A new species of genus Chiloschista (Aeridinae, Vandeae, Epidendroideae, Orchidaceae) from Sumatra Island, Indonesia

Abstract
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
 Figure 1
Figure 1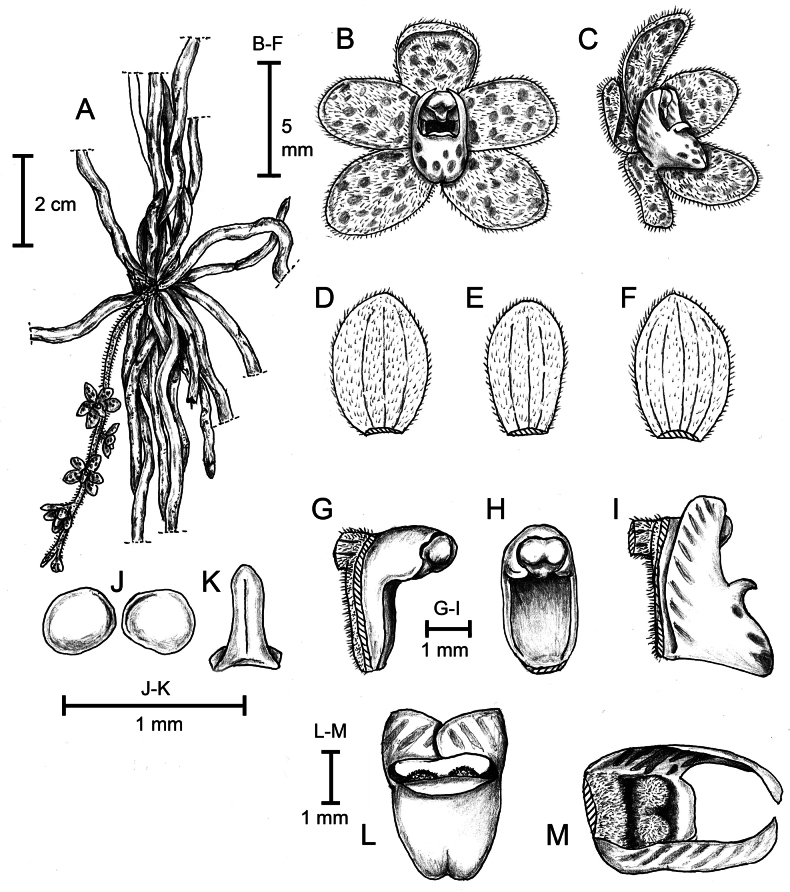 Figure 2
Figure 2 Figure 3
Figure 3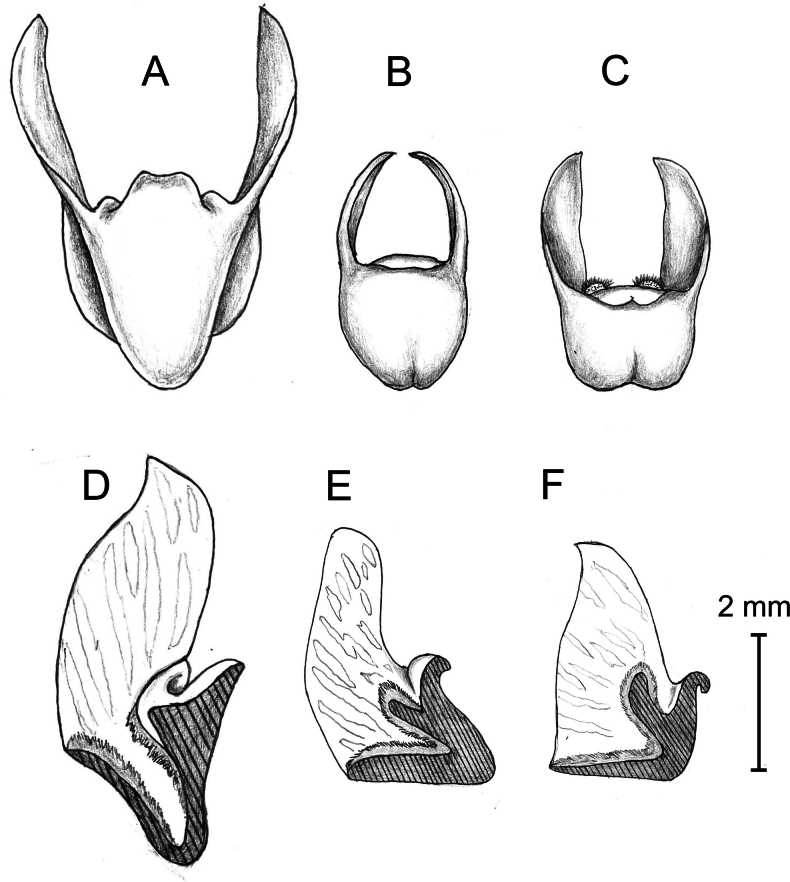 Figure 4
Figure 4| Characters | |||
|---|---|---|---|
| Flower colour | Sepals and petals yellowish cream or yellow with orange or reddish spots, lip yellowish cream with reddish or orange spots on their sac and side lobes with pale reddish orange stripes | Sepals and petals yellowish cream or yellow with orange or reddish spots, lip white or cream with pale reddish or orange spots on their sac and side lobes with pale reddish orange stripes | Sepals and petals yellow with orange-brown blotches, lip white on their sac and side lobes with reddish-orange stripes |
| Dorsal sepal | Oblong-elliptic, 5.0–6.0 mm long × 4.0 mm wide, obtuse to rounded, both surfaces pubescent but abaxial with shorter hairs | Elliptic, 5.0–6.0 × 3.0–3.5 mm, obtuse to rounded, adaxial surface pubescent, abaxial with scattered shorter hairs | Ovate, 5.0–6.0 × 3.5–5.0 mm, obtuse to acute, only adaxial surface pubescent |
| Lateral sepals | Obliquely oblong-elliptic, 6.0 mm long × 4.0–5.0 mm, obtuse, both surfaces pubescent but abaxial with shorter hairs | Elliptic, 4.0–5.0 mm × 3.0–3.5 mm, obtuse to rounded, adaxial surface pubescent, abaxial with scattered shorter hairs | Broadly elliptic to broadly ovate, 5.0–6.0 × 3.5–5.0 mm, obtuse to acute, only adaxial surface pubescent |
| Petals | Oblong-obovate, 5.0–6.0 mm long × 3.0–4.0 mm wide, truncate to rounded, both surfaces pubescent but abaxial with shorter hairs | Broadly elliptic to ovate, 4.0–4.5 × 3.0–3.5 mm, obtuse to rounded, adaxial surface pubescent, abaxial with scattered shorter hairs | Ovate-orbicular, 4.0–5.0 × 3.0–4.5, obtuse, adaxial surface sparsely pubescent |
| Side lobes | Obliquely oblong, slightly falcate, | Triangular, relatively straight, | Obliquely oblong, slightly falcate, c. 2 mm wide at base and narrowed gradually to about 0.9 mm near apex, acute to obtuse |
| Front lobe | Broadly subtrapezoid, short, | Broadly subtrapezoid, short, | Broadly triangular, short, |
