Impacts of perR on oxygen sensitivity, gene expression, and murine infection in Clostridioides difficile 630∆erm
Anna L. Gregory, Hailey E. Bussan, Madeline A. Topf, Andrew J. Hryckowian

TL;DR
Researchers studied how a mutation in the perR gene affects Clostridioides difficile's oxygen sensitivity and infection in mice.
Contribution
The study identifies transcriptomic differences due to a perR mutation and shows no impact on infection in a murine model.
Findings
Restoring PerR function in 630Δerm does not affect growth under anaerobic or low-oxygen conditions.
630Δerm perRWT is more sensitive to ambient oxygen and shows altered gene expression.
The perR mutation does not impact infection or disease in a murine model of CDI.
Abstract
Clostridioides difficile infection (CDI), characterized by colitis and diarrhea, afflicts approximately half a million people in the USA every year, burdening both individuals and the healthcare system. C. difficile 630Δerm is an erythromycin-sensitive variant of the clinical isolate C. difficile 630 and is commonly used in the C. difficile research community due to its genetic tractability. 630Δerm possesses a point mutation in perR, an autoregulated transcriptional repressor that regulates oxidative stress resistance genes. This point mutation results in a constitutively de-repressed PerR operon in 630Δerm. To address the impacts of perR on phenotypes relevant for oxygen tolerance and relevant to a murine model of CDI, we corrected the point mutant to restore PerR function in 630∆erm (herein, 630∆erm perRWT). We demonstrate that there is no difference in growth between 630Δerm and…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
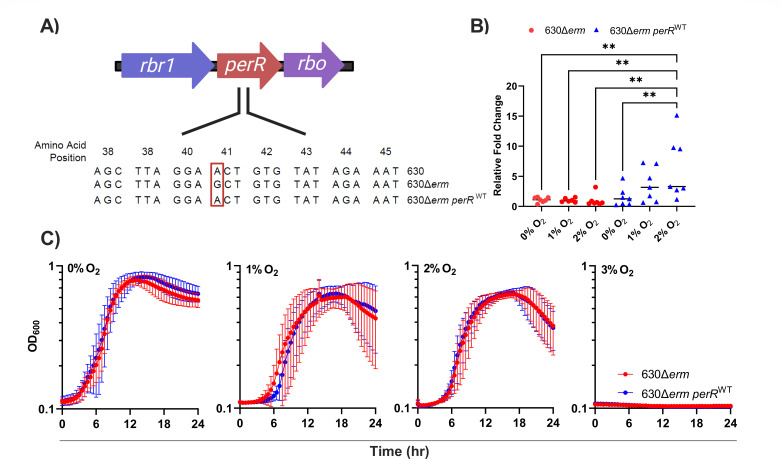 Fig 1
Fig 1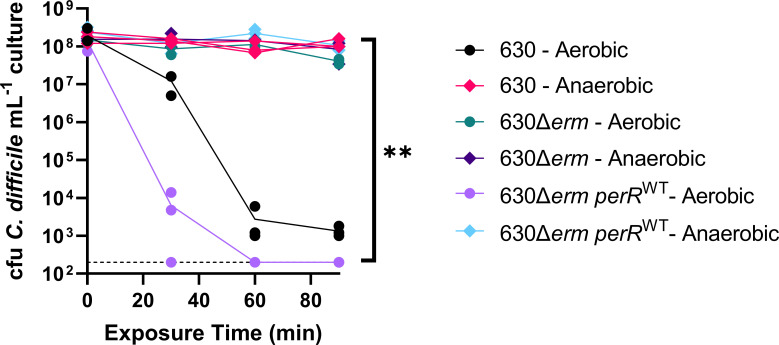 Fig 2
Fig 2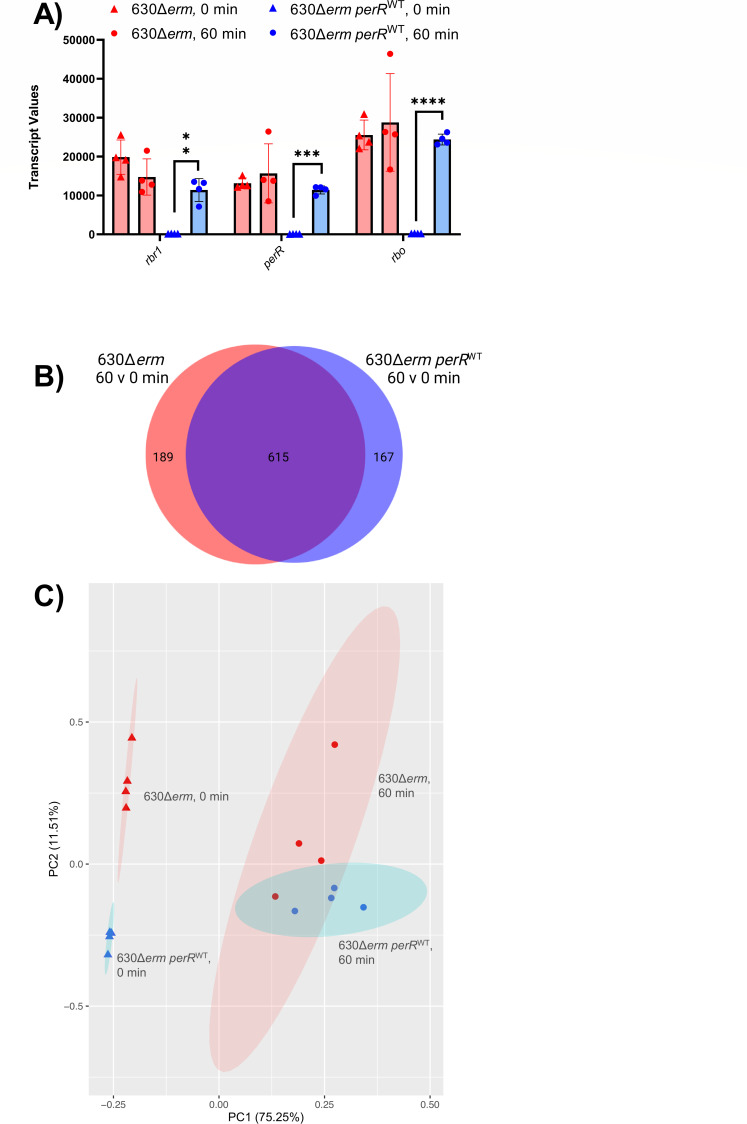 Fig 3
Fig 3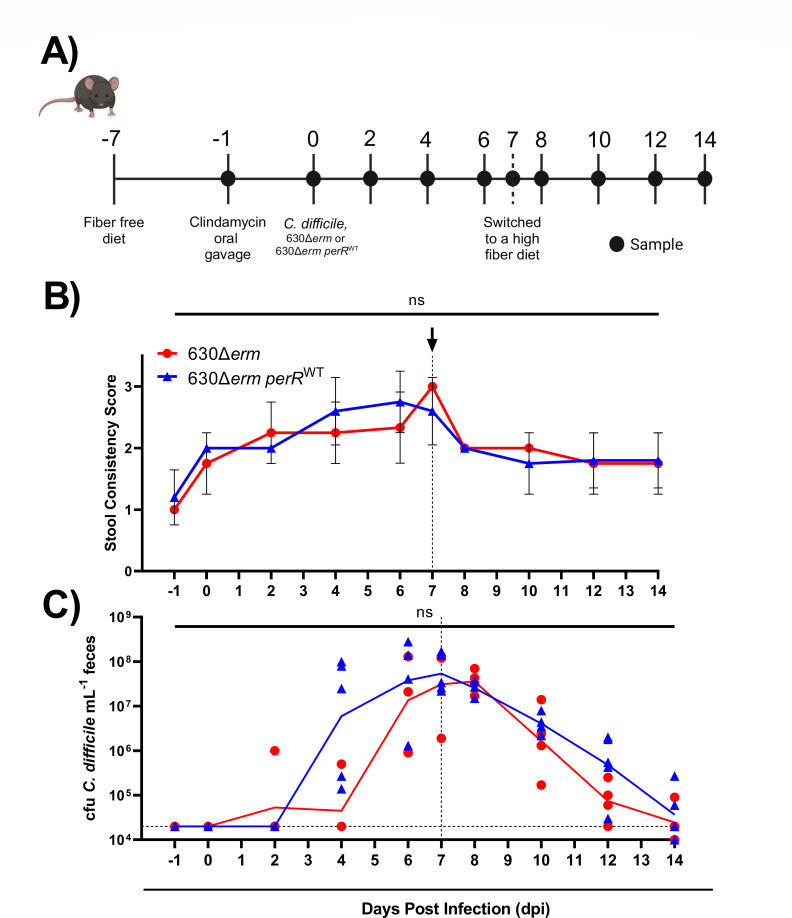 Fig 4
Fig 4| Locustag | Gene | Description | LogFC | |
|---|---|---|---|---|
| CDIF630erm_00875 | Putative diguanylate kinase signaling protein | 2.415049 | 0.003524 | |
| CDIF630erm_00944 |
| Putative rubrerythrin | 6.815261 | 7.17E-14 |
| CDIF630erm_00945 |
| Peroxide-responsive repressor | 6.884237 | 7.34E-14 |
| CDIF630erm_00946 | Desulfoferrodoxin | 6.840431 | 3.36E-14 | |
| CDIF630erm_00947 | Oxidative stress glutamate dehydrogenase (glutamate synthase-like) (rubredoxin) | 6.307636 | 1.02E-13 | |
