Structure of the T=13 capsid of infectious pancreatic necrosis virus (IPNV)—a salmonid birnavirus
Anna Munke, Amr Ahmed Abdelrahim Gamil, Aase B. Mikalsen, Han Wang, Øystein Evensen, Kenta Okamoto

TL;DR
This study reveals the structure of the infectious form of the IPNV virus, which infects salmonid fish, providing insights into its replication and assembly.
Contribution
The first high-resolution structure of the infectious T=13 capsid of IPNV is determined, revealing functional motifs critical for replication and assembly.
Findings
The IPNV T=13 capsid structure was resolved at 2.75 Å, showing similarities to IBDV but with distinct surface loops on VP2.
New structural features include interlocking C-terminal regions of VP2 subunits in the pentagonal assembly unit.
Positively charged residues in capsid pores are speculated to aid intraparticle genome synthesis.
Abstract
Birnaviruses infect a broad range of vertebrate hosts, including fish and birds, and cause substantial economic losses in the fishery and livestock industries. The infectious pancreatic necrosis virus (IPNV), an aquabirnavirus, specifically infects salmonids. While structures on T=1 subviral particles of the birnaviruses, including IPNV, have been studied, structural insights into the infectious T=13 particles have been limited to the infectious bursal disease virus (IBDV), an avibirnavirus. Determining the capsid structure of the T=13 particle of IPNV is crucial for advancing knowledge of its antigenicity, capsid assembly, and possible functional structures. Here, the capsid structure of the IPNV L5 strain has been determined at a resolution of 2.75 Å. The overall structure resembles the T=13 IBDV structure, with notable differences in the surface loops on the P domain of the VP2…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
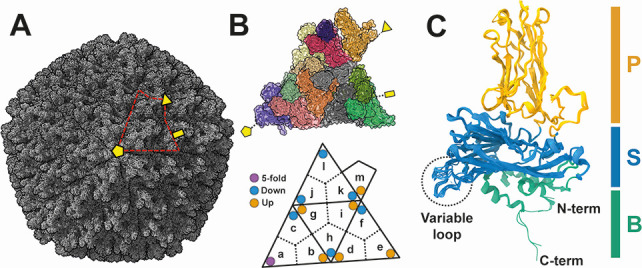 Fig 1
Fig 1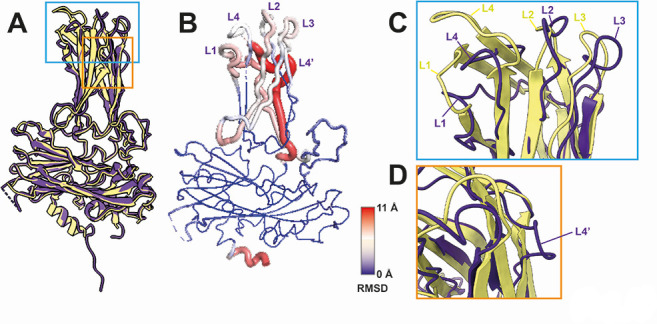 Fig 2
Fig 2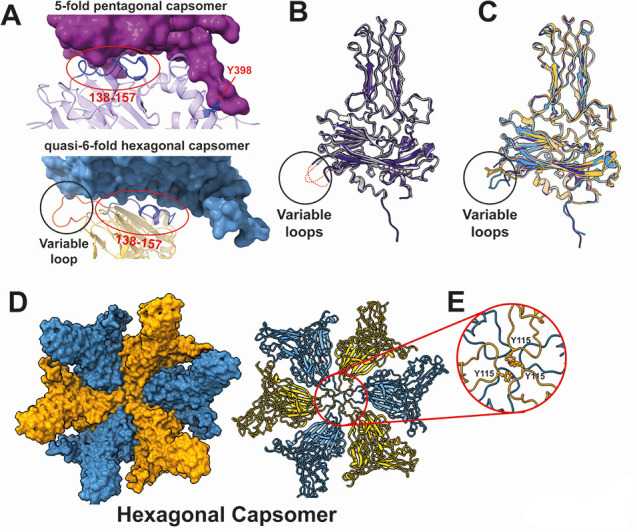 Fig 3
Fig 3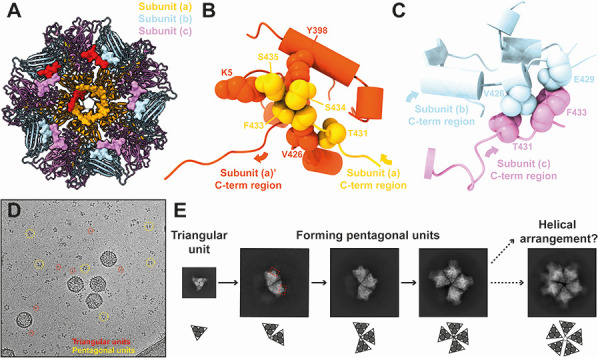 Fig 4
Fig 4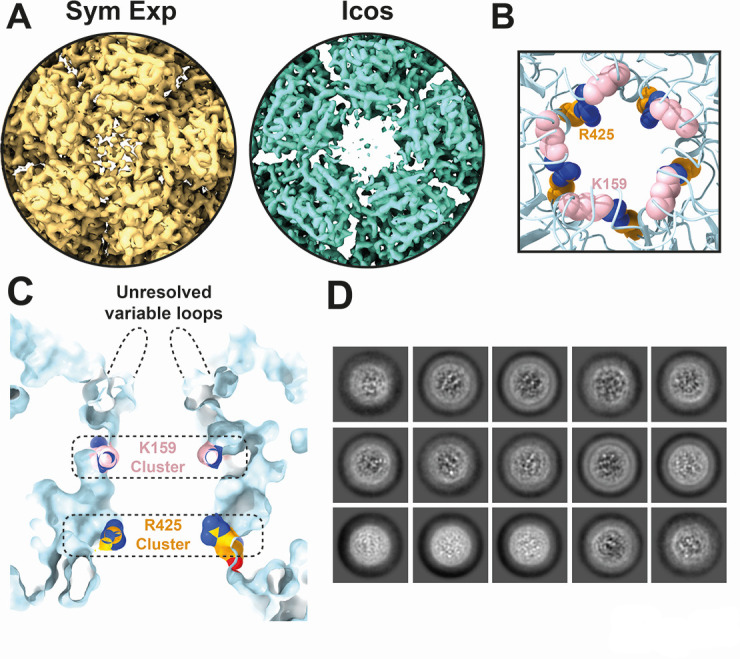 Fig 5
Fig 5- —Vetenskapsrådet (VR)
- —Vetenskapsrådet (VR)
- —Swedish Foundation for International Cooperation in Research and Higher Education (STINT)
- —Norges Forskningsråd (Forskningsrådet)
- —Royal Swedish Academy of Sciences (KVA)
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsAnimal Virus Infections Studies · Viral gastroenteritis research and epidemiology · Virus-based gene therapy research
