Prevalence of Enterocytozoon bieneusi and Blastocystis sp. infection in foxes (Alopex lagopus) in northern China
Yan Tang, Hai-Tao Wang, Xue-Min Li, Zhong-Yuan Li, Qing-Yu Hou, Jing Jiang, Li-Hua Yang, Ya Qin

TL;DR
This study found high rates of E. bieneusi and low rates of Blastocystis sp. in foxes in northern China, with new genotypes identified that may pose public health risks.
Contribution
The discovery of six novel E. bieneusi genotypes and the first detection of Blastocystis ST3 in foxes.
Findings
E. bieneusi prevalence was 48.3%, with Shandong Province showing the highest rate at 87.1%.
Six novel E. bieneusi genotypes were identified, with CHN-F1 being the most common.
Blastocystis sp. was detected at 2.0% prevalence, with only subtype ST3 found in foxes for the first time.
Abstract
To examine the prevalence of E. bieneusi and Blastocystis sp. in foxes in China, this study analyzed the prevalence and distribution of genotypes or subtypes. A total of 352 fresh fecal samples were collected from foxes across five provinces in northern China and analyzed using PCR. The overall prevalences of E. bieneusi and Blastocystis sp. were 48.3% and 2.0%, respectively; the highest prevalences were found in Shandong Province, with 87.1% and 5.4%, respectively. The prevalence rates were influenced by several factors; a breeding scale value <1,500 was related to higher prevalences. Multivariate analysis showed that the region and breeding scale were the main risk factors for E. bieneusi. Eleven genotypes of E. bieneusi were identified, all of which are classified within Group 1. This includes five previously characterized genotypes and six novel genotypes. Among these, CHN-F1 was…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
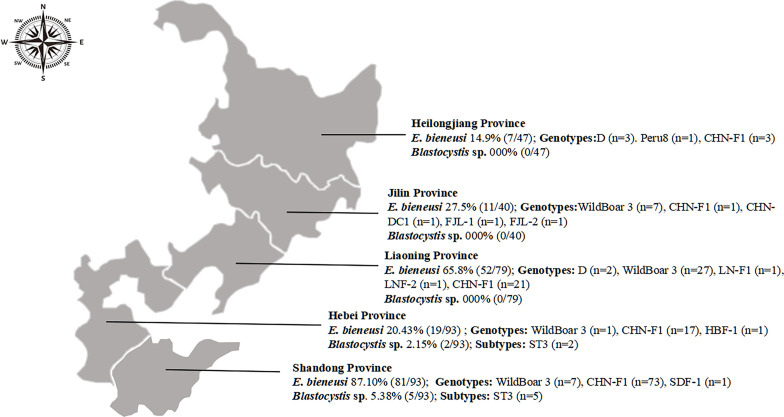 Figure 1
Figure 1 Figure 1
Figure 1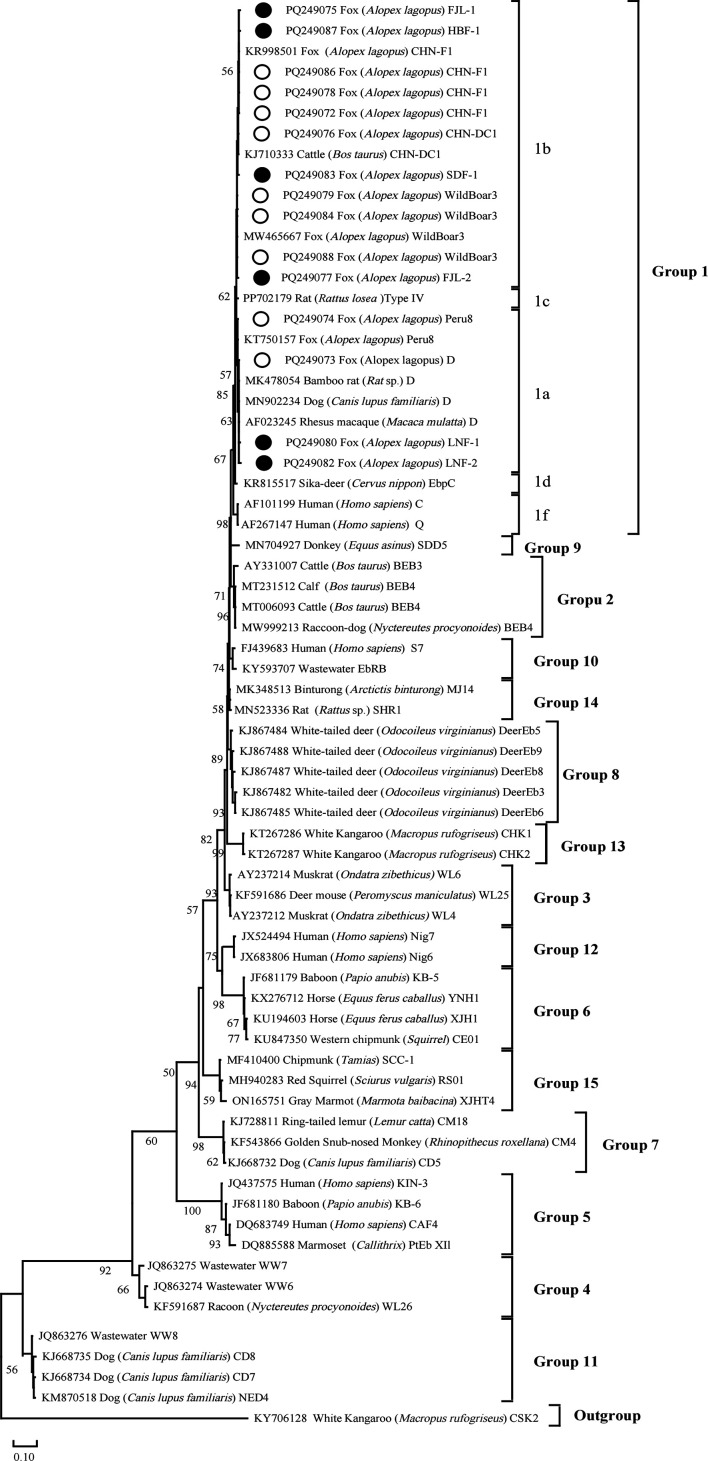 Figure 2
Figure 2 Figure 2
Figure 2- —The Science and Technology Program of Guizhou Province [Qian Ke He Foundation ZK(2024)General 365].
