Preparing high-quality chromosome spreads from Crocus species for karyotyping and FISH
Abdullah El-nagish, Susan Liedtke, Sarah Breitenbach, Tony Heitkam

TL;DR
This paper presents an effective method for preparing high-quality chromosomes in saffron and related crocus species, improving FISH analysis for genetic studies.
Contribution
The study introduces ice water pretreatment as a novel and effective protocol for chromosome preparation in Crocus species.
Findings
Ice water pretreatment yielded the highest metaphase index (2.05%) compared to other methods.
Chromosomes prepared with ice water showed better morphology and suitability for FISH analysis.
DAPI staining and FISH confirmed the effectiveness of ice water pretreatment in both C. sativus and C. cartwrightianus.
Abstract
The saffron-producing Crocus sativus (L.) and its wild relative C. cartwrightianus (Herb.) are key species for understanding genetic evolution in this genus. Molecular-cytogenetic methods, especially fluorescent in situ hybridization (FISH), are essential for exploring the genetic relationships in this genus. Yet, preparing high-quality chromosomes for FISH analysis across Crocus species remains difficult. A standardized protocol for achieving clear and well-separated mitotic chromosomes is still lacking. This study aimed to assess the effectiveness of pretreatments with four chromosome synchronization methods for optimal chromosome spread preparation in Crocus. Root tips of different Crocus species were treated with four chromosome preparation methods namely hydroxyurea-colchicine (HC), nitrous oxide (NO), hydroxyquinoline (HQ), and ice water (IW) pretreatments to investigate their…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
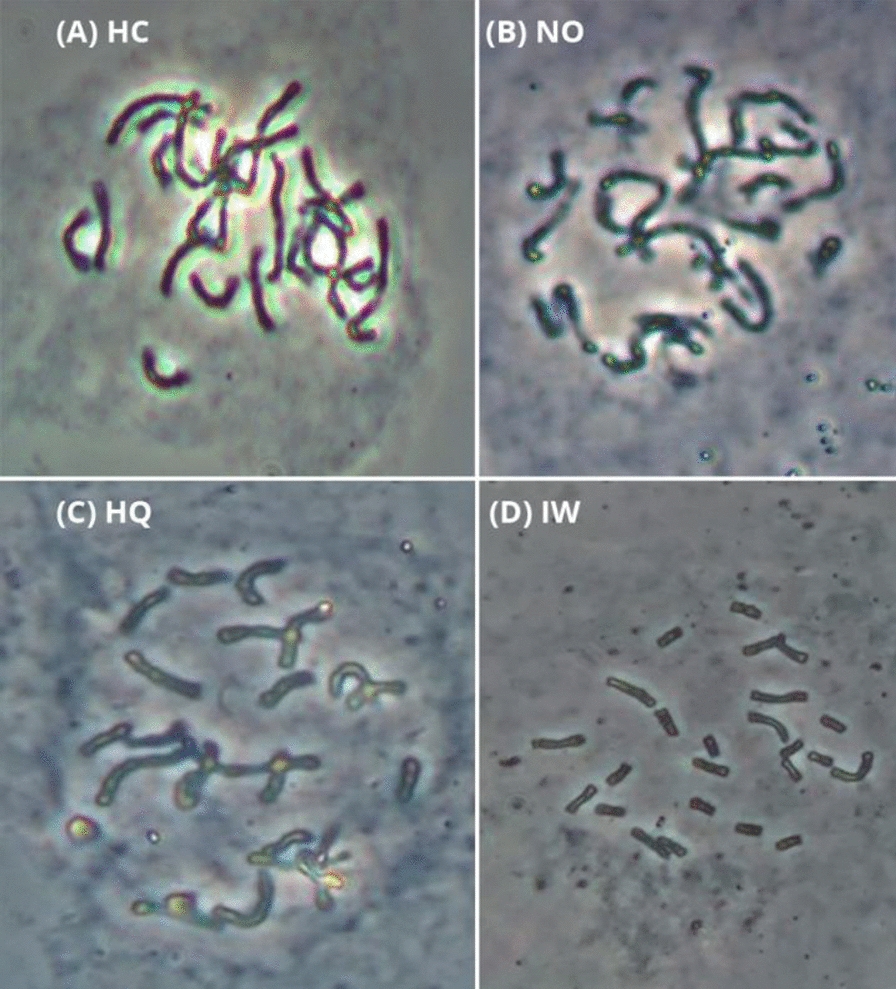 Figure 1
Figure 1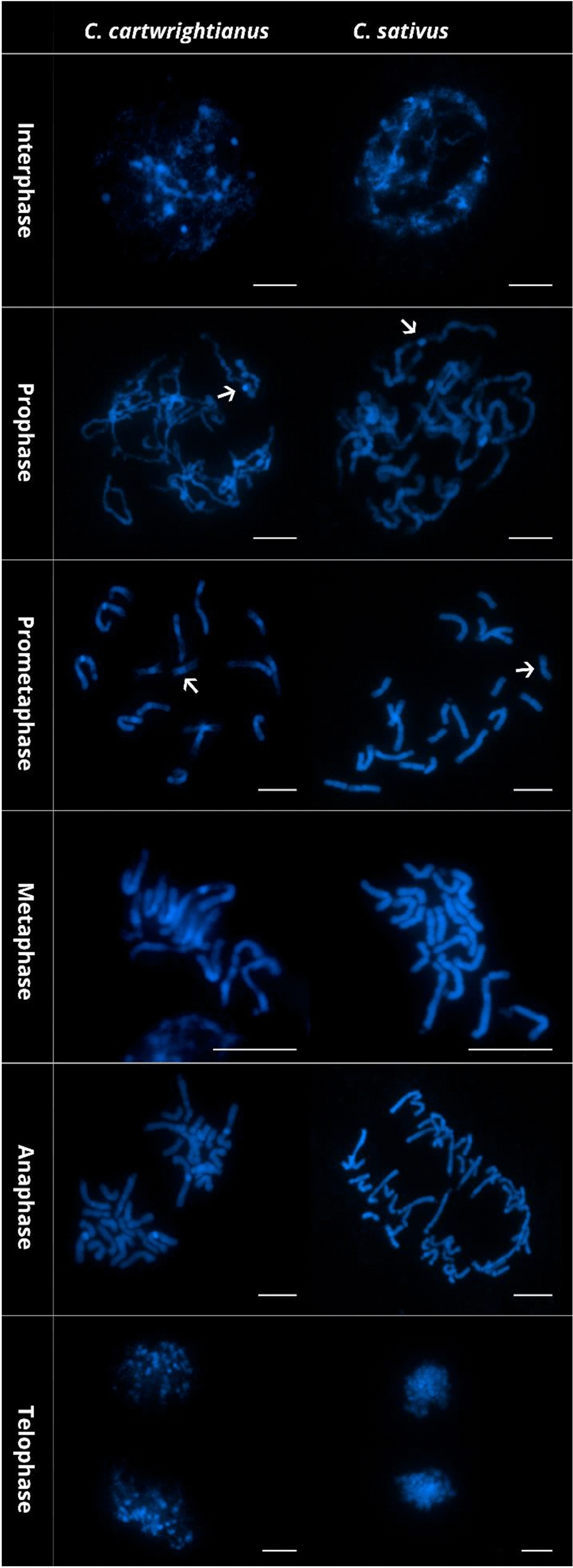 Figure 2
Figure 2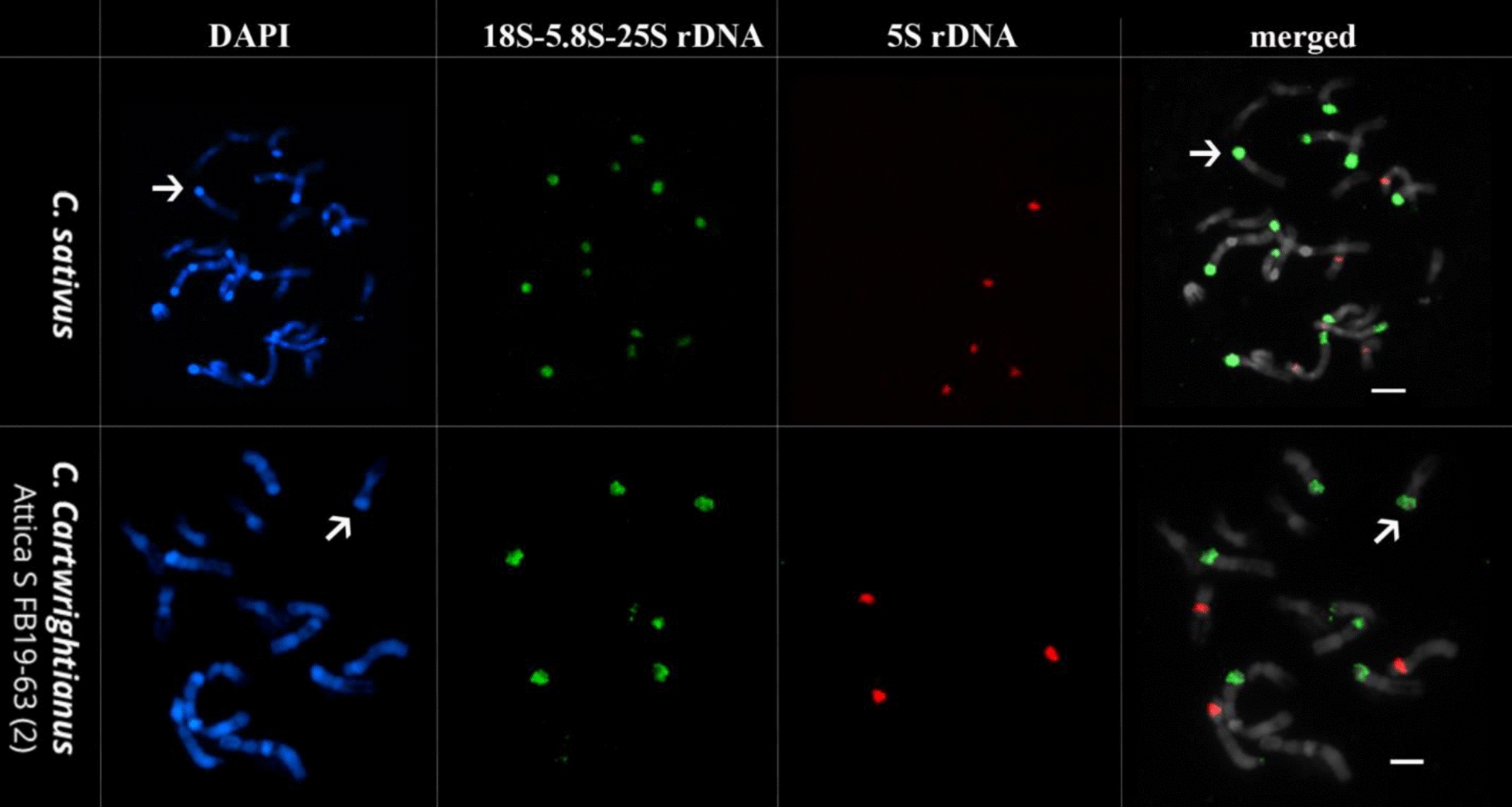 Figure 3
Figure 3- —https://doi.org/10.13039/501100002385Ministry of Higher Education
- —https://doi.org/10.13039/501100001659Deutsche Forschungsgemeinschaft
- —RWTH Aachen University (3131)
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsSaffron Plant Research Studies · Chromosomal and Genetic Variations · Plant Reproductive Biology
Background
The genus Crocus (Iridaceae) comprises around 250 species widely distributed over a wide range of climatic areas [1, 2] and is known for its variable chromosome numbers [3–5]. Among the species of Crocus, only C. sativus is used as a crop and thus receives the most attention. C. sativus is the source of saffron, one of the highest priced spices of the world, processed from dried stigmas of manually harvested flowers. It is a cash crop for agriculture-based communities living off marginal areas in Iran, North Africa, countries surrounding the Mediterranean basin, and Kashmir. Despite its economic relevance, we are just beginning to understand the genomic and chromosomal constitution of saffron crocus and related species.