INTRODUCTION
Birnaviridae viruses are disease-causative agents to vertebrates, encompassing a few species in the genus Aquabirnavirus infecting fishes and genus Avibirnavirus infecting birds. Recently, putative birnaviruses have also been identified in pigs (1). The infectious pancreatic necrosis virus (IPNV), a member of the Aquabirnavirus, can infect diverse salmonid species, leading to lethal outcomes characterized by pancreatic necrosis and catarrhal enteritis (2, 3). Because of its negative impact on farmed fish, control measures, such as vaccination and genetic selection, have been implemented to combat this virus (4, 5). Another important birnavirus pathogen, the infectious bursal disease virus (IBDV), infects chickens and turkeys, leading to severe immunosuppression (5, 6).
Birnaviruses have a bi-segmented dsRNA genome (7, 8). Segment A encodes a large polyprotein consisting of pVP2-VP3-VP4, and, in some species, a short VP5 protein (9, 10). Segment B encodes the RNA-dependent RNA polymerase (RdRp), VP1 (9). The preVP2–VP3–VP4 polyprotein undergoes self-cleavage by the VP4 protease, generating preVP2 and VP3 (11). During particle assembly, preVP2 is further processed to mature VP2 and short peptides by cleaving its C-terminal region (12). The disintegrated birnavirus particle releases bi-segmented dsRNA filaments, which provide the evidence of the formation of ribonucleoprotein complexes (RNPs) (13, 14). In these RNPs, hundreds of VP3s are bound to the viral dsRNA filaments, likely contributing to the compaction of the viral genomes (14). VP1 exists either in a free form or covalently linked to each segmented genome (15, 16). VP3 interacts with VP1 to package the segmented genomes inside the particle (17–19). On the contrary, VP2s assemble into a 60–70 nm T=13 icosahedral single-layer capsid to protect the RNPs (20, 21). The precursor N- and C-terminal regions of VP2 and the C-terminal peptide of VP3 are critical for the formation of infectious large T=13 particles since expressing only the matured form of VP2 results in the formation of non-infectious small ~23 nm T=1 subviral particles (21–26). Short peptide fragments from the cleaved C-terminal region of the preVP2 are present in IPNV particles and also play a critical role in assembling T=13 virus particles (27, 28).