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsParasitic Infections and Diagnostics · Toxoplasma gondii Research Studies · Coccidia and coccidiosis research
Introduction
Enterocytozoon bieneusi and Blastocystis sp. are prevalent microeukaryotes [18, 33, 40, 67]. They can infect various hosts, including humans and foxes. Humans and animals become infected with E. bieneusi and Blastocystis sp. mainly through the fecal–oral ingestion of contaminated water or food [57]. Infections with two microeukaryotes are usually asymptomatic, but they can sometimes cause diarrhea [32, 67].
To date, at least 820 genotypes of E. bieneusi and 48 subtypes (STs) of Blastocystis sp. have been identified [32, 36, 39, 40, 54]. The E. bieneusi genotypes have been divided into 15 phylogenetic groups with different host specificity and zoonotic potential [32]. Group 1 is the largest, containing numerous zoonotic genotypes like D, Ebpc, and Peru 8, which are frequently detected in humans [24]. Even though certain genotypes like CHN-F1, NCF2, and NCF3 within Group 1 have not been detected in human patients thus far, they may still pose a zoonotic transmission risk [8, 32, 44]. The genotypes in Group 2 primarily infect ruminants, but also include the BEB4, BEB6, I, and J genotypes capable of infecting humans [36]. The remaining Groups 3–15 may exhibit host specificity, with some genotypes reported to have the ability to infect humans, such as Group 5 (KIN-3) and Group 6 (Nig 3 and May1) [15, 23]. More than one billion individuals worldwide are carriers of Blastocystis sp., with human infections involving ST1-10, ST12, ST14, ST16, ST23, ST35, and ST41 among the 48 identified subtypes. The prevalence of Blastocystis sp. infection in humans is predominantly attributed to ST1–4 which accounts for approximately 90.0% of cases [4, 21, 25, 29, 42, 65]. Although there are already reports on the prevalence of the E. bieneusi genotypes and the Blastocystis sp. subtypes, additional research is necessary to comprehensively understand their implications for public health [42].
