Transcriptome Analysis Reveals the Role of OsCBM1 in Rice Defense Against Xanthomonas oryzae pv.oryzae
Shuaijun Bie, Youlun Xiao, Li Zhang, Yong Liu, Xiaomin He, Jing Peng, Hongjun Xie, Yang Gao, Xiaojuan Li, Xinqiu Tan, Renyan Huang, Deyong Zhang

TL;DR
This study shows that the OsCBM1 gene in rice helps defend against a bacterial infection by boosting stress-related processes.
Contribution
The novel finding is that OsCBM1 enhances rice resistance to bacterial blight via ROS and SA signaling pathways.
Findings
Overexpression of OsCBM1 increases rice resistance to Xanthomonas oryzae pv. oryzae.
OsCBM1 activates the SA signaling pathway and increases antioxidant enzyme activities.
Knockdown of OsCBM1 reduces resistance and reverses the effects of overexpression.
Abstract
Carbohydrate-binding malectin/malectin-like domain-containing proteins (CBMs) represent a newly discovered subclass of lectins that participate in various biological processes across the bacterial, animal, and plant kingdoms. The OsCBM1 gene in rice enhances reactive oxygen species (ROS) burst, contributing to drought-stress tolerance. Nonetheless, the functions of OsCBM1 in response to biotic stress remain poorly understood. In this research, we discovered that OsCBM1 was activated by Xoo infection, and overexpression of OsCBM1 increased rice resistance to bacterial blight, while suppression of its expression shows the opposite trend. OsCBM1 may influence resistance to bacterial blight by regulating ROS burst and the SA signaling pathway through RNA-seq analysis. Overexpression of OsCBM1 increased SA content and enhanced activities of SOD, POD, and CAT enzymes, whereas knockdown of…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