| Lip’s sac | Ovate to subrectangular from front view, forming a “V” shape in longitudinal section view with a narrower angle (45–50°), | Subrectangular from front view, forming an “L” shape in longitudinal section view with a wider angle (90°), | Triangular from front view, forming a “V” shape in longitudinal section view with a narrower angle (45–50°), |
| Internal lip’s sac ornament | An oblong fleshy thickening arises from basal to middle of the internal front wall, split into 2 lobed hairy callus that rises to sac opening, creating a small narrow cavity between apex of the hairy callus and the thick curved front lobe | A large and high oblong fleshy thickening arises from basal to middle of the internal front wall, split into 2 lobed long protruding hairy callus that rises to sac opening, creating a cavity between apex of the hairy callus and the thick curved front lobe | Internal callus absence |
| 1 | Roots very flattened with a glabrous inflorescence’s peduncle |
|
| – | Roots more or less terete with a hairy inflorescence’s peduncle |
|
| 2 | Sepals and petals narrowly elliptic, the lip’s sac conical with pointed apex |
|
| – | Sepals and petals broadly elliptic, the lip’s sac hemispherical with rounded or slightly bilobed apex |
|
| 3 | Lip’s sidelobes obliquely quadrangular with sac apex rounded |
|
| – | Lip’s sidelobes oblong with sac apex slightly bilobed |
|
| 4 | Lip’s sac has a “V” shape in longitudinal section view with a narrow angle of about 45–50° |
|
| – | Lip’s sac has an “L” shape in longitudinal section view with a wide angle of about 90° |
|
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsPlant and animal studies · Plant and Fungal Species Descriptions · Plant Parasitism and Resistance
Introduction
The Indonesian archipelago is known to be one of the most important orchid diversity hotspots in the world, and it continues to generate new species discoveries, which indicates that there are still many potential areas that need to be explored (e.g., Metusala 2017a; Metusala and Supriatna 2017; Metusala 2019a, 2019b; Metusala and O’Byrne 2020; Saputra et al. 2020; Metusala et al. 2021; Metusala 2024). Recent botanical explorations in