| CDIF630erm_00948 | Putative metallo-beta-lactamase superfamily protein | 2.727654 | 6.00E-09 | |
| CDIF630erm_01262 | Putative conjugative transposon protein | 2.715317 | 0.000238 | |
| CDIF630erm_01792 | Putative diguanylate kinase signaling protein | 2.082062 | 0.00484 |
- —NSF | National Science Foundation Graduate Research Fellowship Program (GRFP)
- —University of Wisconsin-Madison (UW)
- —HHS | National Institutes of Health (NIH)
- —HHS | National Institutes of Health (NIH)
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsClostridium difficile and Clostridium perfringens research · Helicobacter pylori-related gastroenterology studies · Gut microbiota and health
INTRODUCTION
Clostridioides difficile is a leading cause of infectious diarrhea, resulting in an estimated 500,000 annual cases in the USA alone (1). A healthy microbiota typically prevents symptomatic C. difficile infection (CDI) through colonization resistance. However, ecological disturbances, commonly broad-spectrum antibiotics, disrupt the gut microbiota and give C. difficile access to vacant niches, which facilitates CDI (2–4). During dysbiosis, C. difficile can access nutrients otherwise consumed by the gut microbiota, overcome the effects of inhibitory metabolites produced by the microbiota/host, and increase in abundance in the gastrointestinal (GI) tract. During infection, C. difficile produces toxins (TcdA and TcdB; and CDT in hypervirulent strains), which induce host inflammation and are responsible for the diarrhea characteristic of CDI. This diarrhea contributes to transmission of C. difficile spores and allows C. difficile to gain a metabolic advantage by suppressing the recovery of the microbiota (3, 5–7). However, by inducing inflammation, C. difficile also causes the elevation of oxygen (O_2_) and reactive oxygen species (ROS) in the gut lumen (8–12). As an obligate anaerobe, C. difficile has evolved a variety of strategies to resist oxidative stress including sporulation, a versatile metabolism, and oxygen detoxification enzymes such as flavodiirons, rubrerythrins, and desulfoferrodoxin (Rbo) (9, 13–15). Previous work described various aspects of the response of C. difficile to oxidative stress (8, 16–18) and characterized proteins that play roles in oxidative stress tolerance (8–10, 16–19). One important regulator of a subset of these oxygen detoxification enzymes is the peroxide repressor (PerR).