A total of 361 E. bieneusi genotypes have been identified in China, including a variety of hosts, such as humans (5.7%, 188/3271, predominantly genotypes D and Ebpc), non-human primates (17.8%, 948/5318, predominantly genotype D), cattle (14.0%, 783/5594, predominantly genotypes J and I), sheep and goats (31.9%, 978/3068, predominantly genotype BEB6), pigs (55.9%, 1101/1969, predominantly genotypes Ebpc and EbpA), and blue foxes (17.9%, 108/603, predominantly genotype D) [53]. Blastocystis sp. is also widely distributed in China, the average infection rate among humans is 3.4% (3625/107,695), with ST3 being the predominant genotype [10]. Among other animals, the overall infection rate in Artiodactyla (including pigs, cattle, sheep, goats, reindeer, and sika and barking deer) is 27.4% (1050/3828), while in Carnivora (including raccoon domestic dogs and Arctic foxes), the infection rate is 2.8% (11/389) [10]. Another study involved a meta-analysis, showing that the global prevalence of Blastocystis sp*.* was 2.1% (29/1377) in free-living carnivores, compared to a significantly higher prevalence of 8.5% (100/1175) in captive individuals [5].
In China, the fox is an economically important animal, with approximately 3.44 million breeding foxes [66]. However, there is limited research on the prevalence of E. bieneusi and Blastocystis sp. in foxes, as well as their genetic characteristics. This study aimed to investigate the infection and distribution of the E. bieneusi genotypes and the Blastocystis sp. subtypes in foxes in China. Understanding the genetic evolutionary relationship between E. bieneusi and Blastocystis sp. in foxes can help us to effectively prevent and control diseases among fox populations as well as safeguard public health in China.
Materials and methods
Ethics statement
All the procedures used in this study were approved by the Research Ethics Committee for the Care and Use of Laboratory Animals in Qingdao Agricultural University, China. Written informed consent was obtained from the owners of the animals involved in this study.
Collection of samples
From October 2023 to June 2024, a total of 352 feces samples from foxes (Alopex lagopus) were collected across five provinces in northern China. Specifically, the samples were collected from three farms in Heilongjiang (sample number = 47, taken in 2024), two farms in Jilin (sample number = 40, taken in 2024), six farms in Liaoning (sample number = 10, taken in 2023; sample number = 69, taken in 2024), six farms in Hebei (sample number = 93, taken in 2024), and five farms in Shandong (sample number = 93, taken in 2023). Fresh fecal samples, representing approximately 1.0 to 1.5% of the total farmed population, were randomly collected from each farm. All the farms underwent biannual deworming treatments using either ivermectin or albendazole. Diarrhea is defined as the presence of green, watery, or bloody stools. The samples were collected from multiple locations within the farms to avoid being limited to a single area or cage. During the collection process, samples that met the defined criteria for diarrhea were recorded as diarrheal samples. The sampling process involved recording essential information, including the region, the breeding scale, the age of the fox, and the presence of diarrhea, which were documented during the data collection process. The collected samples were transported to a laboratory on ice within 24–48 h and stored at −80 °C until DNA extraction. The storage duration ranged from 1 week to 10 months.
DNA extraction and PCR amplification
DNA extraction from all the samples was conducted utilizing an EZNAR Stool DNA Kit (OMEGA Biotek Inc., Norcross, GA, USA), with the material subsequently stored at −20 °C. The DNA of all the samples was amplified using PCR to determine the presence of E. bieneusi, or Blastocystis sp., based on the ITS region and the small subunit (SSU) rRNA gene [57, 59], respectively. Novoprotein 2×Taq Master Mix (Novoprotein Ltd., E005-01, Suzhou, China) was used for all the PCR amplifications. The reaction conditions and sizes of the product fragments are presented in Table 1. Enterocytozoon bieneusi (genotype D) and Blastocystis sp*.* (subtype 3)-positive DNA previously detected in a laboratory from wild rodents was used as the positive control, while dd H_2_O was employed as the negative control [15, 16]. The PCR products were visualized using the QuickGel 6200 gel imaging System (Mona Biotechnology Co., Ltd., GD50102, Suzhou, China) after electrophoresis on 1% agarose gels.
Table 1. Primers used in the characterization of E. bieneusi and Blastocystis sp.GenePrimer (5′–3′)Product size (bp)Annealing temperature (°C)SSU rRNA of Blastocystis sp.SSU rRNA-F: ATCTGGTTGATCCTGCCAGT60055SSU rRNA-R: GAGCTTTTTAACTGCAACAACGITS gene of E. bieneusiITS-F1: GGTCATAGGGATGAAGAG39257ITS-R1: TTCGAGTTCTTTCGCGCTCITS-F2: GCTCTGAATATCTATGGCT55ITS-R2: ATCGCCGACGGATCCAAGTG
Sequencing and phylogenetic analyses
All amplicons with the expected size were sequenced bidirectionally at the General Biol. Company in Anhui, China. Then, the obtained sequences were blasted in GenBank to finally confirm the positive samples (http://www.ncbi.nlm.nih.gov/BLAST/). The genotypes of E. bieneusi were determined and named based on ~243 bp of the ITS region, according to the established nomenclature system [38]. Phylogenetic trees were constructed using the neighbor-joining (NJ) method in MEGA 11 (v. 11.0.13), with bootstrapping performed using 1,000 replicates to assess the genetic relationships among the E. bieneusi genotypes. CD-HIT (Version 4.8.1) was employed to remove the redundant sequences, solely preserving the unique sequences at a 100% identity threshold. The unique sequences, which serve as representatives of their respective clusters, were subsequently submitted to GenBank (accession Nos. PQ249072–PQ249088 and PQ223428–PQ223432).