Indonesia have usually focused on a few popular, orchid taxa, while groups such as mycoheterotropic and leafless orchids have been comparatively neglected, presumably due to their less attractive habitus, often cryptic growth habit and small flower size (Metusala and Supriatna 2017; Metusala 2020a; Suetsugu et al. 2021; Panday et al. 2022).
The leafless orchid genus Chiloschista Lindl. (1832: 1522) was established in 1832 with C.usneoides (D.Don) Lindl. (1832: 1522) as its type, and there are now 30 accepted species distributed from the Indian subcontinent through to Southeast Asia and Australia (Dalstrom and Kolanowska 2020; POWO 2024). In previous treatments, the genus was classified under various subtribes, such as Sarcochilinae (Schlechter 1926), Aeridinae (Dressler 1993), and Phalaenopsidinae (Szlachetko 2003). However, recent phylogenetic studies showed that Chiloschista is a monophyletic genus, clustered with Phalaenopsis, within the subtribe Aeridinae of the tribe Vandeae (Zou et al. 2015; Liu et al. 2023).
Species of this genus are characterized by monopodial growth, typically as epiphytic or lithophytic herbs. Plants are often seen as a cluster of numerous terete or flattened photosynthetic roots that radiate from a very short stem. Chiloschista flowers are either ephemeral or last up to several days, 3-lobed, saccate or spurred, and usually have a thickened hairy internal callus. Their columns are subterete and short, but have a rather long foot. The anther caps have 2 filiform setae and 4 pollinia in 2 closely appressed pairs (Cockburn et al. 1985; Chen and Wood 2009; Wood 2014; Dalstrom and Kolanowska 2020).
Prior to this article, Indonesia has 4 accepted species: C.javanica Schltr. (1919: 275) is recorded only from Java Island; C.phyllorhiza (F.Muell) Schltr. (1921: 492) is distributed across Java, the Lesser Sunda Islands, and Sulawesi; C.taeniophyllum (J.J.Sm.) Schltr. (1921: 492) is endemic to Maluku (Ambon Island and Banda Islands); and C.treubii (J.J.Sm.) Schltr. (1921: 492) that is also endemic to Maluku (Seram Island and Wokam Island of the Aru Islands Regency) (POWO 2024). There is no prior record of any Chiloschista species from Sumatra, Borneo, and New Guinea Islands.