The crystal structures of T=1 subviral particles from both IBDV and IPNV have been determined, revealing three structural domains within VP2: the P, S, and B domains (21, 23, 24, 29). The S domain employs a typical jelly-roll fold of icosahedral viruses, which implies an evolutionary link between the +ssRNA nodaviruses and tetraviruses and the dsRNA birnaviruses and reoviruses (21, 30). The P domain unique to birnaviruses is considerably variable in amino acid sequence and forms surface protrusions (21, 24). Structural variations in the surface loops of the P domain between the IBDV and IPNV subviral particles likely play a role in regulating virulence and tropism (24). Certainly, mutations in the P domain often result in less virulent strains (31–34). The B domain located inside the virus capsid is believed to play a role in genome encapsulation through its arrangement of α-helices, as described in other ssDNA and +ssRNA viruses that employ the jelly roll fold (30, 35–37). The T=1 icosahedron contains only one VP2 subunit in the asymmetric unit, and the structure of this VP2 has been resolved in a single conformation (24), limiting the understanding of the functional structures of the infectious T=13 capsid. This limitation also hinders the comprehension of possible mechanisms of the T=13 particle assembly.
Another uncertain aspect of the birnavirus capsid structure is the surface pore at each 5-fold axis. This pore possibly has a role in facilitating the synthesis of virus transcripts inside the capsid by incorporating nucleoside triphosphates (NTPs) and releasing +ssRNA virus transcripts. This process, known as intraparticle genome transcription, occurs in other icosahedral dsRNA viruses, such as reoviruses and totiviruses (38–46). However, in birnaviruses, the surface pore might be obstructed by unresolved surface loops of VP2 at the 5-fold axis (20, 21). Previous studies have described the capability of purified birnavirus particles to perform intraparticle genome synthesis (47, 48). In contrast, it has also been suggested that the interior RNPs could be released from the capsid during the endocytosis and synthesize viral transcripts in cellular virus factories (49–51). Whether the RNP functions as a capsid-independent transcription complex in situ remains a pending question.
Considering the importance and unrevealed multifunctionality of the birnavirus capsid, it is critical to determine its infectious large T=13 particles using cryogenic electron microscopy (cryo-EM) single particle analysis. To date, only the cryo-EM structure of IBDV has been resolved (20). In this study, we present the first cryo-EM structure of the salmonid IPNV, providing new insights into the structural mechanisms of VP2 that are essential for infection, particle formation, and intraparticle genome synthesis.
RESULTS AND DISCUSSION
Cryo-EM structure of the T=13 IPNV capsid
The first infectious IPNV capsid structure was determined at a resolution of 2.75 Å using cryo-EM single particle analysis with imposed icosahedral symmetry (Fig. S1 and S3). The achieved resolution enabled the building of an atomic model of the IPNV capsid (Fig. S2 and S3).
The IPNV capsid employs a T=13 laevo icosahedral lattice consisting of 60 copies of the asymmetric unit (highlighted with red dotted line in Fig. 1A). Each asymmetric unit is composed of 13 VP2 subunits [labeled (a)–(m) in Fig. 1B]. These VP2 subunits form trimers, such as the one composed of subunits (a), (b), and (c) (Fig. 1B), which may serve as building blocks of the birnavirus capsid. From another perspective, five subunits (a) form a pentagonal unit at the 5-fold axis, while the remaining subunits contribute to hexagonal units in the capsid (Fig. 1B). The atomic models of the 13 VP2s were built within the asymmetric unit (Fig. 1B) and subsequently superimposed to observe their structural diversity (Fig. 1C). The VP2 subunits are organized into three domains: the α-helix-rich N and C-terminal base (B) domain located on the capsid interior; the conserved β-barrel jelly roll fold shell (S) domain; and the unique β-barrel jelly roll fold projection (P) domain, which is situated on the capsid exterior (Fig. 1C). The P domains of the trimer unit form unique protrusions on the capsid surface. While the overall structure of the VP2 subunits is similar, notable structural variations are observed in the C-terminal region of the B domain (residues 429–435) (Fig. S4) and the variable loop region of the S domain (residues 110–117) (5-fold, down, and up conformations) (Fig. 1B and C). The amino acid sequences are well conserved in these regions among birnaviruses’ VP2 (Fig. S5), suggesting their critical roles.
Capsid structure of the T=13 infectious IPNV. (A) Overall cryo-EM structure of IPNV. The asymmetric unit is encircled by a red dotted line. (B) Subunits (a)–(m) within the T=13 asymmetric unit each depicted in a different color. The positions of “5-fold,” “Down,” and “Up” conformations of the variable loops are indicated by purple, light blue, and orange circles, respectively. (C) Atomic structure of the VP2 subunits, with all 13 VP2 subunits superimposed. The P, S, and B domains are colored orange, light blue, and light green, respectively. Variable loops are shown in a black dotted circle. The 2-, 3-, and 5-fold axes are shown by a yellow rectangle, triangle, and pentagon in panels A and B.
