Reciprocal interactions between CA1 pyramidal and axo-axonic cells control sharp wave-ripple events
Daniel English, Earl Gilbert, Lianne Klaver, Kaiser Arndt, Jongwoon Kim, Xiaoting Jia, Sam Mckenzie

TL;DR
This study explores how interactions between pyramidal cells and axo-axonic cells in the hippocampus control sharp wave-ripple events, which are important for memory consolidation.
Contribution
The study identifies axo-axonic cells as key regulators of sharp wave-ripple dynamics through reciprocal interactions with pyramidal cells.
Findings
Axo-axonic cells show variable spiking during sharp wave-ripples, unlike other interneurons.
Optogenetic silencing of axo-axonic cells increases sharp wave-ripple power and duration.
Pyramidal cells influence axo-axonic cell activity through monosynaptic inputs.
Abstract
Diverse sources of inhibition serve to modulate circuits and control cell assembly spiking across various timescales. For example, in hippocampus area CA1 the competition between inhibition and excitation organizes spike timing of pyramidal cells (PYR) in network events, including sharp wave-ripples (SPW-R). Specific cellular-synaptic sources of inhibition in SPW-R remain unclear, as there are > 20 types of GABAergic interneurons in CA1. Axo-axonic cells (AAC) are defined by their synaptic targeting of the axon initial segment of pyramidal cells, potently controlling spike output. The impact of AAC activity on SPW-R is controversial, due