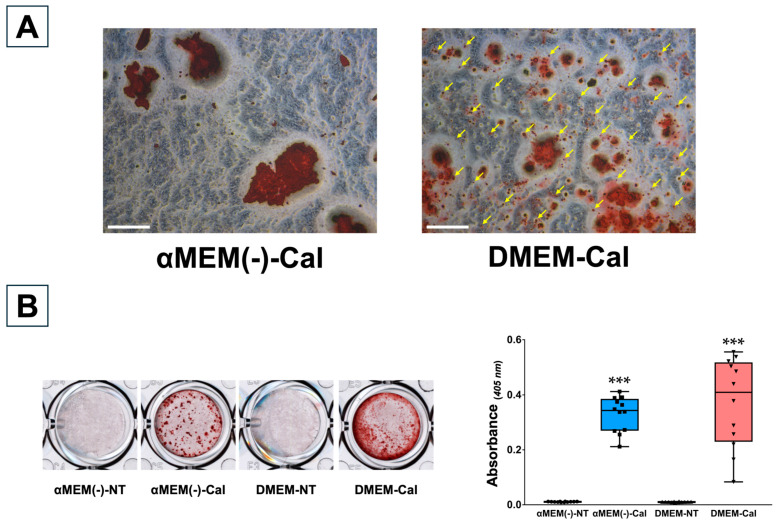 Figure 8
Figure 8- —JSPS KAKENHI
- —JST SPRING
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsNeurogenetic and Muscular Disorders Research · Alkaline Phosphatase Research Studies · Reproductive System and Pregnancy
1. Introduction
The development of new biomaterials and tissue engineering treatments for bone-related diseases relies heavily on animal experiments to evaluate safety and efficacy. However, there is an increasing need to minimize animal use while conducting research efficiently, as mandated by modern scientific standards [1]. To address this, it is essential to establish in vitro evaluation methods that can accurately replicate and screen the diverse bone formation processes occurring in the body.
Since the establishment of culture technology for bone-forming cells [2], functional evaluations have been widely used to assess the biological effectiveness of implant materials on osteoblasts. These evaluations focus on processes such as osteoblast differentiation and bone mineralization and have become the cornerstones of in vitro testing. Despite this, very few new implant materials have been successfully translated into clinical applications based solely on positive in vitro results, such as enhanced osteoblast function. This disconnect is primarily attributed to the frequent discrepancies between in vitro and in vivo functional evaluations [3,4,5], which is thought to be due in part to the complex physiological environment within the body. Currently, no consensus exists on an in vitro bone formation model capable of accurately reproducing the complex physiological environment of bone formation in the body [6]. This remains a significant challenge for researchers and developers.
The MC3T3-E1 preosteoblast cell line is among the most widely used models for studying in vitro bone formation. It has been cited in over 4000 publications, including studies involving its derivatives, highlighting its popularity in the field of bone research [7]. However, MC3T3-E1 cells are known for their instability [8,9]. Wang et al. established a subclone of MC3T3-E1 cells using α Minimum Essential Medium (αMEM) without L-ascorbic acid (αMEM(−)) [9], which differs slightly from the original αMEM formulation used during cell line establishment. This adjustment was based on findings that L-ascorbic acid supports collagen matrix synthesis and stimulates the expression of osteoblast differentiation markers. Despite these considerations, medium conditions are often overlooked in in vitro evaluations of biomaterials [10,11], resulting in significant variability across studies.
Our previous study demonstrated that the freshness and composition of the culture medium significantly influenced osteoblast differentiation-related gene expression, cell proliferation, alkaline phosphatase activity, and mineralization in MC3T3-E1 cells [12]. Specifically, we observed that commercially available liquid αMEM contained inactive L-ascorbic acid, showing no significant differences compared to αMEM(−). However, bone formation processes and mineralized nodule formation induced by MC3T3-E1 cells varied depending on the type of medium used. These findings suggest that multiple in vitro bone formation processes may exist. In contrast, bone formation in vivo is equally diverse, involving distinct pathways such as membranous and endochondral ossification. Additionally, bone formation occurs under various physiological and pathological conditions, including development, remodeling for homeostasis, fracture healing, and ectopic ossification [13,14]. It remains unclear whether in vitro bone formation under different medium conditions mirrors any specific in vivo processes or corresponds to particular states of in vivo bone formation.