Structural comparison between IPNV, IBDV, and other related viruses
The VP2 structure of IPNV shows the greatest similarity to that of IBDV, a member of the same Birnaviridae family (DALI score = 24.6) (Table S1). However, the surface loops of the P-domain are structurally different between the T=1 subviral particles of IPNV and IBDV, demonstrating their involvement in host-specific infection (24). A comparison between the cryo-EM structures of the infectious T=13 IBDV and IPNV particles similarly reveals differences in these surface loops (Fig. 2A and B), reinforcing earlier observations made from the T=1 structures. Both IPNV and IBDV form four corresponding surface loops: L1 (IPNV: 206–222, IBDV: 204–222), L2 (IPNV: 245–260, IBDV: 245–260), L3 (IPNV: 277–287, IBDV: 277–284), and L4 (IPNV: 313–323, IBDV: 313–322), while an additional loop, L4′ (326-334), is unique to IPNV (Fig. 2C and D). These surface loops represent the most structurally divergent regions, especially the IPNV L4′ region (Fig. 2B through D). The IBDV VP2 has a corresponding region of the IPNV L4′ (Fig. S5); however, it displays a more ordered structure. These updated cryo-EM structures of the infectious IPNV and IBDV surfaces reveal more accurate and native insights into the structural differences that may influence host tropism and virulence.
Structural comparisons of VP2 in T=13 IPNV and IBDV particles. (A) Superimposed VP2 structures [subunit (a)] of IPNV (PDB ID 9GG2) (purple) and IBDV (PDB ID 7VRN) (yellow). The root mean square deviation (RMSD) value between 321 pruned atom pairs of IPNV and IBDV VP2s is 0.79 Å (5.3 Å across all 387 atoms). (B) Residual RMSD values between IPNV and IBDV VP2 structures. (C, D) Close-up views of the aligned VP2 P-domains of IPNV (purple) and IBDV (yellow). Five IPNV (L1–4, L4′) and four IBDV (L1–4) surface loops are labeled.
The conserved jelly roll structure of the birnavirus VP2 S domain suggests an evolutionary relationship with ssRNA tetraviruses/nodaviruses within the order Nodamuvirales (21). Indeed, the jelly roll fold in VP2 of IPNV is similar to those found in capsid proteins of viruses from the Nodamuvirales order and Tetraviridae family, including Lake Sinai virus, Nudaurelia capensis omega virus, and Pariacoto virus (Table S1, Dali score > 10). These viruses also display birnavirus-like P-domains on their capsid surfaces (52–56), implying the importance of the P-domain in host-specific receptor binding across a broad range of icosahedral jelly roll viruses.
Unique structural interface of infectious T=13 IPNV particle
The T=13 capsid is composed of pentagonal capsomers, each containing five VP2 (a) subunits at each 5-fold axis and quasi-6-fold hexagonal capsomers formed by the remaining subunits. The adjacent VP2–VP2 interface within these capsomers is stabilized by hydrophobic interactions involving amino acid residues 138–157 of each VP2 subunit (Fig. 3A). However, unique structural interfaces are also observed in both capsomers. In the pentagonal capsomer, an additional hydrophobic interaction involving the Tyr398 residue contributes to VP2–VP2 stabilization, whereas in the hexagonal capsomer, hydrophobic interactions are mediated by the variable loop region (Fig. 3A).
Structure and unique subunit interface of the hexagonal capsomer in T=13 IPNV capsid. (A) Adjacent VP2–VP2 interface in the 5-fold pentagonal capsomer and quasi-6-fold hexagonal capsomer. Amino acid residues 138–157 are colored blue (indicated also by red circles). The side chain of Tyr398 in the 5-fold pentagonal capsomer is also shown in blue. The variable loop is colored by orange in the quasi-6-fold hexagonal capsomer (indicated also by a black circle). (B) Structural comparison of VP2 subunits between the T=1 subviral particle (gray, PDBID: 3IDE) and the T=13 infectious particle [purple, subunit (a)] of the IPNV capsid. Large parts of the VP2 variable loop (red dotted lines) are unresolved in these VP2 structures. Structural variations are observed in their variable loops (black circle) and N- and C-termini (black rectangle). Minor structural variations are also observed in surface loops (black rectangle). (C) Comparison of the “5-fold,” “Down," and "Up" conformations of the VP2 variable loops in the T=13 particle. The VP2 subunit with the "5-fold" conformation is shown in purple; the “Down” conformation is light blue; and the "Up" conformation is orange. (D) Structure of the hexagonal capsomer, where six VP2 subunits form hexagonal capsomers in the T=13 capsid, displaying the “Down” and “Up” conformations of the variable loops. (E) Close-up view of the variable loops at the center of the hexagonal capsomer. The Y115 residues in the three VP2 subunits with the "Up" conformation contribute to forming the quasi-6-fold in the hexagonal capsomer.
