Pre- and post-traumatic boric acid therapy prevents oxidative stress-mediated neuronal apoptosis in spinal cord injury
Turan Kandemir, Ibrahim Sogut, Zeki Serdar Ataizi, Betul Can, Aysegul Oglakci-Ilhan, Dilek Burukoglu-Donmez, Gungor Kanbak

TL;DR
This study shows that boric acid, given before or after spinal cord injury in rats, reduces oxidative stress and prevents nerve cell death.
Contribution
The novel finding is that pre- and post-traumatic boric acid application effectively mitigates oxidative stress and apoptosis in spinal cord injury.
Findings
Boric acid reduced oxidative stress markers like MDA, TOS, and OSI after spinal cord injury.
Boric acid application decreased apoptosis markers Cytc and Casp3 in injured spinal cord tissues.
Histological damage, including cell and myelin damage, was reduced with boric acid treatment.
Abstract
In our study, the neuroprotective efficacy of pre- and post-traumatic applications of boric acid (BA) in rats with experimentally induced spinal cord injury (SCI) was investigated. The experimental animals were divided into four groups: control group (C), SCI group (SCI), BA-treated group before SCI (BA+SCI), and BA-treated group after SCI (SCI+BA). Forty-eight hours after SCI, biochemical levels of malondialdehyde (MDA), total oxidant status (TOS), total antioxidant status (TAS), oxidative stress index (OSI), and cytochrome c (Cytc) and caspase-3 (Casp3) expressions were measured in the spinal cord tissues and were examined histologically. After SCI, oxidative stress markers, such as MDA, TOS, and OSI, and apoptosis markers Cytc and Casp3 showed an increase in levels compared to Group C. The oxidative stress markers that increased after SCI decreased with BA+SCI application, while…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
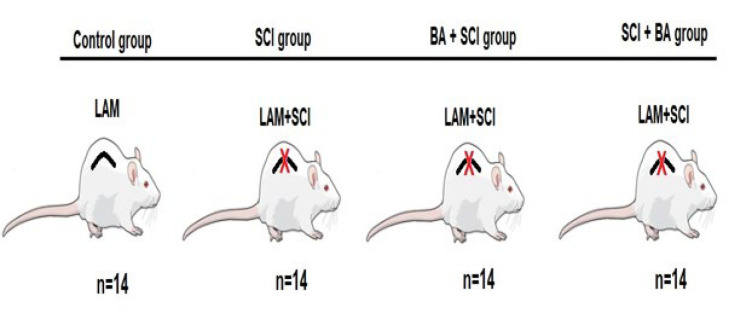 Figure 1
Figure 1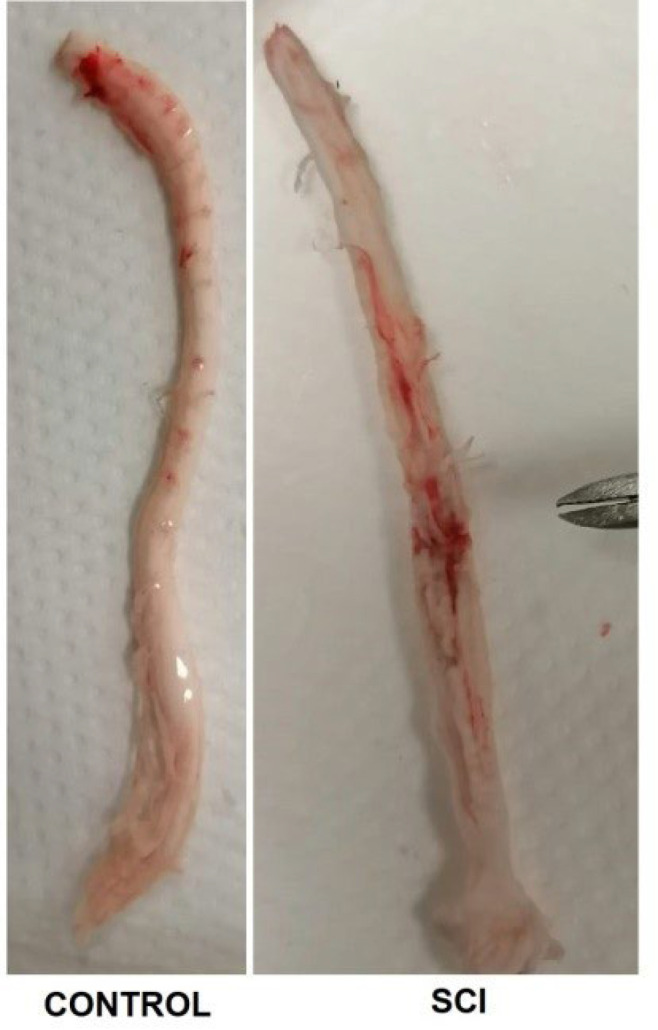 Figure 2
Figure 2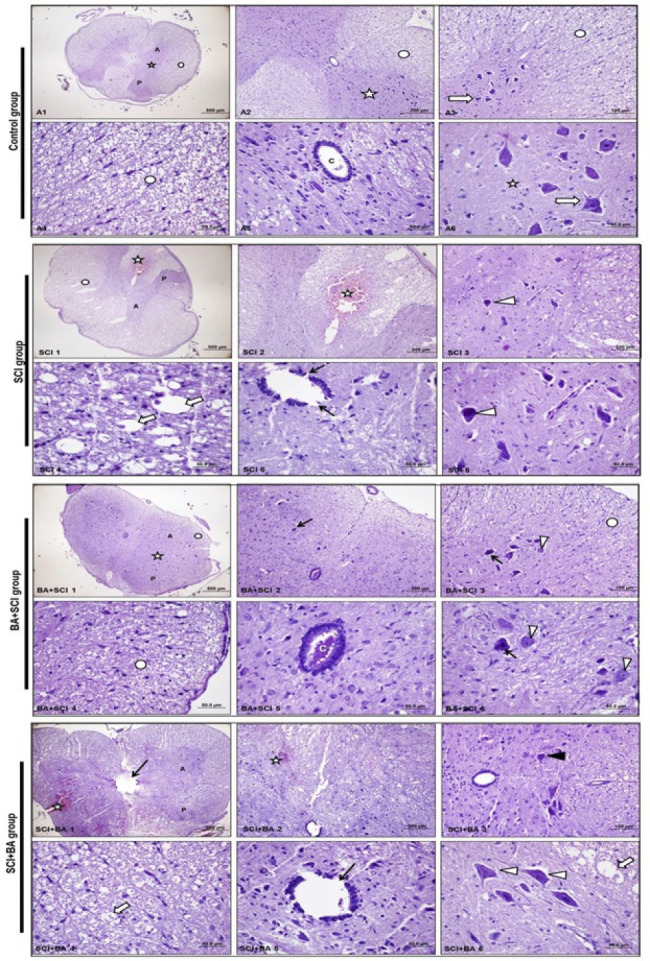 Figure 3
Figure 3| Primer Name | Forward Primer Sequence (5′-3′) | Reverse Primer Sequence (3′-5′) |
|---|---|---|
| Caspase-3 ( | TGACTGGAAAGCCGAAACTC | GTAGAGTAAGCATACAGGAAGTCAG |
| Cytochrome-c ( | AAGGGAGGCAAGCATAAGAC | ATTCTCCAAATACTCCATCAGGG |
| GAPDH | CCTCGTCCCGTAGACAAAATG | TGTAGTTGAGGTCAATGAAGGG |
| Groups → Biochemical Parameters ↓ | Control | SCI | BA+SCI | SCI+BA | p |
|---|---|---|---|---|---|
| MDA | 13.94±3.23 | 17.69±2.6 | 11.25±3.55 | 15.40±3.02 | SCI & BA+SCI ** |