PerR is an autoregulated transcriptional repressor and is a member of the ferric uptake repressor (Fur) family of proteins (19, 20). PerR is involved in oxidative stress responses in multiple bacterial species including Clostridium acetobutylcium, Bacillus subtilis, Streptococcus pyogenes, Streptococcus mutans, Staphylococcus aureus, Campylobacter jejuni, and Clostridioides difficile (21–27). Under anaerobic conditions, PerR binds to its target promoters and represses their expression. Oxidative stress triggers a metal-catalyzed histidine oxidation, and PerR undergoes a conformational change, causing it to release from its target promoters, which induces de-repression of the PerR regulon (20, 27). In S. pyogenes, a perR mutant was hyper-resistant to peroxide. However, it was highly attenuated in a murine model, demonstrating the importance of an appropriately regulated PerR regulon for virulence in vivo (22). A perR mutation identified in a S. mutans ΔspxA1 strain similarly rendered PerR inactive, priming the strain to tolerate oxidative stress. However, this work also showed that PerR had a limited impact on the transcriptional response of S. mutans to hydrogen peroxide (24). In C. acetobutylcium, an obligate anaerobe, a perR mutant was more aerotolerant than wild type, and it was determined that PerR regulates oxidative stress genes, including reverse rubrerythrins, flavodiiron proteins (FDPs), and superoxide-reducing Dfx, as well as two putative enzymes involved in central energy metabolism (28, 29).
In C. difficile, the operon containing perR consists of three genes: rubrerythrin (rbr1), perR, and a desulfoferrodoxin (rbo) (Fig. 1A). Genes within this operon are upregulated upon exposure to 1.5% O_2_ in vitro (8). In ex-germ-free mice mono-colonized with C. difficile and bi-colonized with C. difficile and Bacteroides thetaiotaomicron, the genes of the perR operon were among the 10% most abundant transcripts in C. difficile (2). Studies examining the activity of rbo demonstrated that when inactivated, C. difficile was more sensitive to oxygen exposure. Furthermore, when C. difficile rbo was expressed in Escherichia coli, it demonstrated superoxide scavenging activity (15). Taken together, these data indicate that perR expression is responsive to oxidative stress and that PerR and PerR-regulated genes are important for C. difficile to navigate oxidative stress both in vitro and perhaps in vivo.
*In vitro assays of 630Δerm and 630Δerm perRWT exposed to physiologically relevant levels of O2. (A) Genomic context of perR in C. difficile 630, 630Δerm, and 630Δerm perRWT. The operon containing perR consists of three genes: a rubrerythrin (rbr1), the transcriptional repressor PerR (perR), and a desulfoferrodoxin (rbo). 630Δerm has a point mutation in perR, resulting in a T41A amino acid substitution. (B) RT-qPCR of perR from 630Δerm (red) and 630Δerm perRWT (blue) exposed to 0%, 1%, or 2% O2. Data points represent the relative fold change compared within each strain grown under anaerobic conditions, with rpoC used as the housekeeping gene (n = 7). Statistical significance was determined by two-way analysis of variance (ANOVA) with Tukey’s post-hoc test; *P < 0.01. (C) Growth curves of 630Δerm (red) and 630Δerm perRWT (blue) in mRCM grown anaerobically or hypoxically in the presence of 1%, 2%, and 3% O2. Data points represent the mean OD600 (n = 6–8), and error bars represent the standard deviation. Panel A was created with Biorender.com under agreement #Z41E368.
C. difficile 630Δerm is an erythromycin-sensitive, lab-generated derivative of C. difficile 630 (herein, 630∆erm and 630, respectively). 630∆erm is amenable to allelic exchange procedures and is therefore commonly used in the field for generating knockout mutants. Seven spontaneous mutations were previously identified in 630Δerm relative to 630 (30). These mutations include a single nucleotide polymorphism (SNP) in perR, a SNP in eutG, a SNP in a transcriptional regulator of the GntR family (CD630_35630), and three SNPs in intergenic regions. Additionally, an 18 bp duplication is present in spo0A in 630Δerm (30). The perR point mutation in 630Δerm results in an amino acid substitution at position 41 (T41A) (Fig. 1A). This mutation affects the helix-turn-helix motif of the DNA-binding domain of PerR and results in a constitutively expressed perR operon regardless of O_2_ or H_2_O_2_ exposure. A constitutively expressed perR operon provides 630Δerm with a higher tolerance to O_2_ and H_2_O_2_ than parental strain 630, and plasmid-based complementation of perR from 630 into 630∆erm restores H_2_O_2_-dependent expression of one PerR-repressed gene, rbr1 (27).
Despite these previous findings on the role of perR in C. difficile and other organisms, there are several remaining questions relating to the direct effects of the perR point mutation in 630Δerm on oxidative stress resistance, gene expression, and the resulting impacts on infection. These are important gaps in knowledge, as this point mutation may impact interpretation of previous and future data because 630Δerm is such a widely used strain in the C. difficile field. To address these gaps, we corrected the perR point mutation in 630Δerm to create 630∆erm perR^WT^. We demonstrate that this strain has a repressible perR operon and that there is no growth difference between 630Δerm and 630∆erm perR^WT^ in the presence of 0%–3% O_2_. However, we show that 630Δerm is fitter than 630∆erm perR^WT^ when exposed to ambient air (21% O_2_). We also characterize 630Δerm and 630∆erm perR^WT^ transcriptomes exposed to ambient O_2_. Finally, using 630Δerm and 630∆erm perR^WT^, we demonstrate that functional PerR does not impact C. difficile burdens or diarrhea in a murine model of CDI.
RESULTS
Targeted restoration of the mutant perR allele in 630∆erm and impacts on growth and survival in the presence of oxygen
To determine the impact of a constitutively expressed PerR on in vitro phenotypes, we corrected the point mutation in 630Δerm, generating 630Δerm perR^WT^ (Fig. 1A) (27). Correction of the point mutation was confirmed by whole-genome sequencing and alignment, comparing the parental 630Δerm to 630∆erm perR^WT^. A single SNP at position 1,006,274 (G → A) was identified, indicating the only genetic difference between the two strains was the restoration of a wild-type perR.