mainly to ambiguity of AAC identification. Here we monitored and manipulated opto-tagged AACs in behaving mice using silicon probe recordings. We found a large variability of AAC neurons, varying from enhanced to suppressed spiking during SPW-Rs, in…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
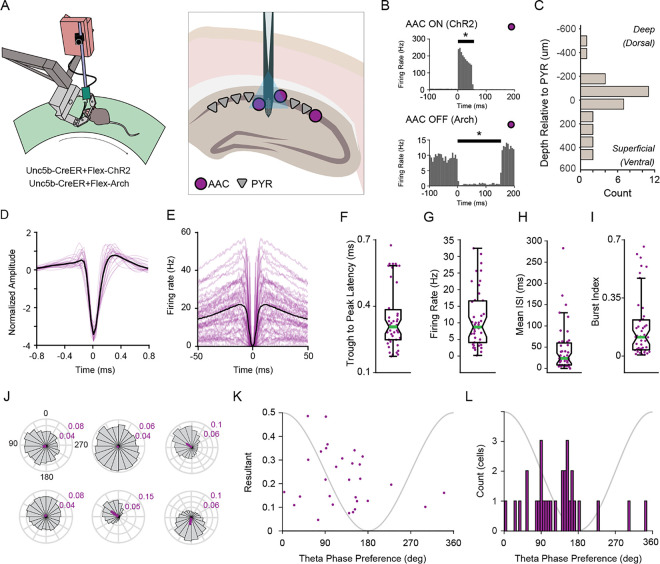 Figure 1
Figure 1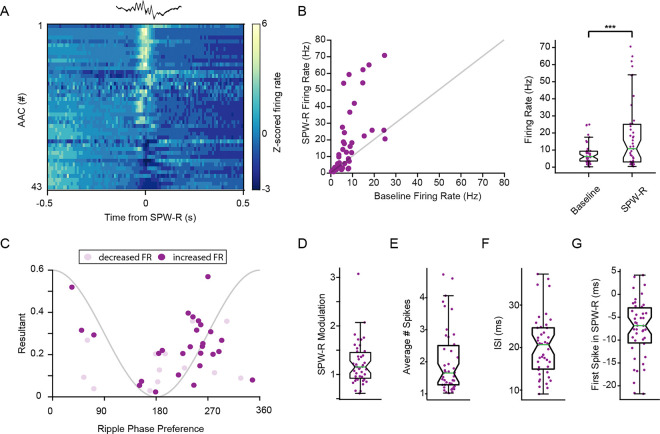 Figure 2
Figure 2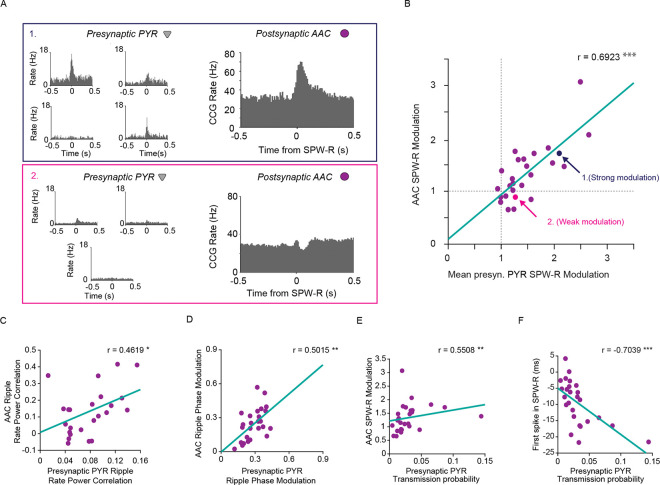 Figure 3
Figure 3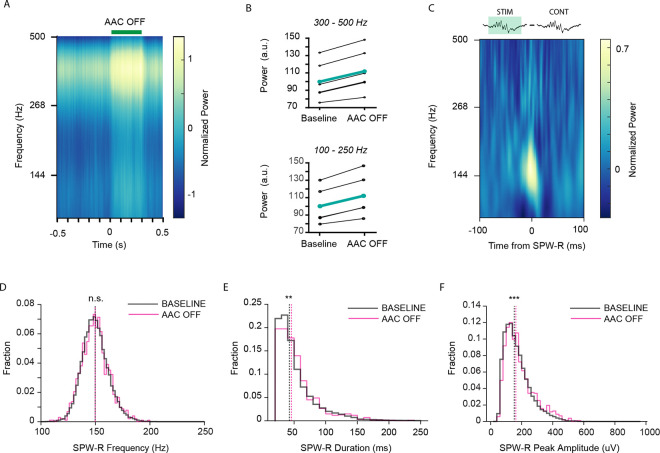 Figure 4
Figure 4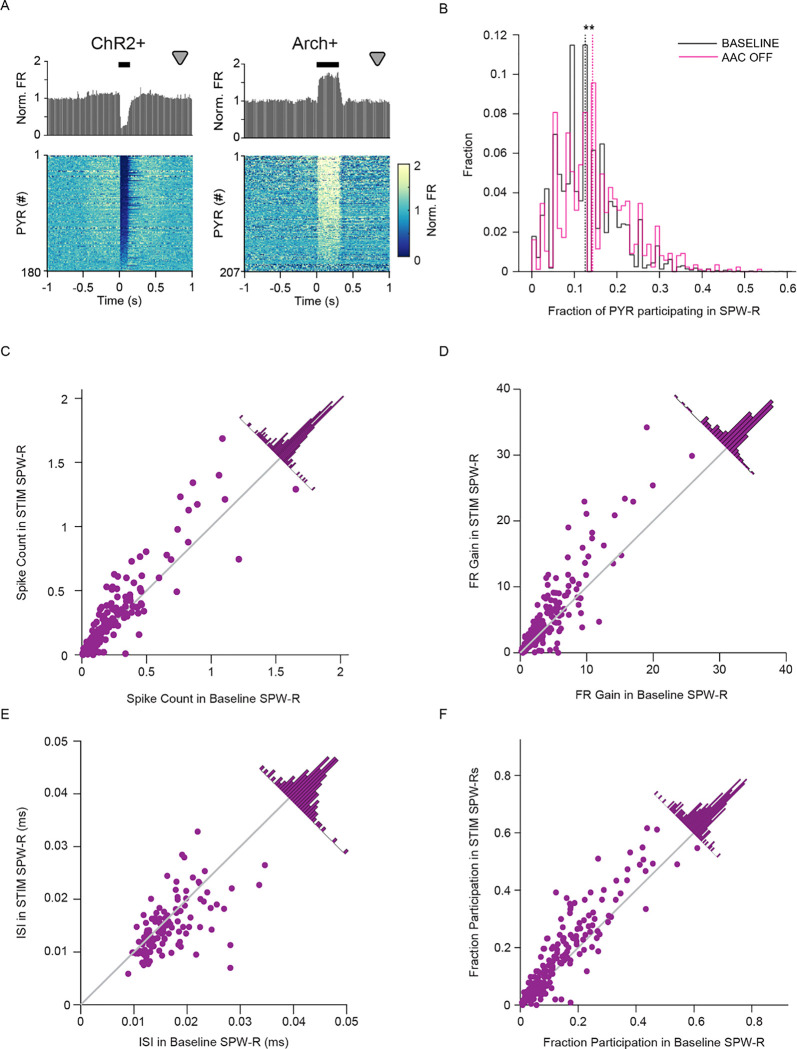 Figure 5
Figure 5Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsPhotoreceptor and optogenetics research · Microtubule and mitosis dynamics · Neuroscience and Neuropharmacology Research
Introduction
Inhibitory actions in the cortex and hippocampus are diverse, with > 20 types of inhibitory interneurons existing along a spectrum, with varied genetic profiles, axonal targeting, and state dependent activity^1–5^. Given the complexity that emerges from the nonlinear dynamics of inhibitory interactions among these types, numerous functions have been attributed to both their global and specific operations^6–8^. Studying unique interneuron subtypes in the hippocampus has proven fundamental in understanding mechanisms underlying circuit function, including their role in various network oscillations, learning, place field tuning and replay of experience^3,4,9–13^.