C. sativus is a male-sterile triploid species harboring eight chromosome triplets (2n = 3x = 24) and having a genome size of 1C = 3.45 Gbp [6]. Due to its triploidy, saffron crocuses can only be propagated vegetatively. As all saffron accessions around the globe have a similar genome, it is generally accepted that triploid saffron emerged only once [3, 7–11]. We and other groups recently showed that cytotypes of Crocus cartwrightianus have been the sole precursors of saffron’s triploidy, and that its emergence can be traced to the Aegean Bronze Age in Greece [3, 4, 12, 13]. C. cartwrightianus is a diploid species (2n = 16) with high genetic diversity [3, 4, 12]. Nevertheless, despite these recent insights into the origin of saffron crocus, the C. cartwrightianus cytotypes that may enable targeted re-breeding and improvement of saffron traits have not yet been identified. Similarly, the origin of the individual chromosomes within saffron’s chromosome triplets is still unclear, especially as one chromosome is heteromorphic [14]. However, genomic and cytogenetic analyses in the genus Crocus may provide detailed insights into the chromosome structure of Crocus species, but robust and widely applicable protocols are still lacking.
Fluorescent in situ hybridization (FISH) is a powerful cytogenetic technique to study structure and function of chromosomes, polyploidy and genome evolution. In particular, the physical mapping of tandemly repeated DNA sequences provides informative cytogenetic landmarks for unequivocal chromosome identification in many plant species [15–19]. The first in situ hybridizations along Crocus chromosomes already showed the potential of repeat probes in this genus, yielding a range of distinct signals and allowing first chromosome assignments [20–22]. Recently, we developed a karyotyping mix for Crocus species that is composed of six tandem repeat probes [4] and that opens the Crocus genus for comprehensive molecular-cytogenetic analyses to clarify the genetic details of saffron crocus’ ancestry. However, streamlining FISH analysis across a range of species, cytoypes and accessions requires a robust protocol for properly dispersed mitotic chromosomes for its application. Despite being studied cytogenetically for several decades, species of the Crocus genus still remain challenging targets for chromosome preparation. Usually, due to the strict annual growth and the small size of the plant, material is limited, especially of wild accessions. In addition, as the chromosomes are large, some preparation techniques such as dropping [23–26] are not recommended. Therefore, a comparative study testing different methods to obtain high-quality mitotic chromosomes from Crocus species is needed.
Here, using the crop plant C. sativus for its high economic value and C. cartwrightianus for its scientific interest, we compared the effectiveness of four chromosome fixation methods for obtaining Crocus chromosome spreads. We use (1) the hydroxyurea-colchicine method, (2) the nitrous oxide method, (3) the hydroxyquinoline method, and (4) ice water treatments and analyze them for their yield in obtaining properly spread mitotic chromosomes and mitotic index.
Methods
Plant materials and time of harvest
We used root tips of C. sativus (corms collected commercially), C. cartwrightianus HKEP 1517 and C. cartwrightianus (Attica S FB19-63 (2)) was provided by F. Blattner and D. Harpke (IPK Gatersleben, Gatersleben, Germany). All plants were grown under glasshouse conditions. Root tips were collected in the early morning hours (07.00–08.00).
Experimental design
Root tips of C. sativus were subjected to four different pretreatments techniques. Each was repeated six times:
- Hydroxyurea-colchicine method (HC) Corms with roots were incubated for 18 h in liquid, 0.5 × LM medium containing 1.25 mM hydroxyurea. The material was kept in 125 mL Erlenmeyer flasks placed on an orbital shaker at 150 rpm at room temperature. After three rinses with 0.5 × LM medium without hydroxyurea, the material was incubated for 6 h in fresh medium followed by treatment with the medium containing 0.6% (w/v) colchicine for 20 h [27]. Treated root tips were excised and fixed in a 3:1 (v/v) ethanol: acetic acid solution for 24 h at 4 °C.
- Nitrous Oxide (NO) Root tips were incubated in a pressure-tolerant cylinder, with nitrous oxide gas applied for 45 min at room temperature at 10 bar [28]. After this, the root tips were fixed in a 3:1 (v/v) methanol: acetic acid solution for 24 h at 4 °C [29].
- Hydroxyquinoline (HQ) Roots from individual corms were collected, pretreated with 2 mM hydroxyquinoline for 5 h and fixed in a 3:1 methanol: acetic acid [4].
- Ice water (IW) Corms with roots were placed in container filled with ice water, kept inside a refrigerator at 4∘ C for 18 h. Roots of 2–3 cm long were cut from and were fixed in methanol: acetic acid (3: 1) for 2 h at 4 °C, fresh fixative was add and kept for 24 h at 4 °C.