During a botanical inventory conducted by the author in 2019, several living specimens of Chiloschista were found growing on semi-open coffee plantation close to the forest in Aceh Province, in the northernmost region of Sumatra Island. Chiloschista roots usually turn darker when wet and look like the color of tree bark, which makes them difficult to find, such that their small but bright flowers are often important to their detection. Several plants, including individuals with flowers, were collected as herbarium and living specimens for Purwodadi Botanic Gardens in East Java. Further observations of the flowering specimens found that this taxon represents an undescribed species with flower characteristics morphologically similar to C.javanica and C.sweelimii. Here I describe it as a new species, as well as the first record of this genus on Sumatra Island. A key to the five species of Chiloschista in Indonesia is also included.
Materials and methods
Morphological measurements were conducted with a loupe and a ruler accurate to 0.5 mm. Environmental data were collected using thermohygrometer and lux–meter. The specimens were also compared with closely related species (C.javanica and C.sweelimii). Observation and morphological studies have been done by examining the relevant literature sources, illustrations, living and herbarium specimens, and photographs. These literature sources included protologues and descriptions of the relevant taxa: C.javanica (Smith 1905a), C.sweelimii (Teck 2016), C.taeniophyllum (Smith 1905b), C.treubii (Smith 1912), and C.phyllorhiza (Mueller 1866; Smith 1939). Photographs were taken with a Sony DSC-W70. Detailed morphological observations were conducted using stereomicroscope Olympus SZX7 with camera EP50. Terminology for morphological description follows Harris and Harris (2001). Conservation status was assessed using the IUCN Red List Category and Criteria. The Extent of Occurrence (EOO) and Area of Occupancy (AOO) were estimated based on GeoCAT (https://geocat.iucnredlist.org/). Six specimens of Chiloschista sp. from Aceh were used for examinations (RIO 9117; 9118; 9119; 9121; 9123; 9124).
Additional examined specimens
-
Chiloschistasweelimii. Malaysia • Malay Peninsula; Lim, S.L. s.n.; K 000891272 • Malay Peninsula; Ong FRI 75458 (KEP).
-
Chiloschistajavanica. Indonesia • Java; Docters van Leeuwen s.n., L 1500077 • Java; leg. ign. s.n.; L 0264545 • Java; Docters van Leeuwen 2402; BO 0057026 • Java; Docters van Leeuwen 2402; BO 0057027 • Java; leg. ign. s.n.; BO 0057030 • Java; Docters van Leeuwen s.n.; BO 0057031 • Java; Docters van Leeuwen s.n.; BO 0057037 • Java; leg. ign. s.n.; BO 0057038.
Taxonomic treatment
Chiloschista
tjiasmantoi
Taxon classificationPlantae AsparagalesOrchidaceae
Metusala sp. nov.
F384924D-3987-587C-9228-9507231E449E
urn:lsid:ipni.org:names:77356634-1
Figs 1A, B, 2, 3, 4B, E
Type.
Indonesia • Sumatra: Aceh Province, c. 900 m, RIO 9118 (holotype, BO!) (detailed localities are not shown here for conservation purpose).
Diagnosis.
Chiloschistatjiasmantoi is morphologically similar to C.javanica, but differs in having oblong-obovate petals (vs. broadly elliptic to ovate petals), narrowly oblique oblong side lobes with truncate to obtuse apex (vs. relatively straight triangular side lobes with obtuse apex), a lip sac that has a “V” shape in longitudinal section view with a narrow angle of about 45–50° (vs. a lip sac that has an “L” shape in longitudinal section view with a wide angle of about 90°), and a very narrow cavity between the apex of the hairy callus and the thick curved front lobe of the lip (vs. a rather broad cavity).
Description.