Axo-axonic cells (AACs) are anatomically distinct from other interneurons in two ways. First, only AACs target the axon initial segment of principal cells. Second, they do not target other interneuron types or each other, but instead, mutually interact with principal cells^14–16^. Despite the hypothesized importance of their distinct anatomical features, knowledge about AAC firing patterns in the behaving animal and their role in circuit control is limited^3,14,17,18^. Spiking activity of AACs has been mainly examined in connection with hippocampal sharp wave ripples (SPW-R), a network event critical in memory selection and consolidation, but their specific role has remained controversial^3,17–20^.
Leveraging the recent availability of the Unc-5b-Cre mouse-line^14^, which enables genetic access to AACs, we conducted in vivo silicon probe recordings to characterize the spiking activity of CA1 AACs in SPW-R. We report that participation of AACs in SPW-R falls along a continuum, driven by diverse interactions with CA1 pyramidal cells. Using optogenetic inhibition of AACs we found that AACs exhibit control over CA1 circuitry, most strongly during SPW-R. SPW-R that occurred during AAC-OFF epochs were of longer duration and higher power, though unchanged in frequency. Similarly, during AAC-OFF SPW-R, CA1 pyramidal cell recruitment was enhanced, suggesting that the magnitude of circuit activation during SPW-R, and potentially cell assembly selection, is controlled by AACs. These results demonstrate a critical role for AACs in CA1 SPW-R.
Results
To characterize the activity of AACs, we recorded LFP and ensemble spiking in the hippocampal CA1 region of awake mice expressing excitatory or inhibitory opsins in AACs (Fig. 1A). Selective opsin expression was achieved by injecting AAV2/5 encoding Cre-dependent opsins into the CA1 layer of adult Unc5B-CreER mice^14^, allowing us to opto-tag AACs. AACs were identified by activation or inactivation (ChR2 or Arch expressing AACs; Fig. 1B). We identified cellular-synaptic properties of 43 AACs and examined how their reciprocal interactions with CA1 pyramidal cells organize circuit activity during SPW-R.
Characteristics of CA1 axo-axonic cells
We quantified physiological metrics of AACs (n = 43 AACs, n = 7 animals, 17 sessions). The somata of AACs were observed mostly in or close to the pyramidal layer (Fig. 1C, μ = 4.33μm relative to center of pyramidal layer, σ = 212.4) in agreement with previous studies^14,17,19^. Spike width was variable, (Fig. 1D) with a trough to peak latency range of 0.18–0.68ms (Fig. 1F, μ = 0.35ms, σ = 0.14), as were spike autocorrelations (Fig. 1E). Mean firing rates were observed from 0.18–32.47hz (Fig. 1G, μ = 11.14hz, σ = 8.62), with inter-spike intervals (ISI) ranging between 0.41ms to 285.71ms (Fig. 1H, μ = 45.45ms, σ = 55.95). A small group of AACs showed transiently fast spiking (“bursting”), while the population had an observed range of 0.0036–0.6621 (burst index is fraction of spikes with less than 6-ms ISI^21^; Fig. 1I, μ = 0.1747, σ = 0.1906). This unexpected spectrum of physiology is interesting as it stands in contrast to the notion that AACs are monolithic, rather they exhibit significant intra-class variability as suggested in Varga et al., 2014^17^.