To confirm wild-type PerR function in 630Δerm perR^WT^, perR-specific RT-qPCR was performed on RNA extracted from 630Δerm and 630Δerm perR^WT^ grown in the presence of 0%, 1%, and 2% O_2_ (Fig. 1B). These O_2_ concentrations were selected because they reflect those present in the colon (31, 32). This analysis revealed that perR transcription was non-responsive to oxygen in 630Δerm, due to constitutive de-repression of its operon. However, perR transcripts in 630Δerm perR^WT^ were elevated as a function of increasing oxygen exposure, which demonstrates the perR operon is de-repressed upon exposure to 2% O_2_. These data confirm that correcting the perR point mutation restored a wild-type, oxygen-responsive phenotype and are supported by previously published RNA-seq data that showed that perR is upregulated in 630 at 1.5% O_2_ (8).
To evaluate the impacts of oxygen-responsive perR expression on C. difficile growth, 630Δerm and 630Δerm perR^WT^ were grown in a complex, rich medium (modified Reinforced Clostridial Medium [mRCM]) in the presence of 0%, 1%, 2%, and 3% O_2_ (Fig. 1C; Fig. S1) (3, 33). We did not observe differences in growth kinetics between the two strains at these O_2_ concentrations. However, the growth of both strains was negatively impacted by increasing O_2_ concentration, and no growth was observed in 3% O_2_. These data partially replicate results of a recent study on O_2_ reductases in C. difficile. There, targeted restoration of PerR function in 630∆erm did not impact C. difficile growth at 1% O_2_ (34).
Despite differences in PerR activity between 630∆erm and 630∆erm perR^WT^, a constitutively de-repressed PerR regulon offers no apparent fitness advantage at physiologically relevant O_2_ concentrations (Fig. 1). Previous work showed that 630∆erm has increased survival relative to 630 upon exposure to ambient O_2_ (27). To determine if 630∆erm perR^WT^ has restored sensitivity to an ambient air (~21% O_2_), an ambient air exposure assay was performed. Cell viability of 630, 630Δerm, and 630Δerm perR^WT^ was quantified after exposure to ambient air for 0 to 90 minutes. Viability of 630 and 630Δerm perR^WT^ were decreased upon exposure to ambient air. However, this exposure did not have an impact on 630∆erm, demonstrating that targeted restoration of perR restores wild-type levels of sensitivity to ambient air in C. difficile (Fig. 2).
*Aerotolerance assay of 630, 630Δerm, and 630Δerm perRWT exposed to ambient air. Aliquots of stationary phase cultures of each strain were exposed to ambient air for 0, 30, 60, and 90 minutes, or maintained under anaerobic conditions and plated on pre-reduced CDMN agar at the indicated timepoints. Colonies were quantified after overnight growth in an anaerobic chamber (n = 3 cultures per strain per condition). Statistical testing was determined by two-way analysis of variance (ANOVA); *P < 0.01.
Differences in 630Δerm and 630Δerm perRWT transcriptomes as a function of ambient air exposure
To better understand genes involved in 630∆erm resistance to ambient air (Fig. 2), we performed RNA-seq on 630Δerm and 630Δerm perR^WT^ at 0 and 60 minutes post-ambient air exposure and compared transcriptomes between the two strains and two timepoints (Tables S2 to S5). The 60 minute timepoint was chosen based on RT-qPCR of perR transcripts from total RNA extracted from 630 at 0, 15, 30, and 60 minutes post-air exposure (Fig. S2), which showed elevated perR transcripts at both 30 and 60 minutes of aerobic exposure compared to anaerobic control. RNA extracted at 60 minutes was high quality via Bioanalyzer (data not shown) and therefore selected as the timepoint. RNA-seq showed that the operon containing perR was responsive to aerobic exposure in 630Δerm perR^WT^ but was expressed at high levels regardless of aerobic exposure in 630∆erm (Fig. 3A). In addition to the genes in this operon, transcripts of five other genes were more abundant under anaerobic conditions in 630∆erm relative to 630∆erm perR^WT^ (Table 1), suggesting a small subset of genes are repressed by PerR in C. difficile. Similarly, there were largely overlapping patterns of gene expression for both strains between 0 and 60 minutes of ambient air exposure. Specifically, although there were differences in cell viability at the 60 minute timepoint (Fig. 2), both strains shared 615 differentially regulated genes (Fig. 3B), many of which were previously predicted/characterized to be involved in oxidative stress resistance (Table S6). This similarity in transcriptional response to ambient air is also evident from principal component analysis (Fig. 3C), which shows that both strain and exposure to O_2_ had a significant impact on the transcriptome (the impact of O_2_ exposure was more significant), but the interaction of the variables was not significant (permutation multivariate analysis of variance [PERMANOVA] ~ time * genotype, time P = 0.001; strain P = 0.006, time * strain, P = 0.085).
*Transcriptional profiling of 630Δerm and 630Δerm perRWT exposed to ambient air. 630Δerm and 630Δerm perRWT were grown to mid-log phase in mRCM and then exposed to ambient air for 0 and 60 minutes. RNA-seq was performed on n = 4 independent cultures, for each strain at each timepoint. (A) Transcript values of the genes in the perR operon (n = 4). Statistical testing was determined by paired t-test; *P < 0.05, **P < 0.01, ***P < 0.001, and ***P < 0.0001. (B) A Venn diagram illustrating that the majority of significantly differentially regulated genes (FC ≥ |2|, P < 0.05) overlapped between the 630Δerm and 630Δerm perRWT. (C) Principal component analysis (PCA) plot of RNA-seq data from 630Δerm (red) and 630Δerm perRWT (blue) exposed to ambient air for 0 (triangle) and 60 (circle) minutes. Ellipses represent 95% confidence intervals based on strain and time of ambient air exposure. Panel B was created with Biorender.com under agreement #N27K433.