Statistical analysis
The chi-square test of SAS (SAS Institute Inc., Cary, NC, USA, v.9.4) was utilized to conduct statistical analysis of the region, the breeding scale, the age of the foxes, and the presence of diarrhea. Forward stepwise logistic regression, using the Fisher scoring technique, was applied to a binary Logit model to assess the impact of multivariate regression analysis on E. bieneusi and Blastocystis sp. infection. All the tests were two-sided, and the results were considered statistically significant if p < 0.05, with corresponding odds ratios (ORs) and their 95% confidence intervals (95% CIs) provided [63].
Results
Prevalence of Enterocytozoon bieneusi and Blastocystis sp.
This study identified a prevalence of 48.3% (170/352, 95% CI [42.97–53.65]) for Enterocytozoon bieneusi and 2.0% (7/352, 95% CI [0.80–4.05]) for Blastocystis sp. Among the 352 foxes examined (Tables 2 and 3). Enterocytozoon bieneusi was detected across all five provinces, with the highest prevalence observed in Shandong (87.1%, 81/93, 95% CI [79.43–93.24]), followed by Liaoning (65.8%, 52/79, 95% CI [54.96–75.93]), Jilin (27.5%, 11/40, 95% CI [14.61–42.51]), Hebei (20.4%, 19/93, 95% CI [12.79–29.28]), and Heilongjiang Province (14.9%, 7/47, 95% CI [5.93–26.70]) (Table 2). The occurrence of Blastocystis sp. was relatively low, being detected solely in Shandong and Hebei Provinces, with infection rates of 5.4% (5/93, 95% CI [1.54–11.06]) and 2.2% (2/93, 95% CI [0.03–6.37]), respectively (Table 3).
Table 2. Factors associated with prevalence of E. bieneusi in foxes.FactorCategoryNo. positive/tested% (95% CI*)Univariate meta-regressionMultivariate analysis^c^OR (95% CI)p-ValueOR (95% CI)p-ValueRegionHeilongjiang Province7/4714.9 (5.93–26.70)Reference^d^<0.00010.408 (0.339–0.491)<0.0001Jilin Province11/4027.5 (14.61–42.51)2.17 (0.75–6.27)Liaoning Province52/7965.8 (54.96–75.93)11.05 (4.35–27.83)Hebei Province19/9320.4 (12.79–29.28)1.47 (0.57–3.79)Shandong Province81/9387.1 (79.43–93.24)39.52 (2.95–11.67)Breeding scale<150083/15354.3 (46.30–62.10)1.53 (1.00–2.33)0.05060.468 (0.275–0.796)<0.0001≥150087/19943.7 (36.88–50.68)ReferenceDiarrheaYes45/7460.8 (49.39–71.68)1.90 (1.13–3.21)0.0156––No125/27845.0 (39.15–50.85)ReferenceAge of FoxAdult^a^53/18229.1 (22.73–35.95)Reference<0.0001––Juvenile^b^117/17068.8 (61.64–75.59)12.38 (7.15–21.45)Total170/35248.3 (42.97–53.65)CI*: confidence interval; a: ≥1 year old; b: <1 year old; c: Only effect factors are listed; d: control group for risk analysis.
Table 3. Factors associated with prevalence and subtypes of Blastocystis sp. in foxes.FactorCategoryNo. positive/tested% (95% CI*)Univariate meta-regressionp-ValueOR (95% CI)RegionHeilongjiang Province0/470 (0.00–3.63)0.0927–Jilin Province0/400 (0.00–4.25)–Liaoning Province0/790 (0.00–2.16)–Hebei Province2/932.2 (0.03–6.37)Reference^c^Shandong Province5/935.4 (1.54–11.06)2.59 (0.49–13.68)Breeding scale<15005/1533.3 (0.93–6.79)0.14393.33 (0.64–17.40)≥15002/1991.0 (0.02–3.01)ReferenceDiarrheaYes3/744.1 (0.51–10.02)0.1673ReferenceNo4/2781.4 (0.31–3.25)2.89 (0.63–13.23)Age of FoxAdult^a^2/1821.1 (0.02–3.28)0.2323ReferenceJuvenile^b^5/1702.9 (0.83–6.12)2.73 (0.52–14.25)Total7/3522.0 (0.80–4.05)CI*: confidence interval; a: ≥1 year old; b: <1 year old; c: control group for risk analysis.