| TOS | 41.50±8.4 | 62±11.16 | 48.75±10.69 | 54.5±7.3 | Control & SCI ** |
| TAS | 2.86±1.17 | 2.06±0.49 | 3.23±1.4 | 2.34±0.28 | No significant |
| OSI | 16.17±7.62 | 31.25±8.21 | 17.36±6.76 | 23.59±4.39 | Control & SCI *** |
| Groups → Molecular Parameters ↓ | Control | SCI | BA+SCI | SCI+BA | p |
| Cytochrome c | 1.0 | 2.183±0.58 | 1.461±0.36 | 1.233±0.61 | Control & SCI *** |
| Caspase 3 | 1.0 | 2.02 | 1.8 | 1.42 | Control & SCI ** |
| Groups→ Histological Parameters ↓ | Control | SCI | BA+SCI | SCI+BA | p |
| Cell damage/Ischemia | 0 (0-0) | 3 (2-3) | 1 (1-2) | 2 (1-2) | Control & SCI **** |
| Myelin damage/Vacuolization | 0 (0-0) | 3 (2-3) | 0 (0-1) | 2 (1-2) | Control & SCI **** |
| Ependymal cell damaged | 0 (0-0) | 2 (2-3) | 0 (0-1) | 1 (1-2) | Control & SCI **** |
| Hemorrhage | 0 (0-0) | 3 (2-3) | 0 (0-1) | 0 (0-1) | Control & SCI **** |
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsMetabolism and Genetic Disorders
Introduction
Spinal cord injury (SCI) is one of the most important health problems of today as it is a serious neurological problem that affects modern society physically, psychosocially, and economically. In many studies, traffic accidents are the first cause of SCI (1, 2). Other causes include work accidents, falls from height, gunshot wounds, and sports injuries. Although the worldwide prevalence of SCI is unknown, a meta-analysis found that the incidence of traumatic SCI was 26.48 per million people (3). Understanding the pathophysiology of SCI is important in designing a therapeutic approach. The pathophysiological process of SCI consists of two stages: primary injury and secondary injury. The mechanical damage that initially occurs in SCI is referred to as primary damage and consists of acute, physiologic, and structural disruption of axons (4). Although the primary damage is limited to the injury site, it triggers the onset of many subsequent physiopathological processes, leading to secondary damage. Secondary damage can last for months. During this period, the blood-spinal cord barrier is disrupted, and apoptosis, increased peripheral inflammatory cells, reactive oxygen/nitrogen species (ROS/RNS) generation, intracellular calcium accumulation, activation of proteases and caspases, lipid peroxidation, and glutamate excitotoxicity such as compression, tearing, shearing, and stretching are observed (5-7).