Protocol of chromosome preparation from Crocus material
Enzyme treatment of root tips
- Wash root tips 1 × in dist. H_2_O for 5–10 min, 2 × in 4 mM citrate buffer (4 mM citric acid and 6 mM sodium citrate), pH 4.5 for 5 min each.
- Dissect meristematic tips using a sharp scalpel and transfer them into a petri dish with 20–30 μl of enzyme mixture. The enzyme solution consists of 2% (w/v) cellulase from Aspergillus niger, 4% (w/v) cellulase ‘Onozuka R10′ from A. niger, 2% (w/v) hemicellulase from A. niger, 0.5% (w/v) pectolyase from Aspergillus japonicus and 20% (v/v) pectinase from A. niger in citrate buffer.
- Incubate at 37 °C for 2.0–2.5 h depending on species (Table 1).
Table 1. Incubation times of different Crocus root tips for all chromosome preparation protocolsSpecies/AccessionIncubation time in enzyme mixtureIncubation time on hot plate C. sativus 2.30 h60 sC. cartwrightianus HKEP 15172.00 h30 sC. cartwrightianus (Attica S FB19-63 (2))2.15 h30 s
Spread preparation
- Single root tips were transferred onto slides, macerated in 50–60 μl of 45% acetic acid with needles for 120 sec. “Methods”. Add an extra drop of 45% acetic acid, mix with a needle, spread on a hot surface at 55 °C for 30–60 s depending on species (Table 1).
Spread fixation
- Surround with drops of freshly prepared fixative (3:1 methanol:acetic acid), dropwise fresh fixative on top of the slide as well.
- Let the fixative run down, rinse with more fixative.
- Air-dry the slides.
- Store slides in a Coplin jar in a freezer until usage.
Assessing the quality of the mitotic chromosome spreads via phase contrast and fluorescent microscopy
The chromosome preparations were assessed via phase contrast and fluorescent microscopy. For the latter, mitotic chromosome spreads were prepared according to the protocol above. Slides were equilibrated in 4 × SSC/0.2% Tween 20 for 5 min at 37 °C. Excess liquid was carefully removed, then, 20 μl DAPI-Citifluor AF1 were added on each slide and covered with glass cover.
Wide-field imaging was performed with a Zeiss Axioimager M1 UV epifluorescence microscope with appropriate filters, and equipped with an ASI BV300-20A camera coupled with the Applied Spectral Imaging software (Applied Spectral Imaging, Carlsbad, CA, USA). The images were processed with Adobe Photoshop CS5 software (Adobe Systems, San Jose, CA, USA) using only contrast optimization, Gaussian and channel overlay functions affecting the whole image equally.
The quality of the mitotic chromosome spreads was evaluated based on chromosome morphology, the absence of overlapping chromosomes, and the clarity of the spread from any debris on the slide. Chromosome spreads were considered high quality if they exhibited well-delineated, intact chromosomes with minimal background noise, which allowed for easy and accurate identification of individual chromosomes eventually facilitating accurate FISH signal detection. Metaphase index was calculated as the percentage of cells at metaphase stage. Based on the quality of the chromosome preparations and the highest mitotic indices, the best protocol was selected and tested for suitability for wild Crocus accessions, using two C. cartwrightianus accessions as use cases.
Probe labeling and fluorescent in situ hybridization
The probe “18SrRNAgene_Bv_probe1” [30] was used for the detection of the rDNA and was labeled with biotin-11-dUTP (Dyomics) by PCR and detected by Streptavidin-Cy3 (Sigma-Aldrich). The probe pXV1 [31] for the 5S rRNA gene was labeled with digoxygenin-11-dUTP by PCR and detected by anti-digoxigenin–fluorescein isothiocyanate (FITC; both from Roche Diagnostics). The hybridization procedure was performed as described previously [31]. Chromosomes were counterstained with DAPI (Honeywell, Charlotte, NC, USA). Prior to FISH, according to the amount of cytoplasm visible under light microscope, we pre-treated the slides with 100 μg/ ml RNase in 2 × SSC for 30 min, followed by 200 μl of 10 μg/ ml pepsin in 10 mM HCl for 15 to 30 min.