Axo-axonic cells preferentially spike at the descending phase of the local theta oscillation
Prior reports have shown strong modulation of AAC spiking by phase of the ongoing theta rhythm^3,17^. Here we report in the largest to date collection of AACs (n = 43). We found that many AACs were strongly theta modulated, similar to many CA1 interneurons. Theta phase preference of six AACs from a single recording session are shown in Fig. 1J, demonstrating the observed variability is not due to inter-animal differences. The majority of AACs exhibit significant theta phase modulation (30/32, Supplementary Table 1, Rayleigh z test). Theta phase preference and phase modulation were correlated (Fig. 1K, rho = 0.7872, p = 9.1922e-5, Circular-linear correlation coefficient), and most AACs spikes were distributed on the descending phase of theta(Fig. 1L). The average theta phase preference for the population was 110.28°, while the most theta modulated AAC exhibited a phase preference of 91.04°. AACs as a population are highly theta modulated cells, with a phase preference for the descending phase of the oscillation^3,17,18^.
Axo-axonic cells are activated in SPW-R
The participation of AACs in CA1 SPW-R is critical to understanding their role in the CA1 circuit, though there are mixed reports as to their level of participation^3,14,17–20^. Ripple-associated spiking activity of 43 AACs (Fig. 2A, n = 43 cells, n = 595 – 16,058, μ = 5,666.4 SPW-R per recording session), revealed diverse participation across the population. On average AACs have significantly increased firing rate during SPWR (Fig. 2B, Left: n = 43 cells, baseline , SPW-R , p = 3.5132e-4, Wilcoxon signed-rank test). Interestingly, out of the 40 AACs that changed their firing rate (ZETA^22^), 27 cells increased and 13 cells decreased their activity (Fig. 2B, Right), which stands in contrast to other types of interneurons with a more uniform response, such as PV + cells (Fig S1, 43/46 cells showed increased activity). AACs demonstrated strong ripple phase modulation with most cells firing on the ascending phase of the ripple oscillation (100–250Hz, Fig. 2C, θμ = 255.02 degrees), similar to previous reports for PV + cells.
Single cell ripple modulation values (SPW-R firing rate gain) ranged from .61–3.01 (Fig. 2D, μ = 1.24, σ = 0.46). The average number of spikes in SPW-R ranged from 1.02–4.73 (Fig. 2E, μ = 2.02, σ = 1.02), while inter-spike intervals of AACs in SPW-R ranged from 9.1–37.2ms, approximating a range of 1–5 cycles of the oscillation (Fig. 2F, μ = 20.3ms, σ = 7.0). Additionally, because SPW-R information content systematically varies from the beginning to end of each event, we calculated the average time of the first spike from the peak of the SPW-R, which ranged from − 21.7–42.0ms (Fig. 2G, μ=−7.5ms, σ = 0.0064), suggesting AACs influence activity throughout the SPW-R event. Together these findings demonstrate that AAC activity in SPW-Rs is variable, however most AACs increase their activity in SPW-R events.
CA1 pyramidal neurons control the participation of axo-axonic cells in SPW-R
To determine if local excitation is a mechanism by which AAC activity is modulated in SPW-R, we quantified excitatory synaptic inputs from local CA1 PYR. Such monosynaptic connections were identified using a validated monosynaptic inference method^23,24^ (based on cross-correlations of putatively-classified pyramidal cells and AACs). For AACs with at least one presynaptic PYR, we calculated the average SPW-R modulation of the presynaptic inputs. Example SPW-R cross-correlations (Fig. 3A) illustrate both strongly and weakly modulated AACs and their partners (note that an AAC with a large peak in the cross-correlation at the time of SPW-R has partners that also show a large peak, Fig. 3A, top). AAC ripple modulation correlated with that of its inputs, such that AACs that received synaptic drive from ripple modulated PYR were themselves strongly ripple modulated. (Fig. 3B, r = 0.6923, p = 1.3325e-4, Spearman’s Rank Correlation; correlation with randomly selected non-presynaptic partner control, r = 0.1915 p = 0.3574, Spearman’s Rank Correlation; see Fig S2 for correlations with other physiological metrics.). We found further evidence that the CA1 PYR modulate AAC activity in SPW-R when examining other metrics. We quantified the correlation between spiking activity and power in the ripple frequency band as well as the magnitude of phase modulation. There was a significant relationship between spike rate power correlation of presynaptic PYR and AACs (Fig. 3C, r = 0.4619, p = 0.0185, Spearman’s Rank Correlation), as well as ripple phase modulation between paired cells (Fig. 3D r = 0.5015 p = 0.0099, Spearman’s Rank Correlation). Given the relationship between presynaptic and AAC activity, we investigated the relationship between strength of presynaptic connections and AAC participation in SPW-R. Both AAC SPW-R modulation and time to first spike were related to spike transmission probability. Stronger spike transmission probability was associated with higher ripple modulation (Fig. 3E, r = 0.5508, p = 0.0041, Spearman’s Rank Correlation), and a shorter delay between first spike and SPW-R peak (Fig. 3F, r=−0.7039, p = 9.2163e-5, Spearman’s Rank Correlation). These results suggest that excitation from CA1 PYR controls AAC activity during SPW-R.