As mentioned above, overproduction of ROS exacerbates oxidative stress, inflammatory response, and various cell death pathways, which play a crucial role in the secondary injury process (8, 9). Enzymatic and nonenzymatic antioxidant mechanisms inactivate free radicals formed under physiological conditions. Enzymatic antioxidants (e.g., catalase (CAT), superoxide dismutase (SOD), glutathione peroxidase (GPx), etc.) cannot respond to the excessive increase of free radicals in the tissue after trauma to the same degree. There is a need to increase the concentration of nonenzymatic antioxidants (e.g., vitamin C, vitamin E, glutathione) in the body that act as radical scavenging agents (10). Under physiological conditions, mitochondria are the largest source of cellular ROS (e.g., superoxide, hydroxyl radicals, hydrogen peroxide, lipid peroxide, and peroxynitrite) as they utilize oxygen by cells during oxidative phosphorylation. However, SCI causes mitochondrial dysfunction, further increasing ROS production (11). Furthermore, ROS uptake by microglia and leukocytes is further increased after SCI (12). Under normal conditions, ROS are balanced by various biochemical processes, such as antioxidant systems. However, after SCI, increased ROS formation and also this imbalance between free radicals and antioxidants leads to the formation of oxidative stress, especially in lipids, proteins, and nucleic acids (13-15). In addition, the blood-brain barrier is disrupted due to endothelial damage caused by free oxygen radicals. As a result, there is an accumulation of ROS at the site of injury. Free radical damage occurs more frequently due to the low superoxide dismutase (SOD), catalase, and glutathione peroxidase (GPx) activities in this region (16). Apoptosis is responsible for cell death after traumatic SCI. Apoptotic processes are triggered by the action of reactive oxygen species (ROS). The increased amount of ROS increases mitochondrial membrane permeability. It leads to the release of pro-apoptotic molecules, such as cytochrome c, from mitochondria and an increase in the level of Caspase-3 from the cysteine protease family, causing apoptosis (17-19). The mechanism of apoptosis in spinal cord injuries depends on caspase-3 and calpain. Following injury, apoptosis mediated by caspase, cytochrome-c release into the cytosol, and an increase in the bax/bcl-2 ratio are observed in spinal cord lesions. Thus, apoptosis is observed in neurons and glial cells (20).
