A maize architect: An epiallele of a PfkB-type carbohydrate kinase affects plant growth and development
Meenu Singla-Rastogi

Abstract
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
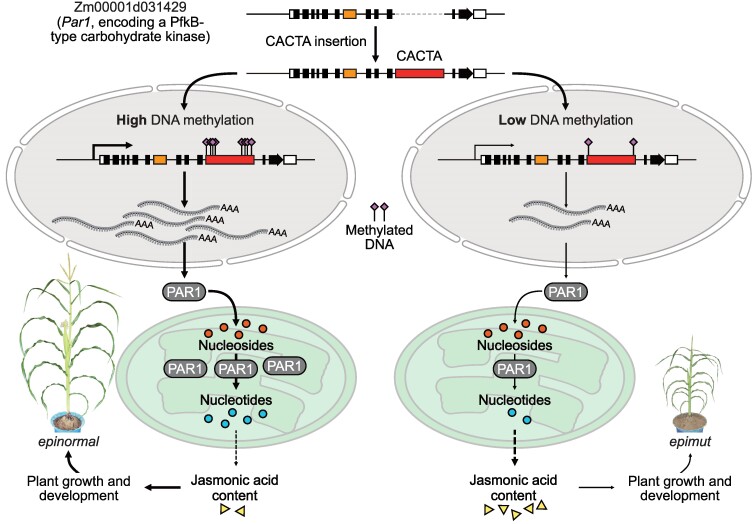 Figure 1
Figure 1Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsPlant nutrient uptake and metabolism · Biofuel production and bioconversion
The maize (Zea mays) genome is known for its complexity, largely due to the high content of transposable elements (TEs), which contribute significantly to both its size and structural variation (Schnable et al. 2009). Transposons, also known as “jumping genes,” are DNA sequences capable of moving from one location to another within the genome. In maize, transposons account for nearly 85% of the genome—substantially more than in many other organisms. The activity of these transposons is tightly regulated, often through epigenetic mechanisms that suppress their movement. These mechanisms include DNA methylation and histone modifications, which maintain genome stability by silencing transposon activity (Slotkin and Martienssen 2007). However, when these regulatory processes are disrupted, transposons become active again, potentially leading to undesired mutations or genomic instability. Interestingly, controlled activation of transposons has allowed researchers to explore genetic diversity and discover new traits, contributing to the development of maize varieties with higher yields, enhanced stress tolerance, and improved quality.
Plant architecture plays a significant role in determining the grain yield in maize. The way a maize plant is structured—how it develops leaves, branches, ears, and roots—directly influences its ability to capture light, absorb nutrients, and support grain production. In a new work, Ruonan Li and colleagues (Li et al. 2025) discovered and characterized a naturally occurring epiallele (epigenetic variant) of a gene encoding a phosphofructokinase B (PfkB)-type carbohydrate kinase, which they named Plant architecture 1 or par1. While constructing a recombinant inbred line population using maize inbred Zheng58 and the landrace BDB, the authors identified a spontaneous mutant with pleiotropic phenotype, exhibiting reduced plant height, shorter and narrower leaves, and larger leaf angle than the wild-type plants. Further genetic analysis revealed that the par1 epiallele was caused by a de novo intronic insertion of a CACTA transposon, a member of Class II transposons known for their “cut-and-paste” mechanism of transposition (Wicker et al. 2007). To confirm the role of the CACTA transposon in the par1 phenotype, the authors conducted genotyping of the F_2_ population. Surprisingly, even though all the mutant plants were homozygous for the insertion, only about one-half of the plants with homozygous insertion exhibited the mutant phenotype. As a result, the authors categorized the plants with homozygous insertion into two groups: epinormal (with a wild-type phenotype) and epimutant (with the mutant phenotype).
This phenomenon of differing phenotypes despite identical genetic sequences is akin to what is observed in identical human twins, where subtle phenotypic differences arise due to epigenetic divergence (Wong et al. 2005). It has been shown that CACTA transposons contribute to phenotypic diversity in maize, with their effects modulated by DNA methylation patterns (Wittmeyer et al. 2018). Through bulk segregant RNA-seq and RT-PCR analyses, the researchers confirmed that the par1 gene was normally expressed in epinormal plants, but its expression was significantly reduced in epimutants. These differences in expression were correlated with the phenotypic differences between the two groups. To identify the underlying epigenetic factors, the researchers employed whole-genome bisulfite sequencing and examined DNA methylation levels around the CACTA insertion site. The analysis revealed high DNA methylation at the edges of the transposon in epinormal plants and low methylation levels in epimutants. This finding supports the idea that DNA methylation influences gene expression, as increased CHH (where H corresponds to A, T, or C) methylation upstream of a gene is typically associated with higher gene expression levels (Li et al. 2015).