Given these challenges, there is an urgent need to understand how culture medium conditions influence in vitro bone formation processes and their relevance to in vivo mechanisms. This study aims to identify gene expression differences in MC3T3-E1 cells under varying culture conditions and to assess their correspondence to primary osteoblast (POB) behavior, with the goal of improving in vitro models for bone formation research.
2. Materials and Methods
2.1. Cells
2.1.1. MC3T3-E1
MC3T3-E1 (cell no. RCB1126) cells were obtained from RIKEN BRC (Ibaraki, Japan). The cells were expanded for two passages in αMEM (Thermo Fisher Scientific, Waltham, MA, USA) supplemented with 10% fetal bovine serum (FBS) (Thermo Fisher Scientific, Waltham, MA, USA), following the recommended culture conditions. The expanded cells were then aliquoted into small portions and stored at −85 °C.
2.1.2. MC3T3-E1 Clone
MC3T3-E1 clone cells were generated from the parental MC3T3-E1 cells using the limiting dilution method. Briefly, the cell concentration of MC3T3-E1 cells was measured using a Trypan Blue solution (Sigma-Aldrich, Saint Louis, MO, USA) with a TC20^TM^ automated cell counter (BIO-RAD, Hercules, CA, USA). In accordance with the published literature [15], the cells were diluted with αMEM (Nacalai Tesque, Kyoto, Japan) supplemented with 10% FBS to 2 cells/mL, and then seeded in 100 µL portions onto 96-well plates (Sarstedt, Numbrecht, Germany). Once single colonies had formed, the cells were expanded, aliquoted into small portions, and stored at −85 °C.
2.1.3. Primary Osteoblasts
Based on the national and institutional regulations and guidelines, all animal experiment procedures were reviewed by the Committee for Animal Experiments and approved by the president of Shinshu University (Approval Number 021078). Primary osteoblasts (POBs) were isolated from the calvaria of C57BL6/N neonatal mouse pups within 1 week of birth, following a modified protocol based on Bakker and Klein-Nulend [16]. The collected calvarial bones were washed, fragmented into 1–2 mm pieces, and digested using a solution containing 0.05 w/v% Trypsin-0.53 mmol/L EDTA 4Na solution (FUJIFILM Wako, Osaka, Japan), and 0.064% collagenase II (Worthington Biochemical, Lakewood, NJ, USA). The reaction was performed at 37 °C for 100 min with intermittent stirring every 10 min. The reaction was halted using Dulbecco’s Phosphate-Buffered Saline (DPBS) containing FBS, and the cell suspension was filtered through a 100 μm cell strainer (Greiner Bio-One, Kremsmuster, Austria). The filtrate was centrifuged at 300× g for 5 min, and the resulting pellet was resuspended in DPBS. The cell suspension was divided equally and cultured in two different media as recommended in the POB harvesting protocol [16,17]: αMEM(−) (Thermo Fisher Scientific, Waltham, MA, USA) supplemented with 10% FBS and Dulbecco’s Modified Eagle Medium (DMEM) (FUJIFILM Wako, Osaka, Japan). The following day, the medium was replaced with fresh media. Cells were passaged upon reaching 80~90% confluency, and cells within two passages were used for experiments. The experimental design is shown in Figure 1.
2.2. DNA Microarray
MC3T3-E1 cells were thawed and cultured in two different media–αMEM(−) and DMEM supplemented with 10% FBS–and then seeded onto 12-well plates (Sarstedt, Numbrecht, Germany) at a density of 6.0 × 10^4^ cells/mL (n = 4). The following day, the cells were divided into two treatment groups: (1) non-treated medium for growth; and (2) calcification-inducing medium, which contained 100 μg/mL of L-ascorbic acid (Nacalai Tesque, Kyoto, Japan) and 5 mM of β-glycerophosphate (Calbiochem, La Jolla, CA, USA). 3 days after induction, total RNA was extracted using a RNA extraction kit (Nippon Genetics, Tokyo, Japan) according to the manufacturer’s protocol. Equal amounts of RNA (5 µL per sample) from four replicates were pooled for each group. DNA microarray analysis was conducted by Macrogen Inc. (Seoul, Republic of Korea) using the Clariom™ S assay for mice (Thermo Fisher Scientific Affymetrix, Santa Clara, CA, USA). In brief, the Affymetrix whole-transcript expression array process was performed according to the manufacturer’s protocol (GeneChip Whole Transcript PLUS Reagent Kit (Thermo Fisher Scientific Affymetrix, Santa Clara, CA, USA)). cDNA was synthesized using the GeneChip WT (whole-transcript) amplification kit according to the manufacturer’s instructions. Sense cDNA was fragmented and biotinylated with TdT (terminal deoxynucleotidyl transferase) using the GeneChip WT terminal labeling kit. Approximately 5.5 μg of labeled DNA target was hybridized to the Affymetrix GeneChip array for 16 h at 45°C. The hybridized array was washed and stained in the GeneChip Fluidics Station 450 and scanned with a GCS3000 scanner (Thermo Fisher Scientific Affymetrix, Santa Clara, CA, USA). The calculation of probe cell intensity data and the generation of analysis files were performed using the Affymetrix^®^ GeneChip Command Console^®^ software ver. 4.0 (Thermo Fisher Scientific Affymetrix, Santa Clara, CA, USA). The analysis files were normalized and analyzed using Analysis Power Tools ver. 2.11.6 (Thermo Fisher Scientific Affymetrix, Santa Clara, CA, USA). After comparing the processed data of each group, the top 10 genes with increased expression on day 3 after calcification-inducing treatment were identified for each medium.