Structural comparison between T=1 and T=13 IPNV particles reveals differences in the variable loop regions of VP2 (Fig. 3B and C), which also exhibit structural diversity among the 13 VP2 subunits in the T=13 capsid (Fig. 1C). The structure of the VP2 variable loop in the subviral particle was not fully resolved (16), similar to the variable loop of the VP2 subunit (a) that is closest to the 5-fold axis in the T=13 particle (Fig. 3B). However, the variable loop of the other 12 VP2 subunits in the asymmetric unit adopts either an up or down conformation (Fig. 3C), contributing to the formation of the necessary structural interfaces within the hexagonal capsomers in the T=13 capsid lattice (Fig. 3D). Six variable loops of the VP2 subunits are located at the center of each hexagonal capsomer, with three up and three down conformations (Fig. 3B). The Tyr115 residues of the three up conformations largely contribute to forming the interface (Fig. 3D and E). A similar structural interface mediated by three Tyr118 residues in the up and down conformations of the variable loops (20) is observed in the hexagonal capsomer of IBDV, highlighting their critical role in forming the T=13 infectious particle in birnaviruses. The structural variations observed in the variable loops in the T=13 IPNV capsid arise from the formation of necessary hexagonal capsomers, which are absent in the T=1 subviral particle. Furthermore, these observations align with the structural resemblance between the VP2 subunit (a), which forms a pentagonal unit at the 5-fold axis, and the VP2 subunit in the T=1 subviral particle, which consists solely of pentagonal units (Fig. 3B).
C-terminal interlocking and possible assembly intermediates
The IPNV atomic model based on residues Thr7 to Asn428 from the T=1 subviral particle crystal structure (Fig. 3B) (24) revealed an extra intensity corresponding to the VP2 C-terminus in some VP2 subunits of the reconstructed T=13 IPNV model. Specifically, this intensity was observed from Glu429 to Ser435 in subunit (a), Glu429 to Ile430 in subunit (b), and Glu429 to Ser434 in subunit (c) (Fig. S6). The other VP2 subunits (d)–(m) were modeled up to Asn428, consistent with the T=1 subviral particle. This extension was previously unresolved in the T=13 IBDV infectious particle and T=1 IPNV subviral particle (20, 24), but its existence has been linked to a possible interlocking role between neighboring VP2 3-fold trimer units in the T=1 IBDV subviral particle (29). In the T=13 IPNV particle, the C-terminal extensions of subunits (a) and (c) are oriented differently (Fig. 1C; Fig. S4). In subunits (a) and (c), Asn428 and Glu429 in the C-terminal region form intrasubunit hydrogen bonds with Lys12 in the N-terminal region, whereas in the other subunits, they form hydrogen bonds with Arg425 (Fig. S6). This shift in the hydrogen bonding results in the distinct orientations of the N- and C-terminal regions in subunits (a) and (c).
The C-terminal extensions create an additional hydrophobic interface and interlock with neighboring subunits on the capsid interior (Fig. 4A and B; Fig. S6). The C-terminal region of subunit (a) (residues Thr431, Phe433–Ser435) interlocks with the adjacent subunit (a)′ at the N-terminal region (residue Lys5), at one α-helix of the B domain (residue Tyr398) and at the C-terminal region (residue Val426) (Fig. 4B; Fig. S6A). In contrast, the C-terminal region of subunit (c) (residues Thr431, Phe433) interlocks with the C-terminal region (residues Val426, Glu429) of the adjacent subunit (b) (Fig. 4C; Fig. S6B). The shorter C-terminal extension of subunit (b) however, does not orient to the neighboring subunit. Instead, it is located close to the C-terminal extension of subunit (c) (light blue in Fig. 4C; Fig. S6B), thus contributing to the interlocking nevertheless. As a result of these interlocking interactions, the five VP2 trimers form a penton protein-protein network complex (Fig. 4A). Long terminal extensions and similar interlocking mechanisms have also been identified in other icosahedral dsRNA viruses, such as toti-like viruses, picobirnaviruses, megabirnavirus, quadrivirus, and reoviruses (38, 42, 57–63), as well as short terminal extensions in Totiviridae viruses (40, 41, 64). These interlocking networks are thought to stabilize the virus capsid, which suggests that the C-terminal extensions in IPNV may have a similar function. The amino acid residues in the C-terminal extension are highly conserved among IPNV strains (Fig. S7), further supporting their critical role.
C-terminal extension of VP2 subunits and the intrasubunit interlocking in pentagonal structural units. (A) C-terminal interlocking network of VP2 subunits at the interior of a 5-fold vertex. Five VP2 subunits from each of the (a), (b), and (c) subunits form a pentagonal structural unit at the 5-fold vertex of the T=13 IPNV capsid. The C-terminal extensions of these subunits are shown in surface representation. Representative C-terminal extensions from subunits (a), (b), and (c) are shown in red. (B) Close-up view of the C-terminal extension region between adjacent subunits (a) and (a)′. (C) Close-up view of the C-terminal extension region between adjacent subunits (b) and (c). Interacting amino acid residues are labeled in panels B and C. (D) Triangular and pentagonal units of IPNV in a raw cryo-EM micrograph. (E) Two-dimensional class averages of the triangular and pentagonal units. Red dotted rectangles show protruded features of the pentagonal units.