Statistical analysis revealed that the infection rate of E. bieneusi was significantly higher in foxes with a breeding scale value of less than 1500 (54.3%, 83/153, 95% CI [43.60–62.10]) compared to those with a breeding scale value greater than 1500 (43.7%, 87/199, 95% CI 36.88–50.68) (p < 0.0001). Notably, the prevalence of E. bieneusi was 60.8% in diarrheal foxes, which is significantly higher than that in non-diarrheal foxes (45.0%, 125/278, 95% CI [39.15–50.85]) (p = 0.0156). Additionally, the prevalence in juvenile foxes (68.8%, 117/170, 95% CI [61.64–75.59]) was significantly higher than that in adult foxes (29.1%, 53/182, 95% CI [22.73–35.95]) (p < 0.001, Table 2).
Similarly, the infection rate of Blastocystis sp. was higher in foxes with a breeding scale value of less than 1500 (3.3%, 5/153, 95% CI [0.93–6.79]) compared to those with a breeding scale value greater than 1500 (1.0%, 2/199, 95% CI [0.02–3.01]) (p = 0.0927). The prevalence of Blastocystis sp. was 4.1% (3/74, 95% CI [0.51–10.02]) in diarrheal foxes, which is higher than in non-diarrheal foxes (1.4%, 4/278, 95% CI [0.31–3.25]) (p = 0.1673). Furthermore, the prevalence in juvenile foxes (2.9%, 5/170, 95% CI [0.83–6.12]) was significantly higher than that in adult foxes (1.1%, 2/182, 95% CI [0.02–3.28]) (p = 0.2323, Table 3).
Risk factors of E. bieneusi and Blastocystis sp.
The region and breeding scale were significant in the final model, indicating its strong influence on E. bieneusi. The region and breeding scale of foxes had a strong effect on E. bieneusi infection in this study. The foxes in Shandong Province (OR = 0.408, 95% CI [0.339–0.491]) appear to exhibit a higher susceptibility to E. bieneusi compared to those in Liaoning, Heilongjiang, Jilin, and Hebei Provinces. A breeding scale value < 1500 (OR = 0.468, 95% CI [0.275–0.796]) indicates greater susceptibility to E. bieneusi than a value ≥ 1500. No significant influencing factors were identified in the presence of Blastocystis sp. in the foxes.
Distribution of E. bieneusi genotypes and Blastocystis sp. subtypes
This study identified eleven genotypes of E. bieneusi and one subtype of Blastocystis sp. among the 352 fox samples. The genotypes of E. bieneusi comprise five known (D, Peru 8, WildBoar 3, CHN-F1, and CHN-DC1) and six newly identified genotypes (LNF-1, LNF-2, HBF-1, SDF-1, FJL-1, and FJL-2). These new genotypes contain a total of 11 polymorphic sites (Table 4). In this study, the recently discovered genotype CHN-F1 exhibited the highest prevalence rate, followed by NCF3, NCF2, and D. Type D was detected in Heilongjiang and Liaoning Provinces. Notably, CHN-F1 was detected in all the five provinces, whereas the remaining new genotypes LNF-1 and LNF-2 were found only in Liaoning Province, SDF-1 was detected in Shandong Province, and HBF-1 was found in Hebei Province. The ST3 subtype of Blastocystis sp. was exclusively detected in this study, limited to Hebei and Shandong Provinces (Fig. 1).
Figure 1. Distribution of E. bieneusi genotypes and Blastocystis sp*.* subtypes.