Recent studies have revealed that one of the main functions of mitochondria is to regulate apoptosis. Neurons are very rich in mitochondria and are the cell type with the highest energy consumption; hence, they have a higher demand for mitochondrial ATP production. Mitochondria are the main source of ROS/RNS production and, therefore, a potential therapeutic target (21). Studies have shown free radical-mediated mitochondrial dysfunction as a significant contributor to secondary damage following SCI. Oxidative damage and mitochondrial dysfunction are interrelated processes that also catalyze other secondary injury events (22).
The most effective therapeutic intervention for SCI is the prevention of secondary injuries, especially by using various antioxidants to reduce the reduction of oxygen free radicals. (13, 16). Boric acid (BA), which is used in this study, is an important trace element for organisms. BA shows Lewis acid properties, forms complexes with glycoproteins and glycolipids containing hydroxyl groups, and has Ca+ chelator properties. It also has antioxidant, antiapoptotic, and anticancer properties as well as oxidative stress-reducing properties (13, 14, 18, 19). Previously, our research team demonstrated the oxidative stress-reducing effect of BA in rats with traumatic brain injury (13). In this study, we investigated the protective role of pre and after-boric acid administration in oxidative stress-mediated mitochondrial dysfunction and neuronal apoptosis.
Materials and Methods
In the current experiment, 56 male mature Sprague Dawley rats weighing 250-260 g were used. The rats were kept in standard animal care cages, with a 12-hour day/night cycle, at a constant temperature of 22 ^°^C±30 ^°^C and a constant humidity (55%±5%). All experimental procedures were conducted in accordance with the guidelines of Eskisehir Osmangazi University Animal Ethics Committee (Protocol no: 847-1/2022) and with the Guide for the Care and Use of Laboratory Animals prepared by the National Academy of Sciences.
The rats were randomly divided into four groups. Control group (C; n=14): Laminectomy without any injury to the spinal cord was performed. SCI (n=14): Laminectomy and SCI was performed. BA+SCI group (n=14): Intraperitoneal (IP) BA was administered (100 mg/kg)(13, 14) one hour before laminectomy and induction of SCI. SCI+BA group (n=14): BA was administered intraperitoneally (100 mg/kg) one hour after laminectomy and inducing SCI (Figure 1).
Due to limited access to spinal cord tissue in experimental SCI, the number of groups was formed as n=14. Seven experimental animals were used for biochemical studies in each group, and the other seven were used for histological and molecular studies.
Preparation and implementation of BA
BA solution was freshly prepared in 0.5 ml saline solution (SS) containing 100 mg/kg doses of BA (Merck, Darmstadt, Germany) just before the study. According to the groups’ content, BA was administered via the IP route one hour before SCI and one hour after SCI(23-26).
Traumatic SCI model and procedure
The experimental animals were subjected to the clip compression model described by Rivlin and Tator to induce spinal trauma (27). During the surgical procedure, a Yasargil FE 720K (with a closing force of 110 g, Aesculap/Germany) clip was used. The surgical procedure was performed on the animals under general anesthesia with intraperitoneal xylazine hydrochloride (10 mg/kg) and ketamine hydrochloride (90-100 mg/kg). After anesthesia, the rats were placed on the operation table in the prone position, the dorsal region was shaved, and local antisepsis was applied with polyvinyl iodine. A single-level total laminectomy was performed by placing the laminae between dorsal 7-10 vertebrae through a midline surgical incision. After careful placement without damaging the dura, the aneurysm was cord-clipped with an aneurysm clip. After one minute, the clip was removed, and the surgical procedure was completed. Muscles and skin were closed in layers with 3-0 silk sutures after all bleeding was stopped. It was accepted that SCI was successfully established as the hindlimbs were paralyzed, and tail spasm was observed after the surgical procedure. After the operation, the rats were placed in their cages, and an antiseptic solution was used daily to prevent wound infection. The rats’ bladders were emptied manually twice a day.