These data suggest that the increased viability of 630∆erm relative to 630Δerm perR^WT^ could be due to elevated levels of PerR-dependent gene products present in 630∆erm cells prior to oxygen exposure, which would prime the cells for oxidative stress. Beyond rbr1, perR, and rbo, five genes were upregulated in 630∆erm relative to 630Δerm perR^WT^ at the t = 0 timepoint Table 1. These genes include a putative oxidative stress glutamate dehydrogenase (CDIF630erm_00947), a putative metallo-beta-lactamase superfamily protein (CDIF630erm_00948), a putative conjugative transposon protein (CDIF630erm_01262), and two putative diguanylate kinase signaling proteins (CDIF630erm_00875 and CDIF630erm_01792). These data suggest that although PerR impacts C. difficile survival in ambient air, PerR does not exert major control, outside of the perR operon, over oxidative stress resistance genes in C. difficile, as previously observed in other microbes (21, 22, 24, 25, 28, 29).
630Δerm and 630Δerm perRWT do not differ in their ability to infect or cause diarrhea in mice
During infection, C. difficile toxins induce host inflammation and elevate ROS (35–37). Therefore, given that 630Δerm and 630Δerm perR^WT^ differ in their ability to survive oxidative stress, we sought to determine the impact of perR on strain fitness during infection. To examine this, we leveraged a well-established murine model of CDI (33, 38). Mice were placed on a fiber-free (FF) diet and gavaged with clindamycin as in Figure 4A to reduce colonization resistance against C. difficile. Then, mice were gavaged with either 630Δerm or 630Δerm perR^WT^ to establish CDI. After 1 week of infection, the mice were switched to a high-fiber diet to determine if fiber-dependent CDI clearance kinetics differ between the strains (38).
CDI in mice infected with 630Δerm and 630Δerm perRWT. (A) Conventional, age-matched male C57BL/6 mice were placed on a fiber-free diet, gavaged with clindamycin, and infected with either 630Δerm or 630Δerm perRWT (n = 4–5). Mice were switched to a high-fiber diet 7 days after being infected with C. difficile. (B) Average stool consistency scores: 1 = hard, dry pellet; 2 = soft but fully formed pellet; 3 = runny, poorly formed pellet. (C) The geometric means of C. difficile burdens for each strain over the experimental time course. The limit of detection for this assay is indicated in the horizontal dashed line at 20,000 CFU C. difficile/mL feces. Statistical testing was performed on CFU counts and stool consistency from mice between strains on each day and was not significant (ns) via Mann-Whitney test. Panel A was created with Biorender.com under agreement #Z41E368. The vertical dashed lines at 7 days post-infection in panels B and C indicate when the mice were switched from a fiber-free to a high-fiber diet.
Consistent with previous literature using 630 in murine models (39, 40), both 630∆erm and 630∆erm perR^WT^ induced mild CDI symptoms. Specifically, there was no difference between the two strains in stool consistency scores nor C. difficile burdens across the 14 days (Fig. 4B and C). All mice experienced an increase in stool softness on 0 day post-infection (dpi) due to administration of clindamycin (Fig. 4B). However, after infection with C. difficile, mice experienced an increase in diarrhea, regardless of the strain with which they were infected. For both strains, fecal burdens of C. difficile were similar when on the fiber-free diet and decreased once the mice were placed on the high-fiber diet, as previously demonstrated (Fig. 4C) (38). Interestingly, there was a slight delay in C. difficile burdens in both strains at early timepoints. Specifically, consistent burdens were not seen until 4 dpi, possibly due to sensitivity of the strains to clindamycin (Fig. S3). Taken together, these data demonstrate that 630Δerm and 630Δerm perR^WT^ do not differ in their ability to infect or cause diarrhea in mice.
DISCUSSION
630Δerm has been an important strain for elucidating the physiology and pathogenesis of C. difficile because of its extensive use to generate mutants via allelic exchange procedures. However, there are several mutations in 630∆erm relative to its parent strain (a clinical isolate) that may impact the generalizability of the findings made in 630∆erm to other C. difficile strains. The mutations in 630∆erm include a point mutation in perR that renders its PerR regulon constitutively de-repressed. Despite knowledge of this perR point mutation and its impacts on expression of genes within the perR operon, the effects of this mutation on C. difficile oxidative stress resistance, on the broader C. difficile transcriptome, and on CDI phenotypes remained poorly characterized.
Here, we restored a wild-type copy of perR in 630∆erm to create 630Δerm perR^WT^. We determined that there is no difference in growth between 630Δerm and 630Δerm perR^WT^ at physiologically relevant O_2_ levels (Fig. 1). However, using these strains, we showed that a constitutively de-repressed PerR regulon allows C. difficile 630Δerm to tolerate ambient air exposure (Fig. 2). Previous work that compared the survival of C. difficile exposed to ambient O_2_ compared 630 and 630∆erm, which have multiple genetic differences in addition to the perR point mutation (27). Our results support the previous findings and the hypothesis that the differences in oxidative stress resistance in these strains were due to perR and not to these other mutations. In addition, given that the T41A point mutation in perR is unique to 630Δerm when compared to 11 diverse clinical isolates of C. difficile (including its parent strain 630) (27), it is reasonable to assume that this mutation was selected due to oxygen exposure during laboratory passage.
To better understand the genes repressed by PerR (which likely contribute to increased O_2_ tolerance by 630∆erm), we performed RNA-seq on 630∆erm and 630∆erm perR^WT^ exposed to ambient air (Fig. 3). This analysis suggested that PerR represses a small fraction of genes in C. difficile. Under anaerobic conditions, eight genes were upregulated in 630∆erm relative to 630∆erm perR^WT^ (Table 1; Table S2). This includes the genes present in the perR operon (rbr1, perR, and rbo), a putative oxidative stress glutamate dehydrogenase, a putative metallo-beta-lactamase, a putative conjugative transposon protein, and two putative diguanylate kinase signaling proteins. Our RNA-seq data also show that large-scale changes to the C. difficile transcriptome occur at 60 minutes post-air exposure, regardless of whether the strain has functional PerR. Specifically, 615 shared genes are differentially regulated in both strains at 60 minutes post-exposure to ambient air relative to the pre-exposure timepoint. This include genes involved in oxidative stress resistance (Table S6), many of which are likely under the control of other oxidative stress regulators (e.g., σ^B^) (12–14, 34, 41). Despite similarities of the transcriptional responses of 630∆erm and 630∆erm perR^WT^ to ambient air, 630∆erm and 630∆erm perR^WT^ have some differences in their responses to this treatment (Fig. 3). Deeper analysis of functional categories of these genes (Tables S6 to S8) revealed oxygen-dependent upregulation of genes involved in Stickland fermentation, ribosome synthesis, and the CRISPR system in 630∆erm and oxygen-dependent downregulation of peptidoglycan and teichoic acid metabolism in 630∆erm perR^WT^. These changes in gene expression mirror the differential survival of these two strains in the presence of oxygen (42).