AACs control network-wide spiking activity and SPW-R dynamics
We found that AACs robustly participate in SPW-R, though whether they influence the generation of SPW-R in the network remains unknown. To address this question, we optogenetically inhibited local AACs expressing archaerhodopsin (Arch). Silencing AACs increased power in the ripple band (100–250hz, Fig. 4A,B, Bottom: n = 5 sessions, 4 mice. n = 843–1,936 300ms stims. p = 0.0020, Paired T-test), as well as power in the 300–500Hz band, which reflects population spiking activity (300–500hz, Fig. 4A,B, Top: n = 5 sessions, 4 mice. n = 843–1,936 300ms stims. P = 3.0224e-04, Paired T-test). We then detected SPW-R and examined properties of such events occurring either in or out of AAC inhibition epochs. Subtraction of baseline ripple PSDs from AAC-OFF ripple PSDs reveal increased power of the ripple frequency at the time of these events. (Fig. 4C, n = 5 sessions, 4 mice. n = 1,495 – 12,364 baseline ripples, n = 64–335 stim ripples), as did event durations (Fig. 4E, n = 5 sessions, 4 mice. n = 37,032 baseline ripples, n = 1,024 stim ripples. p = 0.0072, Wilcoxon rank sum-test), and peak amplitude (Fig. 4F, n = 5 sessions, 4 mice. n = 37,032 baseline ripples, n = 1,024 stim ripples. p = 2.0593e-06, Wilcoxon rank sum test), while ripple frequency was not affected (Fig. 4D, n = 5 sessions, 4 mice. n = 37,032 baseline ripples, n = 1,024 stim ripples. p = 0.8675, Wilcoxon rank-sum test). This suggests that AACs control content and magnitude of SPW-R while other cell types may pace the rhythm (likely basket cells^25^). These data reveal an important role for AACs in controlling SPW-R dynamics at the network level.
AACs control pyramidal cell participation in SPW-R
Pyramidal cells are reliably depolarized during SPW-R, and competition between excitatory and inhibitory inputs compete to determine spiking at the single cell level^12^. Given our findings that most AACs are active in SPW-R, and inhibiting them alters SPW-R dynamics, we next investigated the effects of silencing AACs on PYR spiking. We first looked at the effects of manipulating AAC activity in all states. AAC activation inhibited PYR firing, while AAC silencing resulted in PYR disinhibition, revealed by changes in their firing rates. (Fig. 5A, Left, ChR2: n = 11 sessions, 4 mice. n = 1,063 – 4,459 100ms stims, n = 180 PYR, Right, Arch: n = 5 sessions, 4 mice. n = 843–1,936 300ms stims, n = 207 PYR). These results demonstrate that AACs tonically inhibit PYR. Next, we sought to investigate how AAC inhibition shapes PYR activity selectively during SPW-R by comparing baseline ripples to ripples that occurred during periods where AACs were silenced. We quantified the fraction of PYR that participated in AAC-OFF SPW-R compared to baseline. In AAC-OFF SPW-R, we observed a significant increase in PYR participation (Fig. 5B, Left: AAC: n = 5 sessions, 4 mice. n = 31–55, total: 194 PYR. n = 1,495 – 12,364, total: 37,032 Baseline SPW-R. n = 51–287, total: 868 AAC-OFF SPW-R. p = 5.2716e-16, Wilcoxon rank-sum test). Superficial and deep CA1 PYR receive different patterns of inhibition^26,27^, thus we quantified this effect separately for each subgroup of pyramidal population. In AAC-OFF SPW-R, the fraction of both superficial and deep PYR participation increased (Fig S3, Right: Top, AAC, n = 1,495 – 12,364, total: 37,032 Baseline SPW-R. n = 51–287, total: 868 AAC-OFF SPW-R. n = 11–32, total: 125 Deep cells, n = 5–33, total: 69 Superficial cells. Deep: p = 2.4645e-13, Wilcoxon rank-sum test. Superficial: p = 6.5141e-06, Wilcoxon rank-sum test.). Given that cell assemblies constructed from unique sets of PYR enable the encoding of separate experiences