Epiphytic herb. Roots numerous, spreading, terete to slightly flattened, 2.0–3.5 mm in diameter, greyish-green when wet and becoming grayish-white when dry, grow radially from a short stem as the central, and mature individuals can grow elongated to reach more than 30.0 cm. Stem reduced, very short, erect, simple 2.0–4.0 mm long, up to 3.0 mm in diameter, densely covered with dry stem bracts. Stem bracts are triangular active, and persistent, and encircle the stem tightly. Leaves one or two, 4–7 mm long, deciduous, unseen in cultivation. Inflorescence axillary, arising among the roots gap, pendulous, up to 31.0 cm long in total, peduncle c. 2.0 mm in diam. near base, terete, densely covered with short white hairs, purplish near base and becoming purplish green toward apex, sometimes branched at the base, up to 30 flowered per rachis, flowers arranged spirally in a slightly zig-zag pattern and open simultaneously, each flower can last up to 5 days; flower bracts triangular, 2.0–3.0 mm long × 1.5–2.0 mm wide, acuminate to caudate, pubescent, greenish and soon becoming brown when old. Pedicel and ovary about 2.0 mm long, terete, brownish-green to purplish-green, covered with whitish hairs. Flower rather thin-textured, 1.0–1.2 cm high × 1.0–1.2 cm wide, open widely, sepals and petals yellowish cream or yellow with orange or reddish spots, labellum yellowish cream with reddish or orange spots on their sac, column yellowish green with orange tinge on its foot. Dorsal sepal oblong-elliptic, 5.0–6.0 mm long × 4.0 mm wide, obtuse to rounded, both surfaces pubescent. Petals oblong-obovate, 5.0–6.0 mm long × 3.0–4.0 mm wide, truncate to rounded, both surfaces pubescent. Lateral sepals obliquely oblong-elliptic, 6.0 mm long × 4.0–5.0 mm wide, obtuse, both surfaces pubescent. Lip immobile, minutely papillose externally, 3-lobed, deeply saccate, indistinctly canaliculated ventrally, 2.8–3.2 mm long, 4.0–4.5 mm high (from side lobes apex to basal part of sac), 2.0–2.5 mm wide at front; side lobes erect to slightly curved inwards, obliquely oblong to rather falcate, c. 2.0 mm long × c. 1.5 mm wide near base and narrowed gradually to about 1.0 mm near apex, apex truncate to rounded, yellowish cream with red streaks on internal surface; front lobe subtrapezoid, short c. 1.0–1.5 mm × 1.5 mm, apex truncate to slightly emarginate, curved; sac ovate to subrectangular from front view, “V” shape in longitudinal section view, 2.0–2.5 mm long × 2.0–2.3 mm wide, apex truncate or retuse or slightly bilobed, yellowish cream with pale reddish or pale orange spots on external surface around the apex; callus a fleshy thickening arise from basal to middle of the internal front wall, split into two oblong hairy callus that rises up to sac opening, creating a small narrow cavity between apex of the hairy callus and the thick curved front lobe of the lip; column short 1.5–2.0 mm long (excluding the anther cap), foot about 2.2–2.5 mm long; anther cap cucullate, yellowish or cream, c. 1.5 mm × 1.5 mm; pollinarium two unequal globose on a narrowly linear to triangular stipe, yellow. Fruit not seen.
Distribution, habitat and phenology.
Based on the existing data, the distribution of Chiloschistatjiasmantoi may be restricted to Aceh Province, the most northern part of Sumatra Island. This new species is currently only known from five locations in two different regencies at elevation ranging from 700–1000 m. The populations of this species were mostly found growing epiphytically on old coffee trees (Coffea spp.) and shade trees (Leucaena spp.) in the local coffee plantations, together with Vandapumila (Orchidaceae), in a windy and semi–opened wet habitat with medium sunlight intensity. Flowering recorded in mid-July, early November to late December.
Etymology.
The specific epithet “tjiasmantoi” honors Wewin Tjiasmanto, the chairman of Tjiasmanto Conservation Fund and a philanthropist concerned with the Indonesian plant conservation.
Cultivation.