Further biochemical analysis revealed that the par1 epiallele encodes PAR1, a PfkB-type carbohydrate kinase capable of phosphorylating several nucleosides (inosine, uridine, adenosine, cytidine, and guanosine) into nucleoside monophosphates. This process is part of a nucleotide salvage pathway, which converts nucleosides or nucleobases into their phosphorylated forms. While the role of nucleoside kinases in Arabidopsis has been well studied (Witte and Herde 2020), there are fewer reports on their function in other plants. This study provides valuable insights into the role of PfkB-like proteins in regulating the abundance of nucleosides and nucleotides, which are crucial for plant development.
To investigate whether the loss of PAR1 kinase activity is responsible for the observed stunted phenotype in the mutant, the researchers treated the mutant plants with different nucleotides in vitro. The treatment partially rescued the stunted growth, suggesting that the kinase activity of PAR1 is at least partially responsible for the mutant phenotype. Additionally, RNA-seq analysis comparing the par1 mutants with wild-type plants revealed significant alterations in the jasmonic acid (JA) biosynthetic and signaling pathways. This misregulation of the JA pathway was confirmed by high levels of JA in the mutant plants.
The findings of this study suggest a connection between nucleotide metabolism and the regulation of plant architecture, with particular emphasis on the role of transposon-mediated epigenetic regulation of gene expression. Notably, this research uncovers a molecular mechanism through which nucleotide and JA metabolism influence plant development (Figure). While DNA methylation is typically associated with gene silencing (Xu et al. 2022), this study sheds light on the complex dynamics between high DNA methylation and high gene expression, providing insights into previously unexplained phenomena. However, the precise relationship between nucleotide metabolism and JA metabolism still remains an open question that warrants further investigation.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Li Q, Gent JI, Zynda G, Song J, Makarevitch I, Hirsch CD, Hirsch CN, Dawe RK, Madzima TF, Mc Ginnis KM, et al RNA-directed DNA methylation enforces boundaries between heterochromatin and euchromatin in the maize genome. Proc Natl Acad Sci U S A. 2015:112(47):14728–14733. 10.1073/pnas.151468011226553984 PMC 4664327 · doi ↗ · pubmed ↗
- 2Li R, Xu Y, Xu Q, Tang J, Chen W, Luo Z, Liu H, Li W, Yan J, Springer NM, et al (2025) An epiallele of a gene encoding Pfk B-type carbohydrate kinase affects plant architecture in maize. Plant Cell. 37(1):koaf 017. 10.1093/plcell/koaf 017PMC 1178088639823309 · doi ↗ · pubmed ↗
- 3Schnable PS, Ware D, Fulton RS, Stein JC, Wei F, Pasternak S, Liang C, Zhang J, Fulton L, Graves TA, et al The B 73 maize genome: complexity, diversity, and dynamics. Science. 2009:326(5956):1112–1115. 10.1126/science.117853419965430 · doi ↗ · pubmed ↗
- 4Slotkin RK, Martienssen R. Transposable elements and the epigenetic regulation of the genome. Nat Rev Genet. 2007:8(4):272–285. 10.1038/nrg 207217363976 · doi ↗ · pubmed ↗
- 5Wicker T, Sabot F, Hua-Van A, Bennetzen JL, Capy P, Chalhoub B, Flavell A, Leroy P, Morgante M, Panaud O, et al A unified classification system for eukaryotic transposable elements. Nat Rev Genet. 2007:8(12):973–982. 10.1038/nrg 216517984973 · doi ↗ · pubmed ↗
- 6Witte C-P, Herde M. Nucleotide metabolism in plants. Plant Physiol. 2020:182(1):63–78. 10.1104/pp.19.0095531641078 PMC 6945853 · doi ↗ · pubmed ↗
- 7Wittmeyer K, Cui J, Chatterjee D, Lee T-f, Tan Q, Xue W, Jiao Y, Wang P-H, Gaffoor I, Ware D, et al The dominant and poorly penetrant phenotypes of maize unstable factor for orange 1 are caused by DNA methylation changes at a linked transposon. Plant Cell. 2018:30(12):3006–3023. 10.1105/tpc.18.0054630563848 PMC 6354275 · doi ↗ · pubmed ↗
- 8Wong AH, Gottesman II, Petronis A. Phenotypic differences in genetically identical organisms: the epigenetic perspective. Hum Mol Genet. 2005:14(suppl_1):R 11–R 18. 10.1093/hmg/ddi 11615809262 · doi ↗ · pubmed ↗