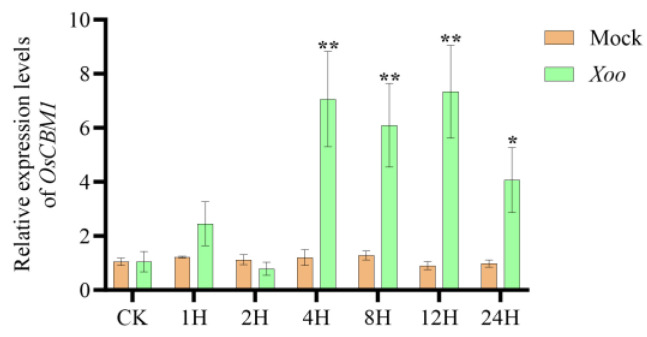 Figure 1
Figure 1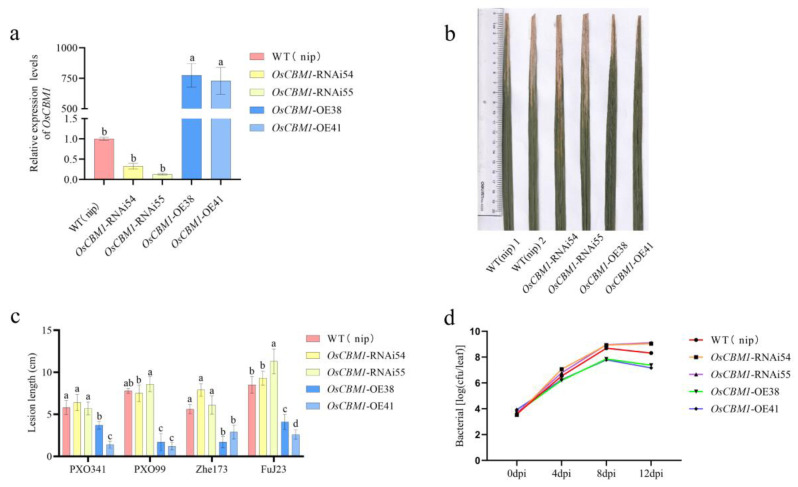 Figure 2
Figure 2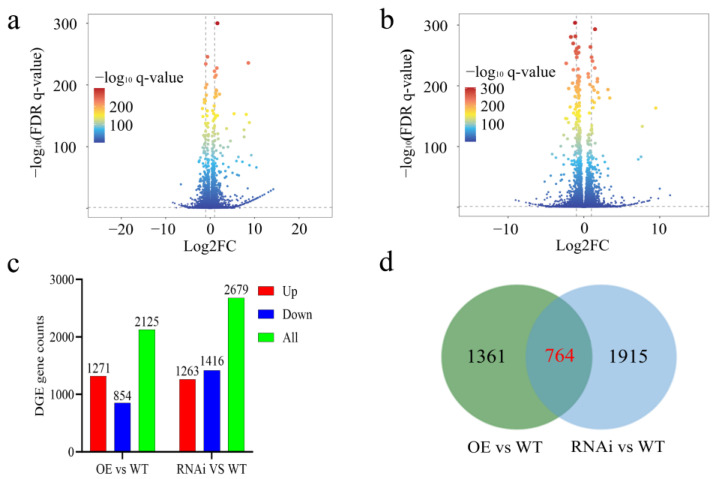 Figure 3
Figure 3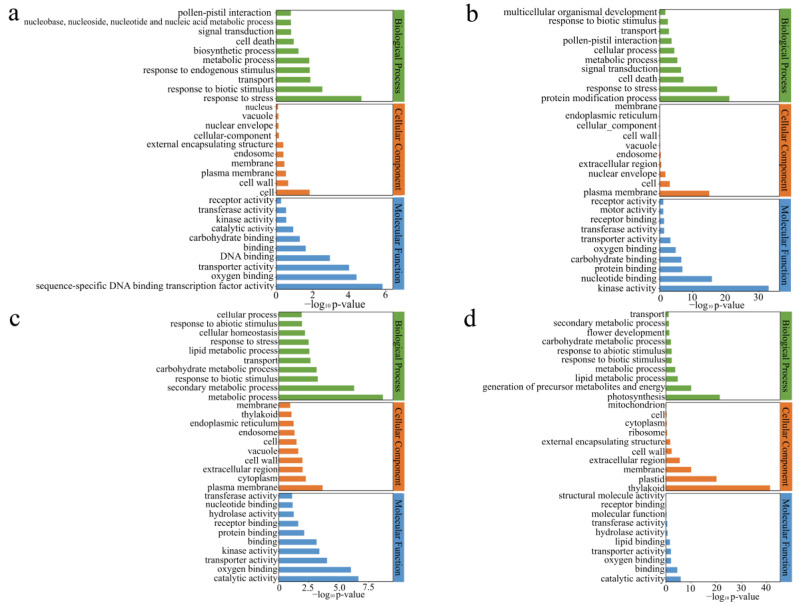 Figure 4
Figure 4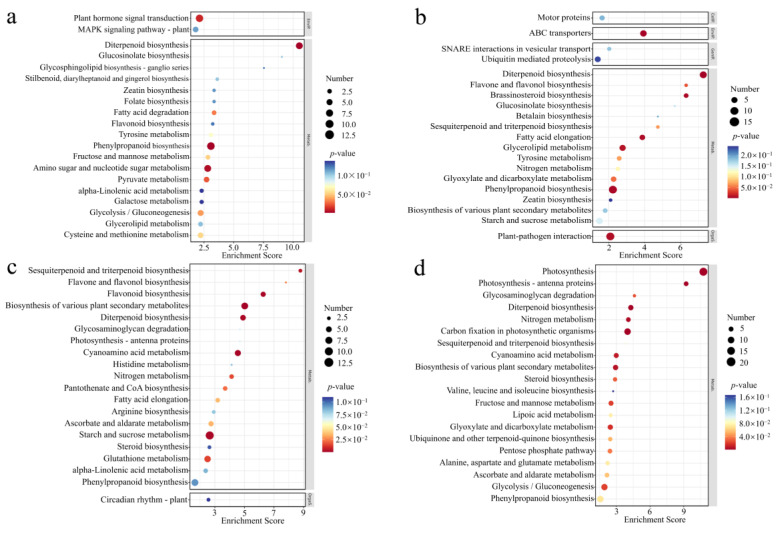 Figure 5
Figure 5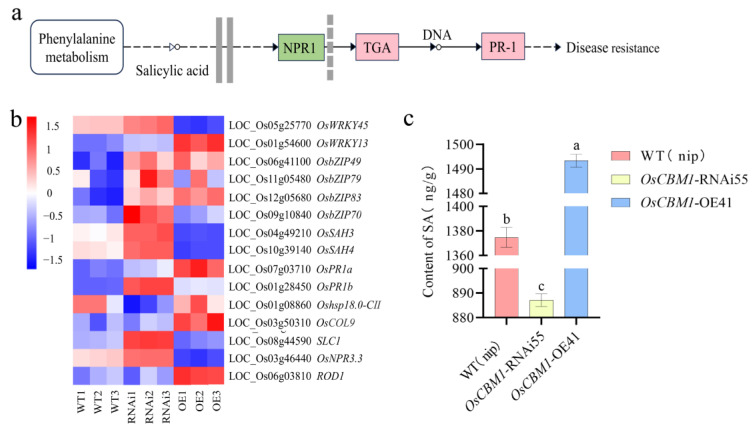 Figure 6
Figure 6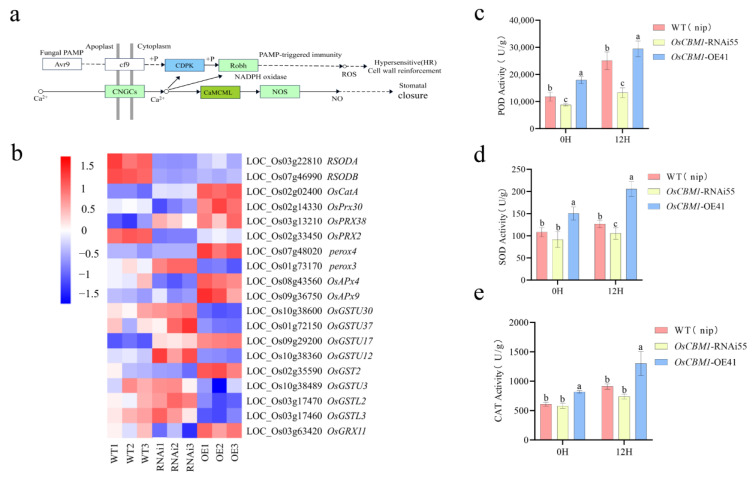 Figure 7
Figure 7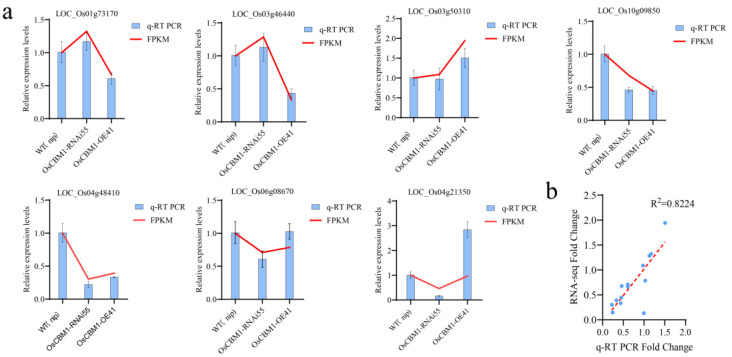 Figure 8
Figure 8- —National Natural Science Foundation of China
- —Hunan Provincial Natural Science Foundation
- —Changsha Municipal Natural Science Foundation
- —Hunan Provincial Agricultural Science and Technology Innovation Fund
- —Hunan Provincial Natural Science Foundation
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsPlant-Microbe Interactions and Immunity · Plant Pathogenic Bacteria Studies · Plant pathogens and resistance mechanisms
1. Introduction
Carbohydrate-binding malectin/malectin-like domain-containing proteins (CBMs), as members of the lectin subfamily, play crucial roles in the biological processes of animals, plants, and microorganisms [1,2]. In plants, CBMs are malectin-like receptor kinases, a newly identified subfamily of receptor-like kinases (RLK). They are also referred to as Catharanthus roseus receptor-like kinase-like proteins (CrRLK1Ls) and play a role in plant immunity, cell wall completion, and other biological functions [3,4,5]. The CrRLK1L family of proteins in plants all possess a malectin-like structural domain, a transmembrane structural domain, and an intracellular serine/threonine kinase structural domain [6]. FERONIA (FER) is the most functional protein in the CrRLK1Ls family, interacting with ROS and Ca^2+^ to