Our data also indicate that there is no difference between 630Δerm and 630Δerm perR^WT^ in C. difficile burdens nor severity of infection in a murine model of CDI (Fig. 4). While our work was performed in conventional mice, previous work showed that rbo, perR, and rbr were among the top 10% most highly expressed genes in gnotobiotic mice infected with 630 (2), suggesting that PerR-dependent gene expression is important during infection. Because longitudinal and radial oxygen gradients are present in the gastrointestinal tract and O_2_ levels can be elevated by antibiotic treatment, it is possible that C. difficile encounters enough O_2_ to de-repress PerR-dependent genes during the onset, establishment, or maintenance of murine infection (31, 32, 41, 43). Therefore, the mouse experiments performed may disguise possible differences in fitness (positive or negative) due to a constitutively de-repressed PerR operon.
In summary, our work establishes that a constitutively de-repressed PerR regulon offers no fitness advantage at O_2_ levels encountered in the distal GI tract (Fig. 1) nor in a mouse model of CDI (Fig. 4). However, a constitutively de-repressed PerR regulon provides C. difficile with tolerance to ambient air (Fig. 2) and impacts gene expression in C. difficile in the presence and absence of oxygen (Fig. 3). Unique to 630Δerm, the perR mutation invokes consideration of selective pressures that this strain may have encountered during exposure to ambient air in laboratory settings (27). This study adds to a growing body of literature on the ways in which obligate anaerobes resist oxidative stress and will contribute to future work in understanding these responses in C. difficile.
MATERIALS AND METHODS
Bacterial strains and culture conditions
C. difficile strains 630, 630Δerm, and 630Δerm perR^WT^ (44, 45) were maintained as −80°C stocks in 25% glycerol under anaerobic conditions in septum-topped vials. C. difficile strains were struck out on Clostridioides difficile agar with moxalactam and norfloxacin (CDMN agar), which is C. difficile agar base (Oxoid) supplemented with 32 mg/L moxalactam (Santa Cruz), 12 mg/L norfloxacin (Sigma-Aldrich), and 7% defibrinated horse blood (HemoStat Laboratories), and were cultured anaerobically for 24 hours. A single colony was picked into 5 mL of pre-reduced BD Difco reinforced clostridial medium (RCM) or a modified RCM (mRCM; RCM without soluble starch and agar) (33). Liquid cultures were grown at 37°C anaerobically for 16–24 hours and used as inocula for growth curves, aerotolerance assays, RNA-seq, RT-qPCR, and murine experiments. All bacterial growth media were pre-reduced for at least 24 hours in an anaerobic chamber (Coy) prior to use in experiments.
For in vitro growth curves, subcultures were prepared at 1:100 dilution in mRCM (3). Growth curves were performed anaerobically or in a hypoxic chamber at 1%, 2%, and 3% O_2_ (Coy). Clindamycin sensitivity growth curves were performed anaerobically in mRCM with clindamycin concentrations spiked into each well. All growth curves were performed in sterile polystyrene 96-well tissue culture plates (Falcon) with low-evaporation lids using a BioTek Epoch2 plate reader at 30 minute intervals. Plates were shaken on the orbital setting for 10 seconds before each read. The OD_600_ of the cultures was recorded using Gen5 software (version 3.10.06).
Generation of C. difficile 630Δerm perRWT
The perR point mutation was corrected using the PyrE allelic exchange system (46). Primers indicated in Table S1 were used to amplify perR from 630 genomic DNA. The amplicon containing perR from 630 was ligated into pMTL-YN3 after AscI and SbfI digestion of the vector and was inserted using New England Biolabs Quick Ligation Kit (M2200S). The plasmid construct was transformed and propagated into One Shot Top10 E. coli (Invitrogen) before transformation into conjugation-proficient E. coli HB101/pRK24 cells. The pMTL-YN3 with 630 perR was conjugated into 630∆erm∆pyrE. Plasmid integrants were selected using brain-heart infusion supplemented (BHIS) agar containing thiamphenicol (10 or 15 µg/mL), cefoxitin (8 µg/mL), kanamycin (30 or 50 µg/mL), and uracil (5 µg/mL). Double crossover events were selected using a defined minimal medium for C. difficile (CDDM) supplemented with uracil (5 µg/mL) and 5-fluoroorotic acid (2 mg/mL). Whole colony PCR using GoTaq Green Master Mix (Promega) amplified perR from potential clones. HypC4III digestion of PCR-purified perR (Zymo DNA Clean and Concentrator-5) confirmed 630 perR integration as this restriction enzyme digests 630 perR but is unable to recognize that cut site in 630Δerm due to the point mutation (Fig. S4). After confirmation of 630 perR integration, the pyrE locus was restored using pMTL-YN1C to generate 630Δerm perR^WT^.
Illumina whole-genome sequencing on DNA extracted (47) from 630Δerm and 630Δerm perR^WT^ was performed by Microbial Genome Sequencing Center (MiGS) with a minimum read count of 1.33 million reads (200 Mbp) per sample. Raw sequencing data were assembled using a reference-guided assembly pipeline (https://github.com/pepperell-lab/RGAPepPipe_CHTC) to C. difficile strain 630Δerm (GenBank: LN614756.1) as previously described (48). Briefly, Fastqc v0.12.1 (49) assessed the quality of the sequences, which were then trimmed using Trimmomatic v0.39 (50). Sequences were aligned to the reference using BWA Mem v0.7.18 (51), and alignments were processed using SAMtools v1.21 (52). Picard v 2.18.25 (https://github.com/broadinstitute/picard) was used to remove duplicates and add read groups. Pilon v 1.78 (53) identified variants. Assembly and alignment quality was determined using Qualimap BamQC v2.2.1 (54). The mean coverage was 160× for 630Δerm and 50× for 630Δerm perR^WT^. Both samples had >95% aligned reads to the reference. To identify the SNP difference between 630Δerm and 630Δerm perR^WT^, a variant call format (VCF) was created using SnpSites v2.5.1 (55).