Table 4. Variations in the ~243 ITS nucleotide sequences among genotypes of the E. bieneusi in foxes.Genotypes (No.)Nucleotide at positionGenBank accession No.174448657793117131191209212D (Reference)GTTGGCTGGTGAF023245 (Intergenic spacer)D (5) PQ249073 CHN-DC1 (1)ATGA PQ249076 FJL-1 (1)ACATGA PQ249075 CHN-F1 (115)AATGA PQ249072 HBF-1 (1)AATGAA PQ249087 LN-F1 (1)A PQ249080 LNF-2 (1)AC PQ249082 FJL-2 (1)CTGA PQ249077 WildBoar 3 (42)TGA PQ249079 Peru8 (1)G PQ249074 SDF-1 (1)AGA PQ249083
Phylogenetic analysis
Phylogenetic trees were constructed to represent the eleven E. bieneusi genotypes (Fig. 2) and the one Blastocystis sp. subtype identified in this study. It was observed that all the eleven E. bieneusi genotypes were grouped in Group 1, with D, Peru8, and the novel genotypes LNF-1 and LNF-2 classified within Group 1a. The remaining detected genotypes CHN-F1, CHN-DC1, and WildBoar 3, as well as the newly discovered genotypes FJL-1, SDF-1, FJL-2, and HBF-1, were classified into Group 1b.
Figure 2. Neighbor-joining tree of E. bieneusi ITS genotypes. Phylogenetic relationships of ITS nucleotide sequences of the E. bieneusi genotypes identified in this study and other reported genotypes. The phylogeny was inferred by a neighbor-joining analysis. Bootstrap values were obtained using 1000 replicates; those with values >50% are shown on the nodes. The genotypes in this study are marked by empty circles, and the novel genotypes are marked by filled circles.
Discussion
Enterocytozoon bieneusi and Blastocystis sp. are significant zoonotic intestinal protozoa that have been understudied in foxes [1]. This study employed molecular epidemiologic methods to investigate the prevalence of these protozoa among foxes in northeastern China. Additionally, this study characterized the predominant E. bieneusi genotypes and Blastocystis sp. subtypes in foxes and evaluated the potential zoonotic risk.
The prevalence of E. bieneusi was 48.3% (170/352), significantly higher than that reported in previous studies in foxes in China, such as 27.7% (53/191) [55], 16.4% (18/110) [66], and 12.3% (37/302) [62]. These studies employed the same PCR method as in the current study. In this study, the prevalence rate of Blastocystis sp. was 2.0%. Other studies have reported that the prevalence of Blastocystis sp. in foxes is 1.9% (4/213) in China [46] and 2.2% (4/213) in Spain [6]. These findings are consistent with the prevalence observed in our study. Strict carnivores tend to exhibit a lower prevalence of Blastocystis sp. than omnivorous or herbivorous species, and Blastocystis sp. in foxes may be caused by the contamination of food or water sources [5, 45].
Regional subgroup analysis revealed significant variation in E. bieneusi prevalence across the five provinces (p < 0.05). The prevalence in this study is much higher than that in previous studies, such as in Jilin (7.7%) [62], Heilongjiang (7.1–27.7%) [55, 62], Hebei (17.7%) [62], and Xinjiang (2.9%) [64]. The previous studies reported 1.9% Blastocystis sp. prevalence in Jilin and Liaoning Provinces [46]. In comparison, the prevalence in Shandong and Hebei Provinces was higher than that given in previous reports. This study presents the first epidemiologic survey on Blastocystis sp. in foxes conducted in Hebei Province. Although it exhibited a low positive rate, this should still raise concerns among individuals. In provinces with a high prevalence of both E. bieneusi and Blastocystis sp., it is important to monitor these microeukaryotes, although their veterinary significance and impact on foxes remain unclear. This includes ensuring strict adherence to environmental hygiene practices on farms and regular disinfection protocols, as well as routine testing. Furthermore, this study revealed a correlation between the breeding scale and the prevalence of these two protists, with farms having a breeding scale value <1500 exhibiting higher prevalence than those with a breeding scale value ≥1500. This could be attributed to the fact that large-scale farms possess greater expertise in feeding management and disease prevention and control [52].
In this study, the infection rate of E. bieneusi was higher in the diarrheal foxes than it was in the non-diarrheal samples (p < 0.05); a similar observation has been made in humans and cattle [36, 61]. Among patients exhibiting various degrees of diarrhea, the prevalence of E. bieneusi is notably higher compared to that among individuals without gastrointestinal (GI) disorders [3, 10, 60]. This may be because E. bieneusi can indeed cause diarrhea in the host, particularly under conditions of self-limiting infection or impaired host immunity [32]. Although the infection rate of Blastocystis sp. was higher in the diarrheal samples than it was in the non-diarrheal samples, the difference was not statistically significant (p > 0.05).