Collection and evaluation of samples
Forty-eight hours after the surgical procedure, the rats were sacrificed under general anesthesia, blood was drawn from their heart directly, and then their spinal cords were excised due to total laminectomy (28)(Figure 2). The obtained tissue pieces were washed in SS and immediately stored at -80 ^°^C for biochemical studies. The tissue pieces were then placed in RNAlater solution (GeneMark, Maharashtra, India) and stored at -20 ^°^C for molecular studies. For histological examination, the samples were placed in 10% neutral formalin fixative and fixed for 48 hr**.**
Biochemical measurements
Total antioxidant status (TAS; mmol Trolox eq/l), total oxidant status (TOS; µmol H2O2 eq/l), and oxidative stress index (OSI; arbitrary unit) were detected using commercial kits (Rel Assay Diagnostics; Gaziantep, Turkey). The OSI levels in the sample were determined as the ratio of the TOS level to the TAS level. Lipid peroxidation was quantified at 532 nm by the measurement of malondialdehyde (MDA) reacted with the thiobarbituric acid according to the method of Ohkawa et al. The results were expressed in nmol/mg protein(29). The tissue protein amounts were measured by a BCA protein assay kit (iNtRON Biotechnology, Gyeonggi-do, Korea).
Molecular measurements
Total RNAs were isolated using the RNAzol RT solution (MRC, Cincinnati, OH, USA) according to the manufacturer’s instructions for quantifying mRNA expression in spinal cords. After completion of RNA isolation, the RNA concentration and purity were calculated with NanoDrop 2000 (Thermo Scientific, MA, USA). For this purpose, 1 µl RNA samples were pipetted in the device for determination of 260/280 and 260/230 ratios. Concentrations of all RNA samples were equalized before reverse transcription. RNAs were reverse transcribed into cDNA by using Script cDNA Synthesis Kit (Jena Bioscience, Germany). The resulting cDNA was amplified by qRT-PCR by using qPCR EvaGreenMaster (Solis BioDyne, Estonia). The real-time conditions were carried out on the CFX-96 RT-PCR System (Bio-Rad, CA, USA) as follows: 50 ^°^C for 2 min, 95 ^°^C for 2 min, and then 35 cycles of 95 ^°^C for 15 sec; 55 ^°^C for 20 sec, and 72 ^°^C for 20 sec. The relative mRNA transcript levels were calculated according to the 2–∆∆Ct method and the relative expression of each gene was normalized to that of glyceraldehyde-3-phosphate dehydrogenase (GAPDH). Primers were obtained from LGC Biosearch Technologies (Novato, CA, USA)(Table 1). All measurements were obtained in triplicates, and the specificity of amplicons was verified by melting curve analysis. The specific primers used are shown in Table 1.
Histological examination
The fixed specimens were subjected to routine histological follow-up methods and embedded in paraffin blocks. Tissue sections of 5 mm thickness were taken from the paraffin blocks with a microtome. The sections were stained with hematoxylin and eosin for microscopic examination. The samples obtained from all groups were evaluated at light microscopic level with an Olympus BH-2 (Olympus Corp., Tokyo, Japan) microscope, and all preparations were photographed with an Olympus DP-70 (Olympus Corp., Tokyo, Japan) digital camera. All specimens were evaluated and scored for cellular damage/ischemia, myelin damage/vacuolization, ependymal cell damage, and hemorrhage (30, 31).
Statistical analysis
SPSS software, version 22.0 for Windows (SPSS, Inc., Chicago, Illinois), was used for the statistical analysis of biochemical and histological data. One-way ANOVA was applied to the data that followed normal distribution, while the Kruskal-Wallis test was applied to the data that did not follow normal distribution. The results were presented as mean±standard deviation and median (minimum-maximum); P<0.05 was considered statistically significant.
Results
Biochemical, molecular, and histological findings
When the MDA levels were compared between the groups, as shown in Table 2, the MDA level increased after spinal injury. It decreased significantly in the group treated with BA before neural injury (BA+SCI)(P<0.01). The TOS level increased significantly after SCI compared to the control group (P<0.01). The TOS level in the BA+SCI group, the increase caused by SCI, was significantly reduced (P<0.05). When the TAS levels were compared between the groups, although antioxidant levels increased with both BA treatments, no significant difference was observed (P>0.05). The OSI level of the SCI group was significantly increased compared to the control group (P<0.001). In the BA+SCI group, the increase in OSI caused by SCI was significantly reduced (P<0.01)(Table 2).