by SPW-R^28–31^ we examined SPW-R associated PYR activity at an individual cell level. On average, PYR exhibited a higher count of spikes in AAC-OFF SPW-R (Fig. 5C, n = 5 sessions, 4 mice. n = 194 PYR, n = 1,495 – 12,364 Baseline SPW-R. n = 51–287 AAC-OFF SPW-R. p = 6.1155e-08, Wilcoxon signed-rank test). Similarly, pyramidal cell SPW-R firing rate gain was significantly higher when AACs were silenced (Fig. 5D, n = 5 sessions, 4 mice. n = 194 PYR, n = 1,495 – 12,364 Baseline SPW-R. n = 51–287 AAC-OFF SPW-R. p = 2.4569e-08, Wilcoxon signed-rank test). Increased firing rate was naturally reflected in decreased inter-spike interval in AAC-OFF SPW-R. (Fig. 5E, n = 5 sessions, 4 mice. n = 194 PYR, n = 1,495 – 12,364 Baseline SPW-R. n = 51–287 AAC-OFF SPW-R. p = 6.9834e-05, Wilcoxon signed-rank test.) Further, AACs participated in a higher fraction of total AAC-OFF SPW-R than baseline SPW-R. (Fig. 5F, n = 5 sessions, 4 mice. n = 194 PYR, n = 1,495 – 12,364 Baseline SPW-R. n = 51–287 AAC-OFF SPW-R. p = 1.8104e-11, Wilcoxon signed-rank test). These results demonstrate AACs control the participation of pyramidal cells in SPW-R events, likely contributing to functions such as replay.
Discussion
Glutamatergic excitation and GABAergic inhibition compete to control PYR participation in SPW-R^25^. In each SPW-R, only a minority of PYR spike even though Vm is depolarized above spike threshold for 10’s of milliseconds, due to dynamically balanced GABAergic inhibition. Inhibition thus plays a critical role in enabling competition-based selection and expression of PYR assemblies. Identifying the source(s) of this inhibition is critical to understanding the CA1 circuit, and likely arises from axo-somatic targeting cells such as basket cells and AACs. Here we identify novel mechanisms by which AACs participate in and control CA1 SPW-R. Notably, these findings strongly suggest that reciprocal interactions between excitatory PYR and inhibitory AACs organize cell assembly construction and overall network activity during SPW-R. This places AACs in a critical position in determining the content of CA1 information delivered to neocortex.
Recruitment of AACs to sharp wave-ripples is explained by monosynaptic inputs from CA1 PYR
CA1 interneuron spiking is controlled both by afferent inputs (e.g., CA3) and excitation and inhibition from local PYR and INT, respectively^32–34^. To determine the activity of AACs and PYR during SPW-R at single-spike resolution, we used silicon probes to obtain CA1 ensemble recordings and opto-tagged AACs. As a population, AAC activity is significantly increased during SPW-R, with a minority of cells exhibiting decreased activity, emphasizing the importance of considering intra-class variability in studying cell types^17^ (Fig. 2A,B). In agreement with these findings, up- and down-modulated AACs have been previously described in studies using both juxta-cellular electrophysiology^26^ and calcium imaging^14,19^. To better understand the driver of AAC spiking in SPW-R, we examined their physiology and excitatory inputs from CA1 PYR. Individual AAC activity in SPW-R is strongly correlated with the activity of presynaptic partners and the strength of these connections (Fig. 3), suggesting that the local circuit can control AAC activity independently of afferent input. An important future question is if the propensity for AACs to participate in SPW-R is consistent across experiences and/or throughout the lifespan of the animal^35^. Such plasticity could be supported both by changes in excitatory inputs we identified as well as inhibition from local interneurons^36^. Longitudinal recording of single AACs would be required to answer these questions.