2.3. RT-Quantitative PCR
MC3T3-E1 cells and POBs were cultured in two different media–αMEM(−) and DMEM supplemented with 10% FBS–immediately after thawing or harvesting, respectively. The cells were seeded onto 12-well plates at a density of 6.0 × 10^4^ cells/mL and 8.0 × 10^4^ cells/mL, respectively, and divided into two groups for each medium: (1) non-treated medium (MC3T3-E1: n = 6, POB: n = 12); and (2) calcification-inducing medium (MC3T3-E1: n = 6, POB: n = 12). 3 days post-induction, total RNA was extracted using an RNA extraction kit, followed by cDNA synthesis using ReverTra Ace qPCR RT Master Mix with gDNA Remover (Toyobo, Osaka, Japan). Gene expression analysis was performed using THUNDERBIRD Next SYBR qPCR Mix (Toyobo, Osaka, Japan) on a Step One Plus Real-Time PCR System (Thermo Fisher Scientific, Waltham, MA, USA). The primers for gene expression analysis (purchased from Takara Bio Inc. (Shiga, Japan)) are listed in Table 1. Glyceraldehyde-3-phosphate dehydrogenase (Gapdh) was used as the housekeeping gene, and relative gene expression levels were quantified using the ΔΔCt method.
2.4. Optical Microscopy
MC3T3-E1 clone cells were seeded at a density of 6.0 × 10^4^ cells/mL in two different media, αMEM and DMEM, immediately after thawing. After 1 day, the cells were divided into two groups for each medium: non-treated medium and calcification-inducing medium. Once the calcification induction process began, the medium was replaced every 3 days. On day 21 of calcification induction, the cellular states under each medium condition were observed using a Leica DM IL LED inverted phase contrast microscope (Leica Microsystems, Wetzlar, Germany).
2.5. Calcification Evaluation
MC3T3-E1 clone cells were cultured in αMEM(−) and DMEM and seeded onto 48-well plates (Sarstedt, Numbrecht, Germany) at a density of 1.0 × 10^5^ cells/mL (n = 6). POB cells, cultured in αMEM(−) and DMEM immediately after collection, were seeded onto collagen-coated 48-well plates (IWAKI, Shizuoka, Japan) at a density of 8.0 × 10^4^ cells/mL (n = 12). After 1 day of culture, the medium for both cell types was divided into two groups: non-treated medium and calcification-inducing medium. To account for cell detachment, half of the medium was replaced every 3 days during the calcification process. Calcification in MC3T3-E1 clone cells was evaluated 21 days after induction, while calcification in POBs was assessed using Alizarin Red S staining after 35 days. The evaluation procedure involved washing cells with DPBS, followed by fixation with ice-cold methanol for 20 min. After removing the methanol, cells were washed with pure water and stained with a 1% Alizarin Red S (Sigma-Aldrich, Saint Louis, MO, USA) solution (pH 6.4) for 5 min. Excess stain was removed by washing with pure water, and the plate was left to dry. For quantification, calcified nodules were dissolved in 5% formic acid (FUJIFILM Wako, Osaka, Japan) with agitation for 10 min. The absorbance of the resulting eluate was measured at 405 nm to determine calcification levels with a PlateReader AF2200 (Eppendorf, Hamburg, Germany).