Results and discussion
Ice water pretreatment is most effective for obtaining metaphase spreads of high-quality in Crocus
To obtain chromosome spreads ideal for karyotyping, FISH and various other cytogenetic analysis, the use of appropriate arresting agent is required. These agents influence chromosome morphology, condensation, and the quality of chromosome spreads. The mechanisms by which these agents work include altering cytoplasmic viscosity, disrupting spindle fiber formation, and interfering with chromosome condensation [32, 33]. These effects ultimately impact the number of dividing cells and their chromosomal morphology and arrangement.
In this study, we evaluated four different pretreatments (Hydroxyurea-colchicine (HC), Nitrous Oxide (NO), Hydroxyquinoline (HQ), and Ice Water (IW)) were compared for their effectiveness in obtaining properly dispersed mitotic chromosomes of C. sativus, focusing on improving the quality of spreads and their suitability for subsequent FISH analysis. These methods were applied to root tips of C. sativus and C. cartwrightianus species. In total, 22,507 cells were evaluated and representative metaphases were selected for illustration (Fig. 1, Table 2).Fig. 1. Mitotic chromosome of C. sativus, obtained with each of the four treatments: HC (A), NO (B), HQ (C), and IW (D). The spreads shown in this figure were taken by phase contrast microscope (Zeiss Axioskop 40) at a magnification power of 400x.Table 2. Comparison of metaphase indices in four methods used for chromosome preparation from C. sativus materialsMethodNo. of cells analyzedNo. of metaphasesMetaphase index (%)Hydroxyurea-colchicine method (HC)3717401.08Nitrous oxide (NO)4714541.15Hydroxyquinoline (HQ)6400741.16Ice water (IW)76761572.05
Comparing all four methods, HC (Fig. 1A) yielded the lowest metaphase index (1.08%). HC-derived metaphases usually featured chromosomes that were difficult to count due to overlapping of chromosome arms. Hence, using HC for chromosome preparation and FISH analysis of Crocus is not recommended (Fig. 1A, Table 2). HU induces cell synchronization at the stage of DNA synthesis. Hydroxyurea is an inhibitor, acting on the ribonucleotide reductase leading to inhibition of DNA synthesis, arresting cells in the S-phase. In contrast, colchicine disrupts microtubules, preventing mitotic spindle formation and arresting cells in metaphase. Although the hydroxyurea/colchicine combination often used for cell synchronization [27, 34, 35], our results suggest that it is not suitable for Crocus, likely due to its inefficiency in chromosome condensation or factors related to concentration, exposure time, and species-specific features. Further optimization of these parameters is needed to improve HU effectiveness in Crocus.
Similar to HC, the chromosome spreads obtained by Nitrous Oxide (NO) were difficult to distinguish also due to overlapping chromosome arms resulting in limited effectiveness in producing high-quality chromosome spreads (Fig. 1B, Table 2), complicating accurate identification and analysis. NO likely interferes with mitotic spindle formation, but the lack of effective chromosome condensation reduced the resolution of the spreads. The exact cause of the low-quality chromosomal spreads with NO in Crocus is unclear. Nitrous oxide might be toxic in Crocus, causing cell cycle arrest before metaphase, as indicated by the low metaphase index. Unlike other pretreatments, NO is not known to affect spindle fiber formation; rather, it prevents proper chromosome alignment along the metaphase plate, hindering spindle fiber attachment [36].
The HQ method (Fig. 1C, Table 2) yielded comparable results, with a metaphase index of 1.16%, slightly higher than NO but still insufficient for obtaining consistently clear, non-overlapping chromosomes. HQ, known to alter cytoplasmic viscosity [32, 33, 37], likely interferes with chromosome condensation, but may not be as efficient as other agents, resulting in a lower metaphase index and suboptimal spread quality.
Weighing all four methods against each other, IW yielded the highest metaphase index (2.05%). We conclude that this pretreatment arrested the cells in metaphase twice more often than the other three methods (Table 2). Moreover, this method is preferred in terms of chromosome morphology, if the chromosomes are to be counted or analyzed by FISH procedure, as they were of preferred shape and length with no overlapping (Fig. 1D).
The increased the metaphase index and preventing overlapping of chromosome arms is evidence that the chilling stress stops DNA synthesis and mitotic activity and inhibits formation of microtubules [38, 39], leading to a better separation of single chromosomes suitable for further analyses.