Molecular apoptotic markers caspase-3 and cytochrome c levels were significantly increased after SCI compared to the control group (P<0.01 and P<0.001, respectively). The cytochrome c levels were significantly decreased in the BA-treated groups compared to the SCI group. This decrease was P<0.05 in the BA+SCI group and P<0.01 in the SCI+BA group (Table 2).
The histopathological examination of the control group was normal (Figure 3, Table 2). In the examination of the SCI group, ischemia in the motor neurons in the anterior horn in the grey matter of the medulla spinal cord and damage in ependymal cells were observed. Intense degenerative changes and hemorrhagic areas were observed in myelin in the white matter (Figure 3, Table 2). The histopathologic examination of the BA+SCI group revealed reduced damage. Although several ischaemic cells were observed in the anterior horn in the grey matter, near-normal multipolar motor neurons with euchromatic nuclei and cell bodies were observed in general examination (Figure 3, Table 2). Although reduced damage was noted in the SCI+BA group, hemorrhagic areas in the white matter, degeneration of myelin, and partial damage to ependymal cells were observed. Although a few ischaemic cells were observed in the anterior horn, overall examination revealed near-normal multipolar motor neurons with euchromatic nuclei and cell bodies (Figure 3, Table 2).
Discussion
Despite all technological and scientific advances, no definitive medical or surgical technique has yet been developed to ensure complete neural improvement in SCI. To reduce the effects of the damage, ensuring adequate spinal cord perfusion in intensive care, using neuroprotective agents such as methylprednisolone sodium succinate, decompressive surgery, and many neuroprotective and neuroregenerative therapy methods have been investigated by creating experimental SCI (23, 27, 32, 33).
With our study, the effects of BA on oxidative stress and apoptosis before and after SCI were investigated for the first time. BA is a trace element for living things. BA plays an important role in cell membrane functions and enzyme reactions, such as affecting the transduction of signals or regulatory ions across cell membranes (34). In the literature, the studies conducted with BA have increased recently, and its effects on nerve cells have also been investigated. In a study, it was reported that boron deprivation caused symptoms such as decreased electrical activity of the brain, loss of consciousness and psycho-motor activity, decreased movement and skills, and poor short-term memory (35). In the literature, there are studies showing that BA is protective against oxidative stress and has neuroprotective properties (13, 14, 36, 37). MDA levels, one of the oxidative stress markers, show an increase during the secondary injury (32, 38). In our study, the MDA level, which is an indicator of the peroxidation of polyunsaturated fatty acids in cell membranes, increased significantly. In addition, the TOS and OSI levels, which are other oxidative stress indicators, increased with SCI. Previous studies investigating the protective effect of minocycline and genistein in rats with SCI also showed an increase in TOS levels after SCI (38, 39). TAS levels have previously been shown to decrease after SCI (40). In our study, the TAS levels decreased moderately after SCI.
The exact mechanism of antioxidant and neuroprotective effects of BA has not yet been established. The effect of BA in the body varies depending on the prescribed dose. In a study, it was shown that low doses of BA showed neuroprotective properties in neurotoxicity induced by aluminum chloride in mouse brains, while high doses did not prevent neurotoxicity (41). In another experimental study, it was shown that 100 mg/kg BA had a positive effect on the recovery of axon and myelin damage in sciatic nerve injury (37). Since we have seen the protective effect of 100/kg BA dose in our other studies, we have previously conducted on brain cells (13, 14). We applied the same concentration of BA in this study. Biochemical markers (MDA, TOS, and OSI levels), which increased as a result of SCI injury, were significantly decreased with BA administration before trauma in this study. A significant decrease in the MDA value was achieved with BA administered after previously induced traumatic brain injury (13). In another study, it was shown that in vitro ethanol decreased the oxidative stress level on rat brain synaptosomes under in vitro conditions by applying BA (14). Again, the increased MDA level in rat brain synaptosomes by sodium fluoride induction was reduced by applying BA (36). BA has been previously reported to exert its antioxidant effect by inhibiting lipid peroxidase and maintaining the integrity of the cell membrane (1, 36, 42). However, there are studies in which nonsignificant antioxidant increases were observed with BA administration, as well as studies in which no increase was observed (14, 43). In our study, the TAS levels decreased after SCI, but administration of BA before and after injury caused moderate increases. In our study, a more effective decrease in the MDA, TOS, and OSI levels was found with BA administered before SCI. This situation emphasized the protective effect of BA on cell membrane integrity rather than its therapeutic properties.