2.6. Statistical Analysis
Statistical analysis was performed using GraphPad Prism Ver. 7.02 (Graph Pad Software Inc., San Diego, CA, USA). The Shapiro–Wilk test was performed to test for normality. Welch’s t-test was applied for comparisons between two groups when normality was established, while the Mann–Whitney U test was used for non-normal data. A 95% confidence interval was assumed, with the significance level set at p < 0.05.
3. Results
3.1. Comprehensive Gene Analysis of MC3T3-E1 Cells Cultured in Different Media After Calcification Stimulation
MC3T3-E1 cells were cultured in αMEM(−) and DMEM, respectively, and their gene expression profiles were compared 3 days after calcification-inducing stimulation using microarray analysis. The results revealed that the top upregulated genes differed markedly between the two culture media. In the αMEM(−) culture, the most upregulated genes, in order, were Lum, Mmp13, Ibsp, F13a1, Alpl, Pmel, Slc27a1, Crabp1, Mmp9, and Gm11099 (a predicted gene) (Table 2). Notably, none of these genes exhibited more than a two-fold upregulation in the DMEM culture.
Conversely, the most upregulated genes in the DMEM culture were Dusp4, Dmp1, Lrp8, Adgrl3, Mnda, Olfr775, Prkg2, Fam64a, Gm13212 (a predicted gene), and Gpnmb, in that order. Among these, only Dmp1 and Gpnmb showed approximately a two-fold increase in the αMEM(−) culture, while the other genes did not exceed a two-fold upregulation.
3.2. Validation of Microarray Data Using RT-qPCR
The microarray results were validated using RT-qPCR for the top upregulated genes, excluding predicted genes, in each medium after 3 days of calcification-inducing stimulation. In MC3T3-E1 cells cultured in αMEM(−), all selected top genes, except Pmel, were significantly upregulated. For Slc27a1 and Mmp9, expression levels were upregulated by 1.71-fold (p = 0.0008) and 1.73-fold (p = 0.0016), respectively, which, while less than two-fold, aligned closely with the microarray results (Figure 2A). In MC3T3-E1 cells cultured in DMEM, all selected top genes, except Adgrl3 and Olfr775, were significantly upregulated. Olfr775 was undetected (cutoff Ct value > 40). These findings were consistent with the microarray results (Figure 2B).
3.3. Optical Microscopy of MC3T3-E1 Clone Cells Cultured in Different Media
Seventeen types of MC3T3-E1 clone cells were cultured in αMEM and DMEM, and their morphological characteristics were observed under an optical microscope 21 days after calcification induction. Significant differences were observed between clones and medium conditions (Figure 3). In the αMEM-calcification induction (Cal) group, Clone 2 exhibited no island-like nodule formation, and brown regions covered a wide area. Clone 3 displayed large black and brown island-like nodules across a wide area, whereas Clone 6 showed black island-like nodules only in some regions. The αMEM-Cal group of Clone 17 was similar to that of Clone 2 but had fewer brown areas overall. In the DMEM-Cal group, Clone 13 showed dark black spherical particles distributed over a wide area, whereas Clone 16 exhibited yellowish-brown spherical particles that were more prevalent and covered a larger area than in Clone 13.
3.4. Calcification of MC3T3-E1 Clone Cells in Different Media
MC3T3-E1 clone cells cultured in αMEM(−) and DMEM were stained with Alizarin Red S on the 21st day after calcification-inducing stimulation to compare the calcification reaction. The calcification response varied by clone and medium type. Representative calcification images of MC3T3-E1 clone cells with notable differences between media are shown alongside the quantification of calcification dissolved in formic acid (Figure 4). In Clone 1, slight staining was observed in both αMEM(−) and DMEM Cal groups compared to their respective non-treated (NT) groups, but no visible calcified nodules were detected. Clone 3 exhibited visible calcified nodules in the αMEM(−)-Cal group (p < 0.0001), with a slightly but significantly higher calcification level in the DMEM-Cal group than in the D-NT group (p < 0.0001). In contrast, in Clone 16, visible calcified nodules were observed in the DMEM-Cal group (p < 0.0001), whereas no significant difference was detected between the αMEM(−)-Cal and αMEM(−)-NT groups (p = 0.7481). The calcified nodule area in Clone 4 was smaller than that of Clones 3 and 16 but was significantly higher in both αMEM(−) and DMEM Cal groups compared to their respective NT groups (p = 0.0002 and p = 0.0017).