mediate pollen tube signaling in the reception and mechanical signaling of Arabidopsis thaliana [7,8]. The CBM1 in rice was discovered to contain just one malectin-like structural domain. and OsCBM1 forms a physical interaction with OsRbohA and OsRacGEF1 to cooperatively regulate ROS production, thereby controlling the sensitivity to the abiotic stress of drought [9]. There are currently no reports on whether the OsCBM1 gene in rice plays a role in regulating the plant’s response to environmental stress.
Rice bacterial blight, triggered by Xanthomonas oryzae pv. oryzae (Xoo), can threaten rice yields [10]. Throughout the process of natural selection, plants have evolved an inherent immune response to combat infestations from pathogenic bacteria. Receptor proteins on plant cell surfaces, known as pattern recognition receptors (PRRs), can detect pathogenic bacteria and activate immunity. This detection process is called pathogen-associated molecular pattern (PAMP) recognition. The immune response triggered by PRRs recognizing PAMPs is associated with what is known as PAMP-triggered immunity (PTI) [11]. In addition, pathogenic bacteria can secrete virulence factors that inhibit plant PTI, and these factors can infiltrate cells and induce plant disease. Plants have coping strategies against pathogens, recognizing specific virulence effectors to trigger a distinct immune response called Effector-Triggered Immunity (ETI), distinct from the Pathogen-Triggered Immunity (PTI) [12]. While PTI serves as the initial defense, reacting to widely conserved pathogen molecules, ETI targets specific pathogen proteins. Despite their differences, PTI and ETI share or intersect in their downstream signaling pathways [13].