Chiloschistatjiasmantoi seems rather difficult to cultivate at lower elevations (300 m. a.s.l). However, it was successfully grown by attaching it to a slab of tree fern with a top dressing of moss to prevent the roots from drying out, under a light intensity of about 30–75% with good air circulation and humidity levels of about 80% or more.
Discussion.
This new species is morphologically similar to C.javanica (Fig. 1C, D, 4C, F) and C.sweelimii (Fig. 4A, D). However, there is a geographical separation among these three species. C.tjiasmantoi is thus far only recorded from Aceh Province, the northernmost region of Sumatra Island; C.sweelimii occurs in the Malay Peninsula and Vietnam, and C.javanica is endemic to Java Island.
Chiloschistatjiasmantoi sp. nov. (A, B) and Chiloschistajavanica (C, D) A inflorescence B flower, oblique view. C flower, front view D flower, oblique view. Photos by Destario Metusala.
Chiloschistatjiasmantoi differs from C.javanica in having narrower oblong-obovate petals, narrowly oblique oblong side lobes with truncate to obtuse apex, a lip sac that has a “V” shape in longitudinal section view with a narrow angle of about 45–50°, a narrow cavity between the apex of the hairy callus and the thick curved front lobe of the lip. Meanwhile, C.javanica has broadly elliptic to ovate petals, relatively straight triangular side lobes with obtuse apex, a lip sac that has an “L” shape in longitudinal section view with a wide angle of about 90°, and a rather broad cavity between the apex of the hairy callus and the thick curved front lobe of the lip.
Chiloschistatjiasmantoi sp. nov. A habitus with inflorescence B flower, natural shape, front view C flower, natural shape, oblique view D dorsal sepal, flat shape E petal, flat shape F lateral sepal, flat shape G column and foot, oblique view H column and foot, front view I column and lip, side view J pollinia K stipe and viscidium L lip, above view M lip interior, back view. Line drawing by Destario Metusala from RIO 9118.
Furthermore, C.tjiasmantoi differs from C.sweelimii in having sepals and petals with hairs on both sides, oblong-obovate petals, a broadly oblong-elliptic dorsal sepal, basal front lobe without any callus, an ovoid to subrectangular lip’s sac from front view with a truncate to slightly bilobed apex, and an oblong fleshy thickening that arises from basal to middle of the internal front wall. This thickening then divides into a 2-lobed hairy callus that rises near the sac opening, creating a cavity between the apex of the hairy callus and the thick curved front lobe. In contrast, C.sweelimii has sepals and petals with hairs only on their adaxial surface, ovate to orbicular petals, an ovate dorsal sepal, basal front lobe with large callus on either side (each c. 1.5 × 1.0 mm), a triangular shaped lip’s sac from front view with pointed obtuse apex, and the absence of prominent internal callus inside the lip’s sac (Teck 2016). A comparison between the new species and its morphological allies is shown in Table 1.
Chiloschistatjiasmantoi sp. nov. Flowering plants in situ. Photos by Alfajaruddin.
This new species will be the fifth Chiloschista species in Indonesia, and the first record of this genus on Sumatra Island. With the exception of C.phyllorhiza, the species are endemic to the country and appear to have restricted distributions. Although the flowers of some species appear nearly identical in appearance, the internal structure of the lip can be very different, which suggests this would be a good key character for further identification of this genus (Gyeltshen et al. 2019; Dalstrom and Kolanowska 2020). A deeper investigation into its floral morphological variation needs to be carried out in the future, as this is essential to support species delimitation of Indonesian Chiloschista (Metusala 2020b; Metusala et al. 2020).
Lip of three Chiloschista species AC.sweelimii, natural shape, front view BC.tjiasmantoi, natural shape, front view CC.javanica, natural shape, front view DC.sweelimii, longitudinal section, flat shape, side view EC.tjiasmantoi, longitudinal section, flat shape, side view FC.javanica, longitudinal section, flat shape, side view. A and D redrawn after Ong Poh Teck in Teck (2016). Drawn by Destario Metusala.
Conservation.