Aerotolerance assays
Liquid cultures of 630, 630Δerm, and 630Δerm perR^WT^ were grown anaerobically in RCM overnight. Each culture (200 µL) was aliquoted into sterile polystyrene 96-well tissue culture plates, with four replicate plates set up, and was exposed to ambient air for 0, 30, 60, and 90 minutes at room temperature. Aeration of cultures using a multichannel pipette was performed immediately after removing from the chamber and approximately every 10 minutes throughout the assay. At each timepoint, one of the 96-well plates was passaged back into the anaerobic chamber. Serial dilutions were performed using pre-reduced phosphate-buffered saline (PBS) and plated on pre-reduced CDMN. Plates were incubated anaerobically for 24 hours at 37°C, and colonies present on CDMN plates were quantified.
Transcriptional profiling of C. difficile response to ambient air
C. difficile 630Δerm and 630Δerm perR^WT^ overnight cultures were back-diluted 1:100 into 35 mL of pre-reduced mRCM in Erlenmeyer flasks and were incubated anaerobically at 37°C until cultures reached a mid-log phase (OD_600_ = 0.3–0.4). At mid-log phase, 5 mL aliquots of the cultures were diluted 1:1 in chilled 1:1 ethanol:acetone and were stored at −20°C to preserve RNA. The remaining cultures were aerobically shaken (220 rpm) at 37°C for 60 minutes. After 60 minutes, 5 mL aliquots were diluted 1:1 in chilled 1:1 ethanol:acetone and were stored at −20°C to preserve RNA (56).
RNA was extracted by centrifuging samples at 3,000 × g for 5 minutes at 4°C. Pellets were washed with 5 mL cold, nuclease-free PBS and were centrifuged at 3,000 × g for 5 minutes at 4°C. The supernatant was removed, and remaining pellets were resuspended in 1 mL TRIzol and processed using a TRIzol Plus RNA Purification Kit (Thermo) with on-column DNase treatment. Purified RNA integrity was confirmed via 2100 Agilent BioAnalyzer and was frozen at −80°C.
RNA-seq was performed by MiGS on high-quality rRNA-depleted RNA extracts (12 million paired-end reads per sample). Quality control and adapter trimming were performed with bcl2fastq (version 2.20.0.445) (57). Read mapping was performed with HISAT2 (version 2.2.0) (58). Read quantification was performed using Subread’s featureCounts (version 2.0.1) (59) functionality. Read counts were loaded into R (version 4.0.2) (60) and were normalized using edgeR’s (61) Trimmed Mean of M values (TMM) algorithm (version 1.14.5). Subsequent values were then converted to counts per million (cpm). Differential expression analysis was performed using edgeR’s exact test for differences between two groups of negative-binomial counts with an estimated dispersion value of 0.1. Transcript level quantification, count normalization, and differential expression analysis were provided using C. difficile 630Δerm (GCA_002080065.1_ASM208006v1) as the reference genome.
The principal component analysis (PCA) plot was generated from the RNA-seq data in R using version 4.4.1. PCA was performed using prcomp from stats package (version 4.4.1) and was visualized using ggplot2 (version 3.5.1). Confidence ellipses were generated using stat_ellipse as implemented in ggplot2. PERMANOVA was assessed by vegan::adonis2 (version 2.6-6.1).
RT-qPCR of perR in C. difficile exposed to varying levels of O2
To identify a timepoint for RNA-seq at which perR was de-repressed, C. difficile 630 overnight cultures were back-diluted 1:100 into 35 mL of pre-reduced RCM and were incubated at 37°C until it reached a mid-log phase (OD_600_ = 0.3–0.4). At mid-log phase, one set of cultures was incubated aerobically at 37°C, shaking at 220 rpm, while the other culture was incubated at 37°C anaerobically. After 0, 15, 30, and 60 minutes, 5 mL aliquots of the cultures were diluted 1:1 in chilled 1:1 ethanol:acetone and were stored at −20°C to preserve RNA. RNA was extracted as described above.
RT-qPCR was performed using GoTaq 1-Step RT-qPCR Master Mix (Promega) according to the manufacturer’s instructions, with a concentration of 1 ng/µL RNA in a final volume of 10 µL. Each reaction was run with three technical replicates. The RT-qPCRs were performed on a QuantStudio 7 Flex (Applied Biosystems), and the threshold cycle (Ct) was determined using QuantStudio Real-Time PCR Software v1.7.2. The cycle run involved a reverse transcriptase activation and inactivation step of 40°C for 15 minutes and 95°C for 10 minutes. The PCR cycle was 95°C for 10 seconds, 60°C for 30 seconds, and 72°C for 30 seconds for 40 cycles, with a melt curve performed afterward. Relative fold change was determined by comparing the Ct values to the average Ct value at 0 minutes for each condition.
To quantify perR transcript levels of C. difficile 630Δerm and C. difficile 630Δerm perR^WT^ exposed to low levels of O_2_, overnight cultures of each strain were back diluted 1:50 into 14 mL of pre-reduced mRCM in 100 × 15 mm circular petri dishes (VWR) to optimize exposure to oxygen. Cultures were incubated at 37°C in 0%, 1%, and 2% O_2_ until they reached a mid-log phase (OD_600_ = 0.3–0.4). At mid-log phase, 5 mL aliquots of the cultures were diluted 1:1 in chilled 1:1 ethanol:acetone and were stored at −20°C to preserve RNA, and RNA was extracted as described above. RT-qPCR was performed using GoTaq 1-Step RT-qPCR Master Mix (Promega), with a concentration of 2 ng/µL of RNA in a final volume of 20 µL. Each reaction was set up with primers amplifying perR or rpoC (housekeeping gene) (Table S1), with two to three technical replicates. The RT-qPCRs were performed on a QuantStudio 7 Flex (Applied Biosystems) as previously described. Relative fold change was determined using the 2^-ΔΔCT^ method by comparing the Ct values to the anaerobic Ct value within each strain (62).