Reactive oxygen species (ROS) are important signaling mechanisms that govern plant development and stress response [14,15]. ROS can be produced and accumulated rapidly during plant infection by pathogens [16]. Energy or electron transfer generates reactive oxygen species (ROS); typical examples include hydrogen peroxide (H_2_O_2_), superoxide radical anion (O_2_^−^), hydroxyl radicals (OH^−^), and singlet oxygen (^1^O_2_) [17]. ROS bursts are essential in rice’s response to various pathogen infections; for instance, the rice heavy metal transporter OsNRAMP1 significantly contributes to plant immunity by managing the equilibrium of metal ions and ROS [18]. OsATL32 ubiquitinates the OsRac5-OsRbohB module, which produces reactive oxygen species, thereby inhibiting rice immunity [19]. SPL50, an ARM repeat protein crucial for regulating ROS metabolism and boosting resistance to blast disease [20]. However, excessive levels of ROS in plants can cause damage to plants. To balance the outbreak of ROS, the enzymatic systems containing superoxide dismutase (SOD), catalase (CAT), and peroxidase (POD) play a crucial role. For example, the overexpression of OsANN9 led to heightened activities of SOD, POD, and CAT, which helped to maintain reactive oxygen species homeostasis and improve drought resistance [21]. Similar to the antioxidant system, the phytohormonal signaling pathways play a significant role in enhancing host resistance by mediating downstream signal transduction and safeguarding cells from oxidative damage. Various phytohormones responsive to biotic stress, including salicylic acid (SA), ethylene (ET), jasmonic acid (JA), abscisic acid (ABA), melatonin, and strigolactone, are crucial for maintaining the equilibrium of redox processes within plant cells [14,15,22,23].
Here, we report that overexpression of OsCBM1 enhanced resistance in rice to the bacterial blight, reducing the lesion length ratio by approximately 36.2% to 84.6% against different pathogens. While suppression of this gene exhibited the opposite trend. This result was due to altered SA content and activity of ROS-related enzymes revealed through RNA sequencing and assays of enzyme activity. Our research findings provide a theoretical basis and breeding reference for the involvement of OsCBM1 in rice disease resistance.
2. Materials and Methods
2.1. Plant and Bacterial Material
Rice (Oryza sativa ssp. Geng) Nipponbare (Nip), OsCBM1-RNAi plants, and OsCBM1-OE plants were provided by the State Key Laboratory of Crop Stress Biology in Arid Area, College of Life Sciences, Northwest A&F University [9]. Seedlings were grown in the paddy field of Hunan Academy of Agricultural Sciences in Changsha under natural conditions. The study utilized Xanthomonas oryzae pv.oryzae, including two Philippine strains (PXO99, PXO341) and two Chinese strains (Zhe173 and Fuj23) provided by the National Key Laboratory of Crop Genetic Improvement, National Center of Plant Gene Research, Huazhong Agricultural University. All Xoo strains were cultured at 28 °C on nutrient agar medium.
2.2. Pathogen Inoculation
To assess the response of rice plants to Xoo, the plants were infected with the Xoo strains using the leaf clipping technique during the booting stage (panicle development) [24]. The disease was evaluated by measuring lesion length 14 days after inoculation. Each bacterial inoculation test was performed at least twice. Furthermore, the disease in some plants was assessed by analyzing bacterial proliferation counting colony-forming units as previously described [25]. To assess bacterial growth, a single leaf infected with Xoo from each plant was used as a single replicate, with three plants in total examined for each sample. The bacterial proliferation in rice leaves was assessed by counting colony-forming units [26].
2.3. Measurement of ROS-Related Enzyme Activities
Rice plants (Nipponbare (Nip), OsCBM1-RNAi, and OsCBM1-OE plants) were cultivated in the paddy field until the booting stage, and leaves were collected 12 h after inoculation with the bacterial blight pathogen and quickly frozen using liquid nitrogen. The activities of superoxide dismutase (SOD) and peroxidase (POD) were evaluated using assay kits following the manufacturer’s guidelines (Beijing Solarbio Science and Technology, Beijing, China). The catalase (CAT) activity was determined using assay kits according to the manufacturer’s guidelines (Beijing Boxbio Science and Technology, Beijing, China). All materials were tested in three technical replicates, with WT (Nipponbare) as the control.
2.4. RNA Extraction, cDNA Library Construction, and Sequencing
Rice leaves of Nipponbare (Nip), OsCBM1-RNAi, and OsCBM1-OE plants were collected in liquid nitrogen at 12 h after inoculation with Xoo at the booting stage. All materials were tested in three technical replicates, with WT (Nipponbare) as the control. Total RNA was extracted according to TRIzol reagent (Invitrogen, Carlsbad, CA, USA). The integrity of RNA was evaluated with an Agilent 2100 Bioanalyzer (Agilent Technologies, Santa Clara, CA, USA). RNA purity and quantity were verified using a NanoDrop 2000 spectrophotometer (Thermo Scientific, Waltham, MA, USA). Transcriptome sequencing and analysis were conducted by OE Biotech Co. (Shanghai, China), and transcriptome libraries were constructed using the VAHTS Universal V5 RNA-seq Library Preparation Kit. The libraries were sequenced using the Illumina Novaseq 6000 sequencing platform, and the raw data of this paper can be obtained from the GSE288740 database of NCBI.
2.5. DGEs Analysis and Functional Annotation
DEG analysis was conducted using DESeq2 software (version 1.12.3), with a log2 fold change greater than 1.5 and a false discovery rate (FDR) below 0.05 as the screening criteria to filter out the differential genes among the three groups, and the FDR was obtained by correcting for the significant p-value. Gene ontology (GO) functional annotation and Kyoto Encyclopedia of Genes and Genomes (KEGG) pathway enrichment analysis of the genes were performed using the analysis of the differentially expressed levels of the genes in the samples. FDR was obtained by correcting the significant p-value.