3.5. RT-qPCR Verification of Primary Osteoblasts Derived from Calvaria
We performed RT-qPCR to compare the top genes in each medium for POBs derived from calvaria cultured in αMEM(−) and DMEM.
For the top genes upregulated in MC3T3-E1 cells cultured in αMEM(−), POBs cultured in αMEM(−) displayed significant expression increases in all genes after calcification stimulation, showing similar trends as in the MC3T3-E1 cells except for Pmel (Figure 5A). POBs cultured in DMEM showed significant increases in Lum (p < 0.0001), Mmp13 (p < 0.0001), Ibsp (p = 0.0068), F13a1 (p < 0.0001), Slc27a1 (p = 0.0243), and Crabp1 (p < 0.0001); however, Alpl was not upregulated (Figure 5B).
Regarding the top genes upregulated in MC3T3-E1 cells cultured in DMEM, significant increases in expression were confirmed in POBs cultured in αMEM(−) for all genes except Olfr775, which was not detected (Ct value > 40) (Figure 5C). In POBs cultured in DMEM, significant expression increases were observed for all genes except Olfr775 and Fam64a (Figure 5D). While the expression differences were less pronounced in POBs than in MC3T3-E1 cells, POBs exhibited similar trends of increased expression for DMEM-upregulated genes, regardless of the culture medium.
Comparing relative expression levels between αMEM(−) and DMEM, significant differences were observed for several genes (Figure 5E). Ibsp, an osteoblast-related marker, showed a significant difference between the αMEM(−)-Cal and DMEM-Cal groups (p < 0.0001). Alpl expression levels differed significantly between the αMEM(−)-NT and DMEM-NT groups (p = 0.0007), as well as between the αMEM(−)-Cal and DMEM-Cal groups (p < 0.0001). Furthermore, F13a1 and Mmp9 exhibited significant differences between αMEM(−)-NT and DMEM-NT and between αMEM(−)-Cal and DMEM-Cal (p < 0.0001 for all comparisons). Gpnmb (osteoactivin) showed markedly higher expression under DMEM conditions, being approximately 16-fold higher in DMEM-NT than in αMEM(−)-NT and 8.5-fold higher in DMEM-Cal than in αMEM(−)-Cal (p < 0.0001 for each comparison). These results highlight distinct gene expression patterns between αMEM(−) and DMEM.
3.6. Calcification Response of Primary Osteoblasts Derived from Calvaria in Different Media
POBs cultured in αMEM(−) and DMEM were stained with Alizarin Red S on the 35th day after calcification-inducing stimulation to assess the calcification response. Calcified nodules were not observed in any NT group, whereas prominent nodules were formed in the Cal group. Under optical microscopy, POBs cultured in αMEM(−) exhibited localized island-like nodules, whereas POBs cultured in DMEM showed small, widely distributed stained granules, indicating distinct calcification patterns between the media (Figure 6A). Representative calcification images and the quantification of calcified nodules dissolved in formic acid are shown in Figure 6B. Quantitative analysis revealed significantly higher calcification in the Cal group compared to the NT group for both media (p < 0.0001 for each comparison).
4. Discussion
In the MC3T3-E1 experiment, based on our previous report [12], we selected a cell concentration by which the cell proliferation ability due to differences in culture medium would not significantly affect the osteoblast differentiation state. αMEM and DMEM are frequently used for culturing MC3T3-E1, but the main differences between them are the concentrations of amino acids, nucleosides, and vitamins. Orriss et al. have suggested that proline, which is necessary for collagen synthesis and is only found in αMEM, may cause differences in the bone formation activity of POBs [18]. Therefore, we expected that the differences in these medium components would also affect gene expression in the preosteoblast cell line MC3T3-E1 cells. When MC3T3-E1 cells were cultured in αMEM(−) or DMEM and subjected to mineralization stimulation, significant differences in gene expression patterns were observed. Notably, microarray results after mineralization stimulation in DMEM did not show increases in established osteoblast differentiation markers, such as Runx2 and Alpl [19]. Taken together with our previous data, we demonstrated that MC3T3-E1 cells underwent mineralization through the stimulation of the calcification, regardless of the medium type; these findings suggest that osteoblast differentiation markers are not necessarily direct indicators of mineralization. Many studies employ osteoblast differentiation markers in the in vitro evaluation of biomaterials and implants; however, our results indicate that such markers may not adequately assess mineralization potential [20,21]. Instead, the top 10 genes with the most pronounced changes in response to each medium in this study are likely osteoblast response genes activated by L-ascorbic acid stimulation. Supporting this hypothesis, the gene expression analysis after mineralization stimulation of POBs cultured in L-ascorbic acid-free αMEM and DMEM revealed increases in most of the top genes identified in MC3T3-E1 cells, with the exception of Alpl. Interestingly, Alpl expression did not increase following mineralization stimulation in POBs cultured in DMEM, a pattern consistent with MC3T3-E1 cells cultured in DMEM. Alpl, which plays a key role in phosphate supply during bone formation, is considered essential for mineralization [22]. Nevertheless, studies on Alpl knockout mice indicate that while bone formation is abnormal, mineralization is not entirely absent [23,24]. These findings suggest the existence of Alpl-independent pathways of bone formation in vivo. The Alpl unchanged event observed in MC3T3-E1 cells cultured in DMEM, as well as the mineralization of Clone 16 and POBs cultured in DMEM, may reflect these alternative pathways. The specific role and the extent of this Alpl-independent calcification process in physiological bone formation remain unclear and warrant further investigation. However, we propose that the use of osteoblasts cultured in DMEM may enable the study of Alpl-independent calcification mechanisms.