The C-terminal extension could be important for assembling the T=13 capsid. During the maturation process of the IPNV polyprotein, the C-terminal region of pVP2 is expected to be cleaved in multiple sites (Ala442, Ala486, and Ala495) to form mature VP2 (65). It is anticipated that the C-terminus should exist at the Ala442 residue in the mature IPNV particle (27). Recombinant expression of only mature VP2 (residues 1–442) results in the formation of a T=1 subviral particle (Table S2), whereas the inclusion of the C-terminal region of pVP2 (residues 1–466) and the C-terminal domain of VP3 results in the formation of T=13 and T=1 icosahedrons, as well as tubular structures (22). Other previous studies have described the importance of post-processed C-terminal peptides in IPNV preVP2 (residues 443–508) and IBDV preVP2 (residues 443–512) for forming the T=13 capsid (27, 28). This study further highlights the additional importance of the previously unresolved C-terminal extension of VP2 (residues 429–435) (Fig. 4A through C). The structure and the interlocking function of the C-terminal extension could be critical for assembling the T=13 capsid and forming the required intermediate building blocks. The observed interlocked penton unit in the IPNV T=13 capsid does not exist in the T=1 subviral particle. The C-terminal interlocking could prevent the incorrect formation of T=1 capsids by assembling penton structural units during pVP2 maturation and assembly.
Many triangular and partial or complete pentagonal units were observed in the raw cryo-EM images of the IPNV sample (Fig. 4D). The 2D class averaging of these units revealed that the triangular units correspond to individual VP2 trimers, while the partial and complete pentagonal units consist of two–five trimer units (Fig. 4E). The formation of these pentagonal units from five triangular units aligns with previously speculated models for the IBDV assembly (23, 66). Notably, in the 2D classes of two–four trimers, the central region of the complex remains relatively closed, whereas the complete penton unit exhibits a more open structure (Fig. 4E). Comparison of the 5-fold pore structures composed of five VP2 subunits in the T=1 and T=13 IPNV capsids, excluding the variable loop region (residues 110–117), reveals a larger 5-fold pore size in the T=13 penton unit (19.8 Å in radius) compared to the T=1 penton unit (18.9 Å in radius) (Fig. S8). This suggests that the penton unit in the T=13 is more sparsely packed, potentially contributing to the observed larger pore in the full pentagonal unit (Fig. 4E). We also hypothesize that the helical structure of the VP2 trimers may form preferentially if the pore does not expand during pentagonal unit assembly possibly due to spatial constraints for accommodating the fifth VP2 trimer. Although the 3D reconstructions of these triangle and pentagonal units are performed, the clear 3D model has not been built yet because of preferred orientation. Additional protruding features are visible in the trimer units in the 2D projections of the pentagonal units, which are absent in the triangular units (red dotted rectangles in Fig. 4E). One speculation is that these protrusions may represent part of the C-terminal regions of pVP2 subunits that need to be rearranged during the transition from triangular to pentagonal units. Further studies on birnavirus assembly intermediates are essential to elucidate the role of the C-terminal extensions and the formation of pentagonal units.
Potential gate structure of IPNV
Five VP2 subunits [subunits (a) in Fig. 1B] form a 5-fold pore structure in the IPNV capsid. As discussed in previous studies of the T=13 IBDV capsid (20, 21), this pore may be obstructed by unresolved variable loops from the five VP2 subunits at each 5-fold axis. As aforementioned, the T=13 IPNV capsid also shows 5-fold pore structures with unresolved variable loops likely positioned within the pore (Fig. 4A; Fig. S8). The 5-fold focused asymmetric reconstruction of the IPNV pore indicates more pronounced obstruction, though a clear structure is not discernible, most likely due to the different conformations of the variable loops (Fig. 5A). It is also likely that the asymmetric reconstruction could still be influenced by the icosahedral symmetry of the capsid. Despite ongoing debate about intraparticle genome synthesis in birnaviruses, the IPNV pore shares similar characteristics with other dsRNA viruses known to facilitate intraparticle genome synthesis. The VP2 subunit exhibits two positively charged residues, Lys159 and Arg425, on the pore surface, which form clusters of positively charged regions (Fig. 5B and C). Similar clusters in other icosahedral dsRNA viruses are believed to be important for recruiting negatively charged NTPs to facilitate RdRp-mediated intraparticle genome synthesis (38, 40–42, 57). Moreover, in single-layered dsRNA viruses that infect multicellular hosts (metazoa), including artiviruses (arthropod totiviruses), mammalian picornaviruses, and fungal megabirnavirus, the gate of their pore is either closed or obstructed by flexible loops or additional proteins (42, 57, 58, 61, 63). Conclusively, these structural features of the salmonid IPNV capsid pore are akin to the previously observed pore structure of the metazoan dsRNA viruses. Understanding the genome synthesis mechanisms in these viruses necessitates determining the interior RNP structures (VP1 and VP3). Although asymmetric reconstruction methods were attempted according to previous dsRNA virus structural studies, a clear 3D model of the genome and RNP structures of IPNV could not be determined most likely due to heterogeneity. However, 2D class-averaged particle images of the capsid-subtracted data set reveal a non-typical organization of IPNV RNPs, differing from the partially spooled dsRNA genomes seen in Totiviridae and Reoviridae viruses (Fig. 5D) (38, 39, 44, 45, 67). This implies that IPNV employs a different genome packaging mechanism for intraparticle genome synthesis.