2.6. qRT-PCR Validation
Seven potential genes were chosen for qRT-PCR analysis to ensure confidence in the transcriptome data. Reverse transcription was performed using the 5X RT kit (abm (New York, NY, USA), Cat. No. G592); qRT-PCR was performed using the BlasTagTM 2X qPCR premix (abm, Cat. Nos. G891, G892), and the procedure was carried out as follows: The process began with 3 min at 95 °C, followed by 40 cycles of 95 °C for 15 s and 60 °C for 1 min each, and the OsActin gene was utilized as an internal control in all qRT-PCR. All samples were subjected to three independent replicates, and the results were analyzed using 2^−ΔΔCt^.
2.7. Statistical Analysis
Statistical analyses of enzyme activity and lesion length were performed using independent samples t-test and one-way variance (ANOVA) using SPSS version 27.0 software (SPSS, Chicago, IL, USA); p < 0.05 was considered significant.
3. Results
3.1. The Expression of OsCBM1 Was Triggered in Rice upon Pathogen Xoo Challenge
To explore whether OsCBM1 can effectively protect rice against Xoo infestation, we examined the expression level of OsCBM1 in response to the pathogen Xoo inoculated at 0 h, 1 h, 2 h, 4 h, 8 h, 12 h, and 24 h using a qRT-PCR assay. and it was observed that the expression of OsCBM1 began to increase significantly at 4 h post-inoculation and reached the highest expression at 12 h, with the highest induced expression exceeding 7-fold. (Figure 1). The results indicated that OsCBM1 expression is induced in response to Xoo infestation in rice.
3.2. OsCBM1 Positively Regulated Rice Resistance to Xoo Infestation
To explore the functional role of OsCBM1 in resistance to the disease caused by Xoo, we obtained knockdown lines and overexpression lines OsCBM1-RNAi54, OsCBM1-RNAi55, OsCBM1-OE38, and OsCBM1-OE41 for additional analysis. Expression levels of OsCBM1 were markedly elevated in OsCBM1-OE plants and decreased in OsCBM1-RNAi plants (Figure 2a). We assessed the disease responses of OsCBM1-RNAi and OsCBM1-OE plants to Xoo by inoculation with Xoo during the booting stage. The OsCBM1-OE plants exhibited significantly enhanced resistance to various Xoo strains from the Philippines (PXO341, PXO99) and China (Zhe173, FuJ23), with lesion lengths approximately 2.7 to 6.6 cm shorter than the wild type and reduced Xoo growth rates (Figure 2b–d). However, all the OsCBM1-RNAi plants showed longer or similar lesion length and Xoo growth rates compared to wild type after inoculation with Xoo (Figure 2b–d). Taken together, our findings showed that OsCBM1 was able to trigger plant immunity after Xoo infestation.
3.3. Transcriptome Analysis Identifies Genes Regulated by OsCBM1
To gain a deeper understanding of the molecular mechanism through which OsCBM1-regulated genes enhance Xoo tolerance in rice, we conducted transcriptome analysis of OsCBM1-RNAi plants and OsCBM1-OE plants, as well as wild-type plants, after Xoo inoculation for 12 h. All materials were tested in three technical replicates, with WT (Nipponbare) as the control. Differentially expressed genes (DGEs) were identified (Figure 3a,b). Among the 2125 DEGs, 1217 genes showed increased expression, and 854 genes showed decreased expression between OsCBM1-OE and WT; there were more DGEs, with 1263 genes up-regulated and 1416 genes down-regulated, compared to OsCBM1-RNAi and WT (Figure 3c). Through Venn diagram analysis, it was found that 764 DEGs were differentially expressed in both OsCBM1 overexpression and suppression plants. Additionally, 1361 DEGs were differentially expressed only in OsCBM1 overexpression plants, while 1915 DEGs were differentially expressed in OsCBM1 suppression plants (Figure 3d).
3.4. Analysis of DGEs in GO
To ascertain the potential functions of DEGs, we conducted GO enrichment analysis to investigate the pathways and biological processes involved with OsCBM1. The genes that were up-regulated in OsCBM1-OE vs. WT were most abundantly represented in the biological processes categories of response to stress and response to biotic stimulus. Other highly enriched categories included cell in the category of cell components and sequence-specific DNA binding transcription factor activity, oxygen binding, transporter activity, and DNA binding in the category of molecular function (Figure 4a). However, the GO terms for OsCBM1-RNAi vs. WT indicated that up-regulated genes were predominantly concentrated in protein modification processes, response to stress, plasma membrane, kinase activity, and nucleotide binding were significantly enriched (Figure 4b).