AACs control network activation and PYR spiking in sharp wave-ripples
Silencing AACs led to increased power in both the spiking band and SPW-R band (Fig. 4A,B), SPW-R events that occurred during AAC silencing were of longer duration, and had higher power and peak amplitude (Fig. 4C,F,G). Interestingly AAC silencing did not alter SPW-R frequency. These data support a model in which AACs robustly control PYR recruitment, while other interneurons such as PV + cells may be more important for synchronizing PYR populations at ripple frequency^25^.
PYR activity in SPW-R was affected significantly by silencing AACs. Inhibition of AACs increased the number of participating PYR (Fig. 5B) and produced increased average spike count, firing rate gain and decreased inter-spike intervals (Fig. 5C,D,E). Additionally, PYR participated in a greater fraction of AAC-OFF epochs than during control SPW-R, suggesting AACs function to select which PYR are allowed to participate in each event. Our findings that during SPW-R, silencing AACs disinhibits PYR, and that PYR drive AAC spiking, suggests lateral inhibition is instantiated by reciprocal interactions between pyramidal cells and AACs. These results support the general hypothesis that individual interneurons serve unique functions in CA1 circuit computations, and that this mechanism supports the construction of and competition between cell assemblies. Our experiments demonstrate this specifically for the understudied AAC interneuron family.
Methods
All protocols and experiments using animals were approved by the Virginia Tech (Blacksburg, VA, USA) Institutional Animal Care and Use Committee (IACUC). The Unc5b-CreER mice were obtained from Z. Josh Huang under a fully executed MTA, and either maintained on a homozygous C57BL/6J background or were bred with FVBN/J wild-type mice to generate heterozygous mice. Male and female adult mice (> 12 weeks) were used in all experiments.
AAV Injection
Mice are induced and maintained at a surgical plane of anesthesia with isoflurane while mounted in a stereotaxic frame. Hair is removed and scalp is disinfected. Bupivicaine is injected once under the scalp before revealing skull. A < 0.2 mm burr hole is made above area CA1 of the hippocampus in both hemispheres. (mm from bregma: −1.8, lateral: +/−1.5). Glass pipette containing AAV5-EF1a-DIO-hChR2 (H134R)-EYFP or AAV5-EF1a-DIO-eArch3.0-EYFP (UNC Gene Therapy Center – Vector Core) is lowered into CA1 using stereotaxic coordinates. 100nL of AAV (titer: 3.8–5.2×10^12^ GC/mL) is injected into tissue (1nLs^−1^) using a nanoliter injector (WPI: MICRO2T & 504127). Following infusion, the glass pipette is left to rest for 5 minutes before slowly removing from the brain. The scalp is then closed with Vetbond (3M).
Tamoxifen injections
Mice were injected intraperitoneally with tamoxifen (100 mg/kg) dissolved in corn oil at 20 mg/ml on days 5,7, and 9 following AAV injection.
Head-bar Implantation
Mice are induced and maintained at a surgical plane of anesthesia with isoflurane while mounted in a stereotaxic mount. Bupivicaine is injected under the scalp before it is removed. The skull is cleaned with 3% hydrogen peroxide, followed by an application of dental adhesive Optibond (Kerr Dental). A small burr hole is made above cerebellum, and a stainless-steel ground wire is inserted between the skull and the brain, parallel to the brain surface, then the wire is affixed to the skull with sterile dental acrylic. A headplate is positioned parallel to the skull above lambda, and is permanently fixed in place.
Craniotomy
One day before recording, mice are induced and maintained at a surgical plane of anesthesia with isoflurane while mounted in the stereotaxic mount. A < 1.0 mm burr hole is made above the hippocampus and the dura is removed. Kwik-cast (World Precision Instruments) is applied to the burr hole, with dental acrylic applied on top, to protect the craniotomy until recording.