Both E. bieneusi and Blastocystis sp. exhibited higher infection rates in juvenile foxes compared to those of the adult foxes. The findings of this study are consistent with prior research, indicating that juvenile foxes are more vulnerable to infection [8, 44]. Further epidemiological data and laboratory studies on their pathogenic mechanisms may be necessary to provide a scientific explanation for the differing infection rates of these two protozoa species in juvenile and adult foxes.
Eleven genotypes of E. bieneusi belonging to Group 1 were identified in the foxes in this study, including five known (D, Peru8, WildBoar 3, CHN-F1, and CHN-DC1) and six newly discovered genotypes (LNF-1, LNF-2, HBF-1, SDF-1, FJL-1, and FJL-2). Genotype D exhibits a strong capacity for cross-species transmission [14], and has been reported in humans in China, highlighting the need for vigilance regarding the potential transmission risk from foxes to humans [27, 35, 49, 50]. Additionally, the genotype Peru 8 has been reported to infect humans in other studies [2, 7]. Although only one Peru8-positive sample was found in this study, this genotype has been consistently detected in foxes, suggesting that they are susceptible hosts for genotype Peru 8 [62]. Notably, genotype CHN-F1 was predominant, found in all the subgroups, with a prevalence of 67.7% (115/170). First identified in dairy cattle in 2015, CHN-F1 has also been detected in raccoon dogs and has exhibited high prevalence rates in foxes in Henan Province (52.1%, 25/48) [44, 55, 66]. Aside from China, only two reports from Europe indicate that CHN-F1 can infect pigeons and chinchillas, suggesting its potential for global spread [22, 68]. The prevalence rates of WildBoar 3 and CHN-DC1 among the positive samples were 24.7% (42/170) and 0.6% (1/170), respectively. WildBoar 3 exhibits a broad spectrum of host adaptability and has been documented in wild boars from Poland, Czechia [30], and Spain [9], as well as in red foxes and stone martens from Portugal [13], and in goats from China [11]. Other reports confirm their presence in foxes, indicating their common occurrence [8, 62]. Phylogenetic analysis identified six new genotypes of E. bieneusi. Group 1 is recognized for its zoonotic potential, indicating that these six novel genotypes may have the capacity for cross-host transmission [20]. Therefore, it is crucial to maintain ongoing attention to these genotypes in future investigations. Consequently, individuals such as fox breeders and those with weakened immunity – such as patients with HIV, children, and the elderly – should be especially vigilant regarding the risk of E. bieneusi transmission from foxes.
Only the ST3 subtype was detected in Blastocystis sp. in this study. This study was the first to identify the ST3 subtype in foxes. Previous studies have reported the presence of the ST1, ST4, and ST7 subtypes in foxes from northeast China [46], while foxes in Spain were found to harbor the ST7 and ST14 subtypes [6]. ST3 is the most common genotype in humans and the predominant subtype of Blastocystis sp. infections in China, with prevalence rates ranging from 43.8% to 62.0% [10, 31, 34, 37]. Another study has shown that the sequences of ST3 subtypes in dairy cows are identical to those found in exposed humans [17], suggesting a potential risk of ST3 cross-species transmission between humans and animals. It should be noted that there is 99.7% homology between the ST3 subtype (PQ223430) studied here and that in humans (MK782518). This indicates potential for the transmission of Blastocystis sp. between foxes and humans. However, additional follow-up studies are required to confirm this hypothesis. Other researchers should concurrently collect fecal samples from both feeders and foxes during the sampling process to determine whether such transmission is possible. ST3 is also widely prevalent among various animal species in China, such as rodents [48], wild rhesus macaques [56], Pallas’s squirrels (Callosciurus erythraeus) [28], pet dogs [26], rabbits [58], pigs [19, 51], pigeons [43], raccoon dogs [46], and beef [47] and dairy cows [12, 41]. Given that ST3 is widely distributed in both humans and a variety of animals in China, it is important to continue investigating the ecological dynamics and transmission routes of Blastocystis sp.
This study investigated the prevalence of E. bieneusi genotypes and Blastocystis sp. subtypes in Chinese foxes, providing compelling evidence for the occurrence of infection in this particular host species. However, it is important to acknowledge certain limitations associated with this research. The sampling scope was confined to northeast China, potentially limiting the generalizability of the prevalence rates of E. bieneusi and Blastocystis sp. in foxes across China. Additionally, this study did not account for variations among fox breeds, which could influence the infection status and genetic distribution of these protists. Therefore, future research should broaden the sampling area and increase the sample size, while also meticulously documenting relevant sample information.