POBs have gained traction as an in vitro model closer to in vivo conditions for evaluating biomaterials and implants [25,26]. Several studies reference Bakker et al.’s protocol in “Bone Research Protocols” for extracting POBs [27,28]. This protocol involves culturing POBs in DMEM supplemented with L-ascorbic acid from the point of extraction. The gene expression profiles observed in this study were derived from POBs cultured under L-ascorbic acid-free conditions and reflect the state of L-ascorbic acid-responsive genes similar to that observed in MC3T3-E1 cells. When evaluating implants using POBs harvested via Bakker et al.’s method, it should be assumed that mineralization stimulation has already occurred. Through this study, we have also confirmed that the calcification stimulation of POBs harvested in DMEM containing L-ascorbic acid yielded no remarkable changes in the expression of numerous genes, including osteogenesis-related genes. This outcome is consistent with previous findings showing that culturing osteoblastic cells in a L-ascorbic acid-containing medium induces the expression of genes known as differentiation-related genes even without additional stimulation [12]. Therefore, the duration of proliferation culture before evaluating biomaterials or implants may significantly influence the results.
The suitability of MC3T3-E1 cells versus POBs for biomaterial evaluation was assessed by comparing gene expression responses to L-ascorbic acid stimulation under varying culture medium conditions. In MC3T3-E1 cells, gene upregulation due to L-ascorbic acid stimulation varied markedly depending on the culture medium. These osteoblastic cells were derived from populations already exposed to L-ascorbic acid during their establishment [2], and their heterogeneity has been well documented [7,9]. This inherent variability likely contributes to the existence of subpopulations, such as Clone 1, that do not respond to additional mineralization stimuli. Conversely, clone experiments revealed that some MC3T3-E1 cells could respond to L-ascorbic acid stimulation, particularly when cultured in a L-ascorbic acid-free medium during proliferation. However, most calcifying clones preferentially responded to specific media, suggesting that the reversibility of calcification stimuli is influenced by the base medium. For instance, in POBs cultured in a L-ascorbic acid-free medium prior to mineralization stimulation, genes related to bone formation, such as Alpl and Gpnmb, exhibited differential expression. The cultivation of POBs in a L-ascorbic acid-free medium requires high cell densities, often necessitating the use of numerous animals, making it impractical for the large-scale screening of biomaterials. In contrast, MC3T3-E1 cells are easily accessible and demonstrate reset gene expression recognized to be related to osteoblast differentiation when cultured in a L-ascorbic acid-free medium. This makes them more practical for biomaterial screening.
Gene expression analysis in animal models suggests that MC3T3-E1 cells cultured in αMEM-based formulations respond to mineralization stimuli in ways that reflect physiological processes such as bone development and fracture healing [13,29]. Conversely, genes such as Dmp1 and Gpnmb, whose expression changed in response to mineralization stimuli in MC3T3-E1 cells cultured in DMEM, have been implicated in bone formation, though their roles in vivo remain poorly understood [30,31]. Interestingly, compensatory bone formation in hypophosphatasia patients is thought to occur independently of Alpl [32], indicating that MC3T3-E1 cells cultured in DMEM might be particularly useful for evaluating biomaterials and therapeutics for such conditions in the field of orthopedics. Further studies are warranted, including histological evaluations focusing on genes extracted with DMEM in animal models of various bone formation situations, as well as in vitro investigations aimed at identifying genes truly related to osteoblast differentiation and calcification to expand this understanding.