The most abundant terms for OsCBM1-OE vs. WT down-regulated genes were metabolic process and secondary metabolic process in the biological processes categories, and plasma membrane in the cell component category, as well as catalytic activity and oxygen binding in the molecular function category (Figure 4c). Compared with down-regulated genes in the OsCBM1-OE vs. WT, the OsCBM1-RNAi vs. WT had unique GO terms, including photosynthesis, generation of precursor metabolites and energy, thylakoid, and plastid (Figure 4d).
3.5. Analysis of DGEs in KEGG
Comparison of DGEs in OsCBM1-OE vs. WT and OsCBM1-RNAi vs. WT. OsCBM1-OE vs. WT up-regulated genes were primarily concentrated in amino sugar and nucleotide sugar metabolism, phenylpropanoid biosynthesis, and diterpenoid biosynthesis, as well as plant hormone signal transduction (Figure 5a). The pathways enrichment for OsCBM1-RNAi vs. WT up-regulated genes mainly included plant–pathogen interaction, phenylpropanoid biosynthesis and diterpenoid biosynthesis (Figure 5b). OsCBM1-OE vs. WT down-regulated genes were concentrated in several metabolic pathways, including starch and sucrose metabolism, biosynthesis of diverse plant secondary metabolites, phenylpropanoid biosynthesis, and glutathione metabolism (Figure 5c). The most differentially enriched pathways in OsCBM1-RNAi vs. WT down-regulated genes included photosynthesis, carbon fixation in photosynthetic organisms, phenylpropanoid biosynthesis, and glycolysis/gluconeogenesis (Figure 5d). These differences in pathway enrichment may result in altered disease resistance of OsCBM1-OE and OsCBM1-RNAi lines.
3.6. OsCBM1 Regulates the Resistance of Rice to Xoo Through the SA Pathway
To explore the downstream regulatory pathways of OsCBM1 functions, we focused on the genes involved in the phytohormone signaling pathways. The SA-mediated disease resistance pathway has several DGEs affecting related pathways, such as NPR1, TGA transcription factors, and protein-related (PR) genes (Figure 6a). In addition, we screened for fifteen DGEs related to SA synthesis and metabolism, which collectively regulate SA-mediated immune processes (Figure 6b). To further verify whether the OsCBM1 affects Xoo resistance by regulating the content of SA, we determined the levels of SA in rice after Xoo infestation for 12 h in OsCBM1-OE, OsCBM1-RNAi, and WT. It showed that the content of SA was importantly elevated in OsCBM1-OE plants compared to WT, while the opposite result was obtained for OsCBM1-RNAi plants (Figure 6c). These results suggest that the SA signaling pathway is essential for the disease resistance function of OsCBM1.
3.7. OsCBM1 Regulates the Resistance of Rice to Xoo Through ROS Burst
Within the plant–pathogen interaction pathway, ROS is crucial for inducing plant disease resistance. Phosphorylation of Ca^2+^-regulated CDPKs upstream of ROS in the plant–pathogen interactions pathway plays an important role (Figure 7a). In addition, we screened ROS synthesis, metabolic regulation, and related enzyme genes in DEGs, which also included glutathione peroxidase, glutathione sulfurtransferase, and intercellular redox balance (Figure 7b). OsCBM1 participates in ROS production of plants with OsCBM1-OE plants containing higher levels of ROS (H_2_O_2_ and O^2−^) than OsCBM1-RNAi plants [9]. It showed that ROS burst could be crucial in conferring resistance against Xoo mediated by OsCBM1. Elevated ROS levels were also accompanied by elevated activities of related scavenging enzymes. Therefore, we analyzed the concentration of SOD (superoxide dismutase), POD (peroxidase), and CAT (catalase) in leaves of OsCBM1-RNAi plants and OsCBM1-OE plants together with wild type. Both relatively higher SOD, POD, and CAT concentrations in OsCBM1-OE plants than in the wild type, irrespective of whether they are inoculated with Xoo or not, while the trend is reversed in OsCBM1-RNAi plants (Figure 7c–e). These findings indicate that the increasing enzymatic activities of SOD, POD, and CAT could be involved in regulating the ROS burst in OsCBM1-OE plants, resulting in enhanced resistance against Xoo infection.
3.8. qRT-PCR Analysis of Related Genes
To verify the accuracy of RNA-seq, several ROS and SA-related genes were chosen for qRT-PCR analysis. Overall, the gene expression results from the qRT-PCR aligned with the RNA-seq findings, suggesting a comparable pattern between the qRT-PCR and transcriptome data. (Figure 8a). The relationship among the fold change in qRT-PCR and RNA-seq (FPKM) data for the seven differentially expressed genes (DEGs) was analyzed using Pearson correlation and linear regression, revealing a notably positive correlation (Figure 8b).