Murine model of C. difficile infection (CDI)
All animal studies were carried out in strict accordance with the University of Wisconsin-Madison Institutional Animal Care and Use Committee (IACUC) guidelines (Protocol #M006305). CDI murine model was performed on age- and sex-matched, conventionally reared C57BL/6 mice bred in-house. Mice used in experiments were between 6 and 9 weeks of age. Mice were fed a fiber-free (FF) diet (Inotiv TD.150689) 1 week before antibiotic exposure. Mice were given a single dose of clindamycin by oral gavage (1 mg/mouse; 200 µL of 5 mg/mL solution); 24 hours later, they were given 200 µL of 630Δerm or 630Δerm perR^WT^ overnight cultures grown in RCM, via oral gavage (n = 4–5 mice per condition; average inoculum 8.6 × 10^7^ CFU/mL).
At 7 dpi, mice were switched from the FF diet to a fiber-rich standard rodent chow (Inotiv Teklad 2916) to observe fiber-dependent C. difficile clearance kinetics (33, 38). Throughout the entire experiment, feces were collected from mice directly into a microcentrifuge tube and were kept on ice. To quantify C. difficile burdens, 1 µL of each fecal sample was collected with a disposable inoculating loop and was resuspended in 200 µL PBS. Tenfold serial dilutions of fecal suspension were prepared in sterile polystyrene 96-well tissue culture plates (Falcon). For each sample, 10 µL aliquots of each dilution, with two technical replicates, were spread on CDMN agar. CDMN plates were incubated anaerobically at 37°C for 16–24 hours. Colonies were quantified and technical replicates were averaged to determine C. difficile burdens (limit of detection = 2 × 10^4^ CFU/mL). Stool consistency scores were also noted while processing fecal pellets. Fecal pellets were assigned a score of 1 = hard, dry pellets, difficult to transect with a disposable plastic culture loop; 2 = soft, fully formed pellets, easy to transect with a culture loop; or 3 = runny, poorly formed pellets, no pressure required to transect with a culture loop (3).
Statistical analysis
All statistical analyses, except for the PERMANOVA, were performed using GraphPad Prism 9.4.1. PERMANOVA was performed as described in “PCA Generation and RNA-seq Data Analysis” section. Details of specific statistical analyses are indicated in figure legends. For all figures, *P < 0.05, **P < 0.01, ***P < 0.001, and ****P < 0.0001.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Guh AY, Mu Y, Winston LG, Johnston H, Olson D, Farley MM, Wilson LE, Holzbauer SM, Phipps EC, Dumyati GK, Beldavs ZG, Kainer MA, Karlsson M, Gerding DN, Mc Donald LC, Emerging Infections Program Clostridioides difficile Infection Working Group. 2020. Trends in U.S. burden of Clostridioides difficile infection and outcomes. N Engl J Med 382:1320–1330. doi:10.1056/NEJ Moa 191021532242357 PMC 7861882 · doi ↗ · pubmed ↗
- 2Ferreyra JA, Wu KJ, Hryckowian AJ, Bouley DM, Weimer BC, Sonnenburg JL. 2014. Gut microbiota-produced succinate promotes C. difficile infection after antibiotic treatment or motility disturbance. Cell Host Microbe 16:770–777. doi:10.1016/j.chom.2014.11.00325498344 PMC 4859344 · doi ↗ · pubmed ↗
- 3Battaglioli EJ, Hale VL, Chen J, Jeraldo P, Ruiz-Mojica C, Schmidt BA, Rekdal VM, Till LM, Huq L, Smits SA, Moor WJ, Jones-Hall Y, Smyrk T, Khanna S, Pardi DS, Grover M, Patel R, Chia N, Nelson H, Sonnenburg JL, Farrugia G, Kashyap PC. 2018. Clostridioides difficile uses amino acids associated with gut microbial dysbiosis in a subset of patients with diarrhea. Sci Transl Med 10:eaam 7019. doi:10.1126/scitranslmed.aam 701930355801 PMC 6537101 · doi ↗ · pubmed ↗
- 4Jenior ML, Leslie JL, Young VB, Schloss PD. 2017. Clostridium difficile colonizes alternative nutrient niches during infection across distinct murine gut microbiomes. m Systems 2:e 00063-17. doi:10.1128/m Systems.00063-1728761936 PMC 5527303 · doi ↗ · pubmed ↗
- 5Koenigsknecht MJ, Theriot CM, Bergin IL, Schumacher CA, Schloss PD, Young VB. 2015. Dynamics and establishment of Clostridium difficile infection in the murine gastrointestinal tract. Infect Immun 83:934–941. doi:10.1128/IAI.02768-1425534943 PMC 4333439 · doi ↗ · pubmed ↗
- 6Jenior ML, Leslie JL, Young VB, Schloss PD. 2018. Clostridium difficile alters the structure and metabolism of distinct cecal microbiomes during initial infection to promote sustained colonization. m Sphere 3:e 00261-18. doi:10.1128/m Sphere.00261-1829950381 PMC 6021602 · doi ↗ · pubmed ↗
- 7Litvak Y, Byndloss MX, Bäumler AJ. 2018. Colonocyte metabolism shapes the gut microbiota. Science 362:eaat 9076. doi:10.1126/science.aat 907630498100 PMC 6296223 · doi ↗ · pubmed ↗
- 8Weiss A, Lopez CA, Beavers WN, Rodriguez J, Skaar EP. 2021. Clostridioides difficile strain-dependent and strain-independent adaptations to a microaerobic environment. Microb Genom 7:000738. doi:10.1099/mgen.0.00073834908523 PMC 8767335 · doi ↗ · pubmed ↗