After traumatic SCI, apoptotic cell death markers, such as cytochrome c, caspase-3, caspase-9, and Apaf-1 (apoptotic peptidase activating factor 1), which are markers of apoptosis, increased with the decrease in the activity of mitochondrial respiratory enzymes (44-46). In our study, parallel with previous studies, cytochrome c and caspase-3 levels were found to increase after SCI. In our study, it was observed that BA decreased the cytochrome c level, which increased after cord injury, and decreased caspase-3 activities moderately in the groups administered before and after SCI. It was previously found that oxidative stress in liver cells increased as a result of chronic alcohol use. As a result, the caspase-3 activity and DNA fragmentation, which are among the markers of apoptosis, increased. BA administration with alcohol caused a decrease in oxidative stress and thus decreased caspase-3 and DNA fragmentation, which are markers of apoptosis (18). In our study, increased oxidative stress after SCI caused apoptosis to be triggered, and we suggest that BA applications decreased the amount of Ca+ and, indirectly, cytochrome c levels during cell death by protecting cell membrane integrity.
In addition, histological features of the spinal cortex of rats with acute traumatic injury in previous studies showed that hematoma, edema, and cell damage increased after SCI (47, 48). In addition, it was found that NeuN and Nissl staining levels, which are markers of mature neurons, decreased after SCI, and the number of TUNEL-positive cells, which is an indicator of apoptosis, increased after SCI (49). In our study, in parallel with the previous studies, it was found that cell, myelin, and ependymal cell damage, as well as hemorrhage levels, increased after SCI. In a study in which BA was applied after spinal cortex ischemia, edema, hemorrhage, and neuronal degeneration were found to be decreased (48). In a study conducted by our group, increased edema and necrotic neuron numbers after traumatic brain injury decreased after BA application (13). In the histological examination of our study, increased cell, myelin, ependymal damage, and hemorrhage areas after SCI showed significant changes in the groups in which BA was applied before and after SCI. This healing was found to be more effective in the group administered BA before trauma. Histologic results were consistent with our biochemical results.
Experimental groups, laminectomy (LAM), spinal cord injury (SCI), 100 mg/kg boric acid (BA)
Spinal cords of control and spinal cord injury (SCI) groups - The damaged area is at the level of the scissors
Table 1: Primers used in real-time polymerase chain reaction analysis for Caspase-3 and Cytochrome-c
<table><thead><tr><th align="left" rowspan="1" colspan="1">Primer Name</th><th align="center" rowspan="1" colspan="1">Forward Primer Sequence (5′-3′)</th><th align="center" rowspan="1" colspan="1">Reverse Primer Sequence (3′-5′)</th></tr></thead><tbody><tr><td align="left" rowspan="1" colspan="1">Caspase-3 (<ext-link xmlns:xlink="http://www.w3.org/1999/xlink">NM_009810.3</ext-link>)</td><td align="center" rowspan="1" colspan="1">TGACTGGAAAGCCGAAACTC</td><td align="center" rowspan="1" colspan="1">GTAGAGTAAGCATACAGGAAGTCAG</td></tr><tr><td align="left" rowspan="1" colspan="1">Cytochrome-c (<ext-link xmlns:xlink="http://www.w3.org/1999/xlink">NM_007808.5</ext-link>)</td><td align="center" rowspan="1" colspan="1">AAGGGAGGCAAGCATAAGAC</td><td align="center" rowspan="1" colspan="1">ATTCTCCAAATACTCCATCAGGG</td></tr><tr><td align="left" rowspan="1" colspan="1">GAPDH<break/>(Housekeeping) (<ext-link xmlns:xlink="http://www.w3.org/1999/xlink">NM_008084.3</ext-link>)</td><td align="center" rowspan="1" colspan="1">CCTCGTCCCGTAGACAAAATG</td><td align="center" rowspan="1" colspan="1">TGTAGTTGAGGTCAATGAAGGG</td></tr></tbody></table>Table 2: Comparison of all groups with biochemical, molecular and histological findings