4. Discussion
4.1. OsCBM1 Enhances Rice Resistance to Bacterial Blight
In a previous study, OsCBM1 was validated to be solely a malectin-like structural domain [9]; CBMs in plants are Catharanthus roseus receptor-like kinase-like proteins (CrRLK1Ls), which are also part of a subclass within the receptor-like kinase (RLK) family. Every protein in this family exhibits a malectin-like structural domain and is involved in cell wall integrity, plant immunity, and other processes [4,6,7]. The transcript of OsCBM1 is primarily expressed in leaves, and the gene is localized to the endoplasmic reticulum and plasma membrane. It can be stimulated for expression by a broad spectrum of biotic hormones and abiotic stress factors. These features suggest that OsCBM1 is a stress-responsive protein [9]. In this study, OsCBM1 positively controls the resistance of rice to bacterial blight. Firstly, OsCBM1 showed increased expression against Xoo infection (Figure 1). Secondly, the OsCBM1-OE plants exhibited markedly enhanced resistance to different Xoo strains from the Philippines (PXO341, PXO99) and China (Zhe173, FuJ23), whereas the OsCBM1-RNAi plants showed significantly reduced resistance (Figure 2b,c). Third, the growth rate of bacterial blight in OsCBM1-OE plants showed a marked reduction compared to the wild type (Figure 2d).
4.2. ROS Bursts Are Essential in the Positive Regulation of Rice Bacterial Blight Resistance by OsCBM1
ROS possess multiple antimicrobial functions in plants, including directly killing pathogens, triggering phytochelatin synthesis, inducing programmed cell death, and activating defense genes [16,27,28]. However, excessive ROS can be toxic to cells, so plants have innately evolved to maintain ROS balance through enzymatic and non-enzymatic scavengers. Enzymatic scavengers like SOD, POD, CAT, APX, and others modulate ROS homeostasis [16]. Here, we believe that OsCBM1 may confer resistance by regulating the burst of ROS. First, the differentially expressed genes from the transcriptome are enriched in ROS-related pathways through KEGG pathway analysis (Figure 4). Some genes in the ROS-related pathways are validated by qRT-PCR, which is consistent with the transcriptomic data (Figure 8). Third, through enzymatic activity assays, it was found that the OsCBM1 gene positively regulates the enzymatic activity changes in SOD, POD, and CAT both before and after inoculation with Xoo (Figure 7). Furthermore, Rbohs are crucial for plant stress tolerance. During rice infestation with M. oryza, OsRbohA and OsRbohB in rice are activated for expression [29]; OsCBM1 directly engages with OsRbohA, enhancing ROS levels and improving drought tolerance in rice [9]. OsRacGEF1 was identified as a ROP nucleotide exclusive to plants exchange factor (PRONE)-type GDP/GTP exchange factor (GEF) family gene. It is activated by chitin at the plasma membrane, engages with OsCERK1, and has its S549 phosphorylated by OsCERK1. Together, they co-regulate resistance to rice blast disease [30]. OsCBM1 physically interacts with OsRacGEF1 to positively regulate ROS production [9]. These results demonstrate that OsCBM1 may promote the burst of ROS and thereby boost rice defense against bacterial blight through interaction with OsRacGEF1 and OsRbohA.
4.3. OsCBM1 May Regulate Rice Resistance to Bacterial Blight by Increasing Salicylic Acid (SA) Content
Salicylic acid (SA) is regarded as a vital hormone for plant defense, enhancing resistance to biotrophic and semibiotrophic pathogens. It is integral to the fundamental defense mechanisms, the intensification of localized immune reactions, and the development of systemic acquired resistance [31]. In this study, we propose that OsCBM1 confers disease resistance because overexpression of this gene increases the content of SA. The reason is that through transcriptomic KEGG enrichment of hormone signaling pathways, variation in the expression of SA-associated genes and changes in SA content were confirmed by qRT-PCR and hormone assays (Figure 4, Figure 7 and Figure 8). SA mediates oxidase reductase activity to regulate wheat immunity against glume blight (Septoria nodorum) [32]. It also promotes resistance against Pseudomonas syringae in Arabidopsis thaliana [33]. Furthermore, SA is involved in regulating colonization of the root by particular bacterial families to mediate plant pathogen resistance [34]. The NPR1 protein plays a crucial role within the SA signaling cascade, and the level of NPR1 expression can influence the expression level of downstream PR-1 [35]. The differential expression of NPR1-related genes in OsCBM1-OE plants in this study may represent a pathway through which OsCBM1 is involved in regulating SA-mediated disease resistance.