Ice water pretreatment is also effective for wild crocus species and allows following the chromosomes through cell cycle
To test the most effective method also for wild Crocus species, we applied the ice water pretreatment also to C. cartwrightianus, the diploid progenitor species of C. sativus. Using IW pretreatment, chromosomes from both species, C. sativus and C. cartwrightianus, were easily stained with DAPI, resulting in clear, well-resolved chromosome spreads that allowed for detailed visualization of each stage of mitosis (Fig. 2). The preparations showed high-quality chromosome morphology, with distinct and non-overlapping chromosomes across the mitotic phases, facilitating accurate structural analysis as shown later by the FISH analysis. In the DAPI-stained images of both C. sativus and C. cartwrightianus prepared using the IW method, all stages of mitosis (interphase, prophase, prometaphase, metaphase, anaphase, and telophase) were clearly observed. Each stage displayed its characteristic features of chromosome condensation and organization (Fig. 2).Fig. 2DAPI-stained chromosome spreads of C. cartwrightianus and C. sativus. The scale bar equals 10 µm. Arrows indicate the centromeric constriction
The DAPI-staining of both C. sativus and C. cartwrightianus chromosomes enabled the identification of intercalary and terminal heterochromatin and of the mostly weakly stained centromeres, often also visible as a constriction (Fig. 2; arrowed).
Metaphases resulting from ice water pretreatment are well-suited for FISH follow-up
Chromosome spreads obtained from root tips prepared by the IW method were evaluated for their applicability for FISH analysis using 18S-5.8S-25S and 5S rDNA probes (Fig. 3).Fig. 3. Fluorescent in situ hybridization (FISH) of C. sativus and C. cartwrightianus (name given in each panel). DAPI-stained mitotic chromosomes are shown in blue. Probes used are 18S-5.8S-25S rRNA genes (green), and 5S rRNA genes (red). Arrows indicate exemplary DAPI-positive chromosome regions, which are shown to be enriched in rDNA.
As presented (Fig. 3), the chromosome spreads produced with IW were highly useful for follow-up analysis by FISH. In this example, the genetic identity can be assigned to the large terminal chromosome regions, which are strongly condensed and detectable as DAPI-positive blocks. These consist mostly of rRNA genes (Fig. 3, arrowed, Table 3). In C. sativus, we detected 12 arrays of 18S-5.8S-25S rRNA genes, including five major sites and seven moderate sites (Fig. 3, arrowed), and five major 5S rRNA gene arrays (Fig. 3, arrowed). Eight hybridization signals were detected in metaphase chromosomes of C. cartwrightianus using 18S-5.8S-25S rDNA probe, of which six were strong signals while the remaining two were weak signals. FISH analysis using 5S rDNA revealed five moderate signals in C. sativus and three strong signals in C. cartwrightianus. The number, relative signal strength and presence on chromosomes of the two rDNA sites are listed in Table 3.Table 3. Hybridization of cytogenetic landmarks in C. sativus and C. cartwrightianus using two rDNA probesTandem repeatSpeciesNumber of sitesNumber of labeled chromosomesmajormoderateminortotal18S-5.8S-25S rRNA genesC. sativus5701212C. cartwrightianus602885S rRNA genesC. sativus05055C. cartwrightianus30033Total number1412228
Conclusion
Summarizing, we present a comprehensive comparison of four methods (HC, NO, HQ, and IW) for preparing mitotic chromosome spreads in Crocus sativus. The results indicate that ice water pretreatment (IW) is more suitable, yielding the highest metaphase index (2.05%) and providing the best chromosome morphology for further analysis. In contrast, HC, NO, and HQ yielded lower metaphase indices and produced chromosomes with poor morphology, making them less suitable for cytogenetic studies in Crocus. The unsuitability of the three techniques may result from non-optimized parameters for Crocus. Future studies to optimize the parameters of these techniques for Crocus are needed. IW pretreatment was also equally well with C. cartwrightianus, the diploid wild progenitor species.
The suitability of this method for cytogenetic analysis across different Crocus species was demonstrated by a) enabling us of following the different stages of mitosis in both species with a clear identification of DAPI-stained metaphase chromosomes that provided detailed insights into chromosomal structure, b) the successful FISH hybridization to chromosomes enabling the visualization of key chromosomal features such as heterochromatin and centromeres in both cultivated and wild crocus species.
The reference list from the paper itself. Each links out to its DOI / PubMed record.