Potential gate for intraparticle genome transcription in birnavirus. (A) Five-fold pore structure as revealed by icosahedrally averaged (Icos) and symmetry expanded (Sym Exp) cryo-EM reconstructions of the IPNV capsid. The contour level of each cryo-EM map is adjusted to 5σ. (B) Top view of the 5-fold pore. (C) Cross-section of the 5-fold pore. (B, C) Residues 110–117 (unresolved variable loops) of the VP2 are removed for visualization. Positively charged residues of the gate, five K159 and R425 residues, are colored pink and orange, respectively. (D) Representative particle images obtained from capsid-subtracted two-dimensional class averaging.
MATERIALS AND METHODS
Cell culture and IPNV sample preparation
The IPNV SP strain (IPNV-L5) used in this study was isolated from head kidney samples obtained from cultured Atlantic salmon (Salmo salar L.), originating from sites with suspected IPNV infection. The initial isolation was performed in CHSE-214 cells, and the seed virus stock used for propagating IPNV-L5 samples in this study was propagated three times in RTG2 cells. The VP2 gene of IPNV L5 varied from the classical virulent isolate of IPNV (GenBank ID: AY379740) at 12 amino acid positions listed in Table S3.
The purification protocol for IPNV particles was optimized from previously reported papers (8, 68). Infected culture fluid (ICF) was centrifuged at 5,000 × g at 4°C for 30 min to remove cell debris. Solid PEG 20,000 and sodium chloride were added to the ICF to achieve final concentrations of 5 (w/v) and 2.2% (w/v), respectively, and the mixture was stirred overnight at 4°C. The virus-containing pellet was obtained by centrifuging the ICF at 15,300 × g at 4°C for 60 min. The pellet was resuspended in 1 mL of PBS (−). The sample was applied on a 15–45% (w/v) sucrose gradient and ultracentrifuged at 130,000 × g at 4°C for 2.5 h. The SDS-PAGE result of the sucrose gradient fractions is shown in Fig. S1A. The fractions containing filled particles with VP1, VP2, and VP3 protein bands were pooled and diluted in PBS (−) up to 13 mL. The diluted virus fractions were ultracentrifuged at 130,000 × g at 4°C for 2 h. The virus pellet was resuspended in 50 µL of PBS (−), and then concentrated to 20 µL. The final protein concentration of the sample was calculated to be 1.08 mg/mL, which was used for the cryo-EM grid preparation.
Cryo-EM data acquisition and map reconstruction
The data acquisition parameters are listed in Table S4. Cryo-EM grids were obtained by flash-freezing 3 µL of purified IPNV particles on glow-discharged holey carbon grids (Quantifoil R2/2, Cu 300 mesh; Quantifoil Micro Tools GmbH). The grids were plunge-frozen in liquid ethane after blotting for 3 s under 4°C and 100% humidity using a Mark IV Vitrobot (Thermo Fisher Scientific). The frozen grids were initially screened with a Glacios cryo-EM (Thermo Fisher Scientific) at the Uppsala Cryo-EM Center. The complete data set was collected with a Titan Krios (Thermo Fisher Scientific) installed with a Gatan K3 BioQuantum detector and an electron energy filter (20 eV slit width) at the SciLifeLab Cryo-EM facility. Image movies (30 frames per movie) were acquired in counted superresolution mode at a nominal magnification of 81,000×, corresponding to a pixel size of 1.058 Å/px. The total electron exposure was 26.8 e^−^/Å^2^ over 1.34 s per movie. Defocus values were randomly selected in 0.2 µm increments within the range of 0.7 to 1.5 µm underfocus for each movie.