Conclusion
There was a significant prevalence of E. bieneusi among foxes, with genotype CHN-F1 being widely distributed. Additionally, foxes carried genotypes D and Peru 8, which infect humans. Furthermore, new genotypes of E. bieneusi (LNF-1, LNF-2, SDF-1, HBF-1, FJL-1, and FJL-2) were identified for the first time, all belonging to Group 1 with a potential zoonotic risk. ST3 was found to be the predominant subtype responsible for human Blastocystis sp. infection in China. Moreover, this study provides the first evidence that foxes serve as carriers of ST3. Consequently, foxes represent a plausible source of transmission for both E. bieneusi and Blastocystis sp. infection. This investigation contributes to our understanding of the prevalence and genetic diversity of E. bieneusi and Blastocystis sp. among the fox populations of China, while highlighting its public health significance in preventing potential zoonotic transmission mediated by foxes.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Abarca N, Santin M, Ortega S, Maloney JG, George NS, Molokin A, Cardona GA, Dashti A, Koster PC, Bailo B, Hernandez-de-Mingo M, Muadica AS, Calero-Bernal R, Carmena D, Gonzalez-Barrio D. 2021. Molecular detection and characterization of Blastocystis sp. and Enterocytozoon bieneusi in cattle in northern Spain. Veterinary Sciences, 8, 191.34564585 10.3390/vetsci 8090191 PMC 8473172 · doi ↗ · pubmed ↗
- 2Akinbo FO, Okaka CE, Omoregie R, Dearen T, Leon ET, Xiao L. 2012. Molecular epidemiologic characterization of Enterocytozoon bieneusi in HIV-infected persons in Benin city, Nigeria. American Journal of Tropical Medicine and Hygiene 86, 441–445.22403314 10.4269/ajtmh.2012.11-0548 PMC 3284359 · doi ↗ · pubmed ↗
- 3Aydemir S, Halidi AG, Ekici A, Cengiz ZT. 2023. Investigation of the presence of Enterocytozoon bieneusi and Encephalitozoon intestinalis in immunosuppressed patients with diarrhea by IFA and real time PCR methods. Indian Journal of Medical Microbiology 44, 100362.37356849 10.1016/j.ijmmb.2023.02.005 · doi ↗ · pubmed ↗
- 4Aykur M, Malatyali E, Demirel F, Comert-Kocak B, Gentekaki E, Tsaousis AD, Dogruman-Al F. (2024). Blastocystis: a mysterious member of the gut microbiome. Microorganisms, 12, 461.38543512 10.3390/microorganisms 12030461 PMC 10972062 · doi ↗ · pubmed ↗
- 5Caballero-Gomez J, Avalos G, Matas-Mendez P, Figueiredo AM, Castro-Scholten S, Jimenez-Martin D, Koster PC, Santin M, Bailo B, Cano-Terriza D, Sarmento P, Neves N, Carrapato C, Gonzalez-Barrio D, Mateo M, Garcia-Bocanegra I, Dashti A, Sanchez S, Carmena D. 2025. Dietary profiles of wild carnivores and Blastocystis occurrence: the case of the endangered Iberian lynx (Lynx pardinus) and systematic review. Research in Veterinary Science 184, 105518.39765196 10.1016/j.rvsc.2024.105518 · doi ↗ · pubmed ↗
- 6Calero-Bernal R, Santin M, Maloney JG, Martin-Perez M, Habela MA, Fernandez-Garcia JL, Figueiredo A, Najera F, Palacios MJ, Mateo M, Balseiro A, Barral M, Lima-Barberoi JF, Koster PC, Carmena D. 2020. Blastocystis sp. subtype diversity in wild carnivore species from Spain. Journal of Eukaryotic Microbiology 67, 273–278.31691450 10.1111/jeu.12772 · doi ↗ · pubmed ↗
- 7Chabchoub N, Abdelmalek R, Breton J, Kanoun F, Thellier M, Bouratbine A, Aoun K. 2012. Genotype identification of Enterocytozoon bieneusi isolates from stool samples of HIV-infected tunisian patients. Parasite, 19, 147–151.22550625 10.1051/parasite/2012192147 PMC 3671435 · doi ↗ · pubmed ↗
- 8Chen M, Wang H, Li X, Guo Y, Lu Y, Zheng L, Liang G, Sui Y, Wang B, Dai H, Dong H, Zhang L. 2024. Molecular epidemiology of Enterocytozoon bieneusi from foxes and raccoon dogs in the Henan and Hebei Provinces in China. BMC Veterinary Research, 20, 53.38341563 10.1186/s 12917-024-03883-6PMC 10858577 · doi ↗ · pubmed ↗