In Vivo recording
Unc5b-CreER mice were obtained from the lab of Z. Josh Huang under full MTA. All animals used in experiments were homozygous Unc5b-CreER or hybrid Unc5b-CreER × FVB/NJ (JAX #001800) mice. Additionally, PV-Cre mice were acquired from the Jackson Laboratory (Jax #017320). During the time of experiments, mice were between the ages of 16–30 weeks old. Mice of both sexes were used in all experiments. Mice, trained and habituated to head-fixed navigation, are placed in the head fixation apparatus, and a silicone probe (Poly3, Neuronexus, or P1, Cambridge Neurotech, with optic fiber attached, or 4×8 uLED, Neurolight technologies), is lowered through the craniotomy into the brain until SPW-R are observed and left in place for 30–45 minutes until tissue is relaxed. The neural signals were amplified, digitized and recorded with RHD2000 system (Intan Technologies LLC).
Optical Stimulation
Optical pulses were delivered through a sharpened optic fiber connected to a Thorlabs laser diode driver and laser diode or optical pulses are delivered through LEDs in a 4×8 uLED probe (Neurolight Technologies).
Data collection
Unc5b-CreER ChR2, Arch, and PV + data was collected for this study. A selection of PV + data was used with permission from the Buzsaki Data Repository^24,37^.
Data analysis
Data analysis was conducted in Matlab (Mathworks) and custom scripts were used to analyze the extracellular and spike data. Spike sorting was performed using Kilosort1^38^ with custom semi-automated reclustering^39^ followed by manual curation using Phy. Cell Explorer^40^ was used to characterize neuron properties including cell position, autocorrelation, wave form characteristics, firing rate characteristics, putative monosynaptic connections (followed by manual refinement) and SPW-R modulation. The Buzcode repository was used to calculate cross correlations and for SPW-R detection. The HippoCookBook repository and Circular Statistics Toolbox (Mathworks) were used to quantify oscillation phase preference and phase modulation. Rate power correlation was calculated by correlating spike trains with power in the SPW-R band (100–250Hz). Opto-tagged AACs were identified using CellExplorer to classify putative cell type, and Zenith of Event-based Time-locked Anomalies (ZETA^22^) to determine significance of the response to stimulation. Further, opto-tagged AACs were manually refined through observation of peri-stimulation time histogram to optical stimulation. Significance of SPW-R modulation was determined using CellExplorer and ZETA. For detection of SPW-R characteristics in and out of stimulation (Fig. 4), SPW-R were classified as having the peak of the event within stimulation times. For detection of spikes from cells in SPW-R events (Fig. 5), only SPW-R that had the entire duration within a stimulation pulse were considered INT-OFF SPW-R.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Bugeon S (2022) A transcriptomic axis predicts state modulation of cortical interneurons. Nature 607:330–33835794483 10.1038/s 41586-022-04915-7PMC 9279161 · doi ↗ · pubmed ↗
- 2Harris KD (2018) Classes and continua of hippocampal CA 1 inhibitory neurons revealed by single-cell transcriptomics. P Lo S Biol 16:e 200638729912866 10.1371/journal.pbio.2006387 PMC 6029811 · doi ↗ · pubmed ↗
- 3Klausberger T (2003) Brain-state- and cell-type-specific firing of hippocampal interneurons in vivo. Nature 421:844–84812594513 10.1038/nature 01374 · doi ↗ · pubmed ↗
- 4Klausberger T, Somogyi P (2008) Neuronal Diversity and Temporal Dynamics: The Unity of Hippocampal Circuit Operations. Science 321:53–5718599766 10.1126/science.1149381 PMC 4487503 · doi ↗ · pubmed ↗
- 5Lapray D (2012) Behavior-dependent specialization of identified hippocampal interneurons. Nat Neurosci 15:1265–127122864613 10.1038/nn.3176 PMC 3433735 · doi ↗ · pubmed ↗
- 6Buzsáki G, Chrobak JJ (1995) Temporal structure in spatially organized neuronal ensembles: a role for interneuronal networks. Curr Opin Neurobiol 5:504–5107488853 10.1016/0959-4388(95)80012-3 · doi ↗ · pubmed ↗
- 7Cobb SR, Buhl EH, Halasy K, Paulsen O, Somogyi P (1995) Synchronization of neuronal activity in hippocampus by individual GAB Aergic interneurons. Nature 378:75–787477292 10.1038/378075 a 0 · doi ↗ · pubmed ↗
- 8Tremblay R, Lee S, Rudy B (2016) GAB Aergic Interneurons in the Neocortex: From Cellular Properties to Circuits. Neuron 91:260–29227477017 10.1016/j.neuron.2016.06.033PMC 4980915 · doi ↗ · pubmed ↗