A total of 27,945 movies were collected for three-dimensional (3D) cryo-EM model reconstruction. Image processing and 3D reconstruction were conducted using CryoSPARC version 4.3.1 (69) and a local CPU/GPU computer cluster. Initial preprocessing involved patch motion correction using frames 3–27, followed by contrast transfer function (CTF) estimation via patch CTF. Template picking and two-dimensional (2D classifications selected 35,498 particles, including 12,898 dsRNA genome-filled particles. The 3D reconstructions were performed on downsampled particles (1.284 Å/px). An initial model was generated from 6,471 particles via ab initio reconstruction. The best achievable resolution was obtained by processing all selected 35,498 particles using the homogeneous refinement option with imposed icosahedral symmetry. During refinement, per-particle defocus and per-group CTF parameters were optimized, with spherical aberration, beam tetrafoil, and anisotropic magnification fitting enabled. Ewald sphere correction was also performed. The final 3D map of IPNV L5 achieved a resolution of 2.75 Å based on Fourier shell correlation at a 0.143 cutoff (70) (Fig. S1C). Due to the large size of the particle, 70 nm in diameter, the block-based reconstruction was also performed (20, 71, 72); however, no significant improvement was observed. The obtained cryo-EM map was used for building atomic models of the VP2 subunits. The 2D classification of the IPNV interior structure was performed using capsid-subtracted images of the 12,898 filled particles. The structure of the IPNV capsid was also determined at a resolution of 2.78 Å using only filled particles, resulting in a structure closely resembling that obtained from all particles, with comparable side chain orientations (Fig. S3). This finding suggests that there are no significant structural differences between empty and filled IPNV capsids. Asymmetric reconstruction was done using a previously described symmetry expansion and local reconstruction method for icosahedral dsRNA viruses (38, 67). A mask covering one 5-fold vertex was applied during local reconstruction.
Atomic modeling, refinement, and rendering of IPNV capsid
The initial atomic model of the VP2 CP of IPNV-L5 was predicted from the VP2 sequence using AlphaFold2 (73). The highest-scoring predicted model was then first fitted into the subunit (a) region (the closest subunit to the 5-fold vertex) of the cryo-EM map using UCSF Chimera (74). The atomic model was then manually refined using Coot version 1.0.06 (75) and further automatically refined using PHENIX 1.20.1 (76) in iterative cycles. The refined atomic model of VP2 was then manually fitted to each of the other 12 VP2 subunit locations [subunits (b)–(m)] within an asymmetric unit of the icosahedral map. These 12 VP2s were also refined using Coot and Phenix software. Validation statistics of the atomic models are shown in Table S4. For rendering the cryo-EM maps and the atomic models, UCSF Chimera and ChimeraX were used (74, 77).
Structural comparison
The resolved structure of IPNV VP2 and corresponding CPs from other viruses that employ a jellyroll fold were extensively compared using the DALI server (78). The structural alignment of VP2 subunits from IBDV and IPNV capsid structures was generated using the TM-align algorithm (79), and the residual RMSD was calculated in PyMOL as described previously (36, 38, 79).
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Yang Z, He B, Lu Z, Mi S, Jiang J, Liu Z, Tu C, Gong W. 2021. Mammalian birnaviruses identified in pigs infected by classical swine fever virus. Virus Evol 7:veab 084. doi:10.1093/ve/veab 08434659797 PMC 8516818 · doi ↗ · pubmed ↗
- 2Evensen O, Rimstad E. 1990. Immunohistochemical identification of infectious pancreatic necrosis virus in paraffin-embedded tissues of Atlantic salmon (Salmo salar). J Vet Diagn Invest 2:288–293. doi:10.1177/1040638790002004061965633 · doi ↗ · pubmed ↗
- 3WOLF K, SNIESZKO SF, DUNBAR CE, PYLE E. 1960. Virus nature of infectious pancreatic necrosis in trout. Proc Soc Exp Biol Med 104:105–108. doi:10.3181/00379727-104-2574313845654 · doi ↗ · pubmed ↗
- 4Evensen Ø, Leong J-AC. 2013. DNA vaccines against viral diseases of farmed fish. Fish Shellfish Immunol 35:1751–1758. doi:10.1016/j.fsi.2013.10.02124184267 · doi ↗ · pubmed ↗
- 5Giambrone JJ, Donahoe JP, Dawe DL, Eidson CS. 1977. Specific suppression of the Bursa-dependent immune system of chicks with infectious bursal disease virus. Am J Vet Res 38:581–583.195492 · pubmed ↗
- 6Mahgoub HA, Bailey M, Kaiser P. 2012. An overview of infectious bursal disease. Arch Virol 157:2047–2057. doi:10.1007/s 00705-012-1377-922707044 · doi ↗ · pubmed ↗
- 7Dobos P. 1976. Size and structure of the genome of infectious pancreatic necrosis virus. Nucleic Acids Res 3:1903–1924. doi:10.1093/nar/3.8.1903987579 PMC 343048 · doi ↗ · pubmed ↗
- 8Dobos P, Hill BJ, Hallett R, Kells DT, Becht H, Teninges D. 1979. Biophysical and biochemical characterization of five animal viruses with bisegmented double-stranded RNA genomes. J Virol 32:593–605. doi:10.1128/JVI.32.2.593-605.1979228080 PMC 353591 · doi ↗ · pubmed ↗