One limitation of this study is that it only investigated the difference in status at one time point, 3 days after the calcification stimulus, and so the changes over time were unclear. In addition, no detailed investigation was conducted into the differences in products such as cell matrix that are brought about by variations in gene expression. As such, the direct relationship with calcification is partially unclear. Future research is needed to clarify these points.
5. Conclusions
Comprehensive gene expression analyses in MC3T3-E1 cells cultured in different media revealed distinct top-ranking genes in each case, underscoring the impact of medium conditions on in vitro bone formation processes. The in vitro development evaluation of novel biomaterials for bone disease treatment requires the careful consideration of culture medium conditions, as these significantly influence gene expression during mineralization. In addition, while most of the top genes in MC3T3-E1 cells showed upregulation in POBs cultured in both media, they appear to be responsive to L-ascorbic acid stimulation rather than direct markers of calcification. Variations in Alpl expression, a known osteoblast differentiation marker, and differences in mineralized nodule formation were also observed. It is a very interesting result that there can be an osteoblast state with mineralization ability without increasing Alpl, a known differentiation indicator, and this may correspond to each process of bone formation in the body. In other words, the distinct gene expression profiles in MC3T3-E1 cells may represent multiple bone formation pathways, both in vitro and in vivo. Future investigations on these genes could pave the way for more accurate screening methods for implant evaluation in clinical orthopedics, addressing a critical need in clinical practice.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Stein M. Elefteriou F. Busse B. Fiedler I.A. Kwon R.Y. Farrell E. Ahmad M. Ignatius A. Grover L. Geris L. Why animal experiments are still indispensable in bone research: A statement by the European calcified tissue society J. Bone Miner. Res.2023381045106110.1002/jbmr.486837314012 PMC 10962000 · doi ↗ · pubmed ↗
- 2Kodama H. Amagai Y. Sudo H. Kasai S. Yamamoto S. Establishment of a clonal osteogenic cell line from newborn mouse calvaria Jpn. J. Oral Biol.19812389990110.2330/joralbiosci 1965.23.899 · doi ↗
- 3Mendes S.C. Tibbe J.M. Veenhof M. Both S. Oner F.C. van Blitterswijk C.A. de Bruijn J.D. Relation between in vitro and in vivo osteogenic potential of cultured human bone marrow stromal cells J. Mater. Sci. Mater. Med.2004151123112810.1023/B:JMSM.0000046394.53153.2115516873 · doi ↗ · pubmed ↗
- 4Hatt L.P. Armiento A.R. Mys K. Thompson K. Hildebrand M. Nehrbass D. Müller W.E.G. Zeiter S. Eglin D. Stoddart M.J. Standard in vitro evaluations of engineered bone substitutes are not sufficient to predict in vivo preclinical model outcomes Acta Biomater.202315617718910.1016/j.actbio.2022.08.02135988660 · doi ↗ · pubmed ↗
- 5Mollentze J. Durandt C. Pepper M.S. An in vitro and in vivo comparison of osteogenic differentiation of human mesenchymal stromal/stem cells Stem Cells Int.20212021991936110.1155/2021/991936134539793 PMC 8443361 · doi ↗ · pubmed ↗
- 6Kohli N. Ho S. Brown S.J. Sawadkar P. Sharma V. Snow M. García-Gareta E. Bone remodelling in vitro: Where are we headed?: -A review on the current understanding of physiological bone remodelling and inflammation and the strategies for testing biomaterials in vitro Bone 2018110384610.1016/j.bone.2018.01.01529355746 · doi ↗ · pubmed ↗
- 7Hwang P.W. Horton J.A. Variable osteogenic performance of MC 3T 3-E 1 subclones impacts their utility as models of osteoblast biology Sci. Rep.20199829910.1038/s 41598-019-44575-831165768 PMC 6549152 · doi ↗ · pubmed ↗
- 8Baba T.T. Restoration of mineral depositions by dexamethasone in the matrix of nonmineralizing osteoblastic cells subcloned from MC 3T 3-E 1 cells Calcif. Tissue Int.20006741642110.1007/s 00223000115711136541 · doi ↗ · pubmed ↗