4.4. The Immunity of Rice Against Bacterial Blight Potentially Regulated by OsCBM1 Might Also Involve Other Mechanisms
In this study, we found that in the disease-resistant OsCBM1-OE plants, some disease-related genes are expressed, for example, OsPR1a, OsWRKY71, ROD1, OsCYP71Z2, and OsPep3 (Figure S1). Quick activation of the OsPR1a protein is crucial for Xa21-dependent defense against Xoo [36]. OsWRKY71 is crucial for the defense mechanism in rice. Elevating the expression of the OsWRKY71 gene boosts rice resistance to virulent leaf blight by stimulating OsNPR1 and OsPR1b at an earlier stage in the defense signaling pathway [37]. OsPep3 enhances rice’s resistance to Xoo. Hormonal and transcriptomic analyses of OsPep3-treated leaves showed that JA biosynthesis, lipid metabolism, and phenylpropanoid metabolism may contribute to immunity triggered by OsPep3 [38]. OsCYP71Z2 can enhance rice resistance to pathogenic bacteria by regulating the synthesis of plant antitoxins, thereby [39]. Here, there are also some genes that are down-regulated in disease-resistant plants, such as NRR, OsGLIP1, OsBON3, OsCaML2, RH1, RH2, RH3, OsMPK17, OsWRKY76, and others, NRR acts as a negative regulator of disease resistance in rice and interacts with NH1 in rice. Overexpressing NRR plants showed increased sensitivity to Xoo [40]. OsGLIP1 and OsGLIP2 are lipases involved in the hydrolysis of lipids and adversely affect the defense response of rice against bacteria and fungi [41]. OsBON3 promotes rice growth but negatively regulates leaf blight resistance. Pathogen infestation altered the subcellular localization of OsBON1 and OsBON3, which could represent an initial reaction to pathogen invasion [42]. OsCaML2, a target of Osa-miR1422, adversely affects rice resistance to Xoo, and the microRNA Osa-miR1422 improves rice disease resistance by suppressing OsCaML2 [43]. RH1, RH2, and RH3 are three NRR homologous proteins. RH1, RH2, and RH3 can effectively inhibit NH1-mediated transcriptional activation and suppress natural immunity [44]. OsMPK17 negatively regulates Xa21-dependent defense against Xoo in rice [45]. Overexpression of OsWRKY76 reduces defense against leaf blight in rice [46].
5. Conclusions
Our results suggest the possibility that OsCBM1 positively regulates rice defense against Xoo through altering SA levels and enzyme activities of SOD, POD, and CAT. Among them, OsCBM1 is up-regulated following Xoo infection, and its overexpression boosts defense against bacterial blight in rice, whereas knockdown OsCBM1 leads to decreased resistance. OsCBM1 likely affects the plant’s defense by regulating the ROS burst and SA signaling pathways, as indicated by RNA-seq data. Enhanced expression of OsCBM1 raises SA levels and enhances the activity of enzymes like eSOD, POD, and CAT, while reduced OsCBM1 expression results in reduced enzyme activities. Gene expressions linked to SA and enzyme activity were confirmed using qRT-PCR. These results further clarify the function of OsCBM1 in enhancing biotic stress resistance and offer insights for developing disease-resistant rice strains.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Bellande K. Bono J.J. Savelli B. Jamet E. Canut H. Plant lectins and lectin receptor-like kinases: How do they sense the outside?Int. J. Mol. Sci.201718116410.3390/ijms 1806116428561754 PMC 5485988 · doi ↗ · pubmed ↗
- 2Schallus T. Jaeckh C. Fehér K. Palma A.S. Liu Y. Simpson J.C. Mackeen M. Stier G. Gibson T.J. Feizi T. Malectin: A novel carbohydrate-binding protein of the Endoplasmic reticulum and a candidate player in the early steps of protein N-glycosylation Mol. Biol. Cell 2008193404341410.1091/mbc.e 08-04-035418524852 PMC 2488313 · doi ↗ · pubmed ↗
- 3Boisson-Dernier A. Kessler S.A. Grossniklaus U. The walls have ears: The role of plant Cr RLK 1Ls in sensing and transducing extracellular signals J. Exp. Bot.2011621581159110.1093/jxb/erq 44521252257 · doi ↗ · pubmed ↗
- 4Galindo-Trigo S. Gray J.E. Smith L.M. Conserved roles of Cr RLK 1L receptor-ike kinases in cell expansion and reproduction from algae to angiosperms Front. Plant Sci.20167126910.3389/fpls.2016.0126927621737 PMC 5002434 · doi ↗ · pubmed ↗
- 5Nissen K.S. Willats W.G.T. Malinovsky F.G. Understanding Cr RLK 1L function: Cell walls and growth control Trends Plant Sci.20162151652710.1016/j.tplants.2015.12.00426778775 · doi ↗ · pubmed ↗
- 6Franck C.M. Westermann J. Boisson-Dernier A. Plant malectin-like receptor kinases: From cell wall integrity to immunity and beyond Annu. Rev. Plant Biol.20186930132810.1146/annurev-arplant-042817-04055729539271 · doi ↗ · pubmed ↗
- 7Feng W. Kita D. Peaucelle A. Cartwright H.N. Doan V. Duan Q. Liu M.-C. Maman J. Steinhorst L. Schmitz-Thom I. The FERONIA receptor kinase maintains cell-wall integrity during salt stress through Ca 2+ signaling Curr. Biol.20182866667510.1016/j.cub.2018.01.02329456142 PMC 5894116 · doi ↗ · pubmed ↗
- 8Kessler S.A. Lindner H. Jones D.S. Grossniklaus U. Functional analysis of related Cr RLK 1L receptor-like kinases in pollen tube reception EMBO Rep.20151610711510.15252/embr.20143880125490905 PMC 4304734 · doi ↗ · pubmed ↗