Chiloschistatjiasmantoi was recorded from five locations in Aceh Province. Based on currently available data, the Extent of Occurrence (EOO) of this species is 117.01 km^2^ with user-defined cell width = 2 km (criterion B1: < 5,000 km^2^) and Area of Occupancy (AOO) value of 20.00 km^2^ (criterion B2: < 500 km^2^). Their natural habitat is threatened by land conversion, especially from large-scale coffee plantations. Existing populations were estimated to be less than 2,500 mature individuals (≤ 250 in most subpopulations) where they mostly found attached to the branches of coffee trees and its shade trees (Leucaena spp.). Unfortunately, coffee plantations provide a vulnerable habitat for this new orchid species as they can be pruned or cut down at any time. Moreover, many coffee farmers believe that this orchid is a harmful parasitic plant and eradicate them by clearing the coffee branches from any epiphytic plants. This species has also been found traded on domestic online platforms as an ornamental orchid, although currently in small quantities as its habit and small flowers are considered less attractive to Indonesian hobbyists. In many cases, unsustainable commercial harvesting may have a significant impact on the viability of wild orchid populations (Hinsley et al. 2018). Therefore, I consider this species to likely be in the Endangered category B1 and B2+ab (i,ii,iii,iv), C2+a (i) (IUCN Red List Categories and Criteria).
It has been well-documented that ecosystems in the highland of Aceh Province are severely affected by climate change, particularly through increased temperatures (Schroth et al. 2015). This could also have a major impact on various sensitive plant species, such as many highland epiphytic orchids, including natural populations of Chiloschistatjiasmantoi. Therefore, conservation-based researches are needed to determine the vulnerability of these species to climate change-related pressures, especially drought stress (Al Farishy et al. 2017; Arimy et al. 2017; Metusala 2017b; Metusala et al. 2017; Suffan et al. 2021; Ishmah et al. 2021; Trimanto et al. 2023).
Key to the species of Chiloschista in Indonesia
**: **
Supplementary Material
XML Treatment for Chiloschista tjiasmantoi
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Al Farishy DD Nisyawati Metusala D (2017) Characterization anatomical leaf blade five species Nepenthes from Kerinci Seblat National Park, Kerinci regency, Jambi Province. In: Mart T, Triyono D, Sugeng KA (Eds) AIP Conference Proceedings 1862, 030115. 10.1063/1.4991219 · doi ↗
- 2Arimy NQ Nisyawati Metusala D (2017) Comparison of leaf anatomy on some Nepenthes spp. (Nepenthaceae) from highland and lowland habitat in Indonesia. In: Mart T, Triyono D, Sugeng KA (Eds) AIP Conference Proceedings 1862, 030111. 10.1063/1.4991215 · doi ↗
- 3Chen X Wood JJ (2009) Chiloschista.Flora of China 25: 470–471. http://flora.huh.harvard.edu/
- 4Cockburn W Goh CJ Avadhani PN (1985) Photosynthetic carbon assimilation in a shootless orchid, Chiloschistausneoides (DON) LDL.Plant Physiology 77(1): 83–86. 10.1104/pp.77.1.8316664034 PMC 1064461 · doi ↗ · pubmed ↗
- 5Dalstrom S Kolanowska M (2020) A new yellow-flowered Chiloschista (Orchidaceae: Aeridinae) from Thailand.Lankesteriana 20(2): 241–248. 10.15517/lank.v 20i 2.43454 · doi ↗
- 6Dressler RL (1993) Phylogeny and classification of the orchid family. Dioscorides Press, 1–330.
- 7Gyeltshen C Dalstrom S Gyeltshen N Tobgay K (2019) A new spotted Chiloschista (Orchidaceae: Aeridinae) From Bhutan.Lankesteriana 19(1): 23–29. 10.15517/lank.v 19i 1.37030 · doi ↗
- 8Harris JG Harris MW (2001) Plant identification terminology: an illustrated glossary. Spring Lake Publication, 1–206.