<table><thead><tr><th align="left" rowspan="1" colspan="1">Groups → Biochemical Parameters ↓</th><th align="center" rowspan="1" colspan="1">Control</th><th align="center" rowspan="1" colspan="1">SCI</th><th align="center" rowspan="1" colspan="1">BA+SCI</th><th align="center" rowspan="1" colspan="1">SCI+BA</th><th align="center" rowspan="1" colspan="1">p</th></tr></thead><tbody><tr><td align="left" rowspan="1" colspan="1">MDA<break/>(nmol/mg protein)</td><td align="center" rowspan="1" colspan="1">13.94±3.23</td><td align="center" rowspan="1" colspan="1">17.69±2.6</td><td align="center" rowspan="1" colspan="1">11.25±3.55</td><td align="center" rowspan="1" colspan="1">15.40±3.02</td><td align="center" rowspan="1" colspan="1">SCI & BA+SCI **</td></tr><tr><td align="left" rowspan="1" colspan="1">TOS<break/>(µmol H<sub>2</sub>O<sub>2</sub> eq/l</td><td align="center" rowspan="1" colspan="1">41.50±8.4</td><td align="center" rowspan="1" colspan="1">62±11.16</td><td align="center" rowspan="1" colspan="1">48.75±10.69</td><td align="center" rowspan="1" colspan="1">54.5±7.3</td><td align="center" rowspan="1" colspan="1">Control & SCI **<break/>SCI & BA+SCI *</td></tr><tr><td align="left" rowspan="1" colspan="1">TAS<break/>(mmol Trolox eq/l)</td><td align="center" rowspan="1" colspan="1">2.86±1.17</td><td align="center" rowspan="1" colspan="1">2.06±0.49</td><td align="center" rowspan="1" colspan="1">3.23±1.4</td><td align="center" rowspan="1" colspan="1">2.34±0.28</td><td align="center" rowspan="1" colspan="1">No significant</td></tr><tr><td align="left" rowspan="1" colspan="1">OSI<break/>(OSI; arbitrary unit)</td><td align="center" rowspan="1" colspan="1">16.17±7.62</td><td align="center" rowspan="1" colspan="1">31.25±8.21</td><td align="center" rowspan="1" colspan="1">17.36±6.76</td><td align="center" rowspan="1" colspan="1">23.59±4.39</td><td align="center" rowspan="1" colspan="1">Control & SCI ***<break/>SCI & BA+SCI **</td></tr><tr><td align="left" rowspan="1" colspan="1">Groups → Molecular Parameters ↓</td><td align="center" rowspan="1" colspan="1">Control</td><td align="center" rowspan="1" colspan="1">SCI</td><td align="center" rowspan="1" colspan="1">BA+SCI</td><td align="center" rowspan="1" colspan="1">SCI+BA</td><td align="center" rowspan="1" colspan="1">p</td></tr><tr><td align="left" rowspan="1" colspan="1">Cytochrome c</td><td align="center" rowspan="1" colspan="1">1.0</td><td align="center" rowspan="1" colspan="1">2.183±0.58</td><td align="center" rowspan="1" colspan="1">1.461±0.36</td><td align="center" rowspan="1" colspan="1">1.233±0.61</td><td align="center" rowspan="1" colspan="1">Control & SCI ***<break/>SCI & BA+SCI *<break/>SCI & SCI+BA **</td></tr><tr><td align="left" rowspan="1" colspan="1">Caspase 3</td><td align="center" rowspan="1" colspan="1">1.0<break/>(1-1)</td><td align="center" rowspan="1" colspan="1">2.02<break/>(1.77-2.17)</td><td align="center" rowspan="1" colspan="1">1.8<break/>(1.23-2.25)</td><td align="center" rowspan="1" colspan="1">1.42<break/>(1.06-1.71)</td><td align="center" rowspan="1" colspan="1">Control & SCI **</td></tr><tr><td align="left" rowspan="1" colspan="1">Groups→ Histological Parameters ↓</td><td align="center" rowspan="1" colspan="1">Control</td><td align="center" rowspan="1" colspan="1">SCI</td><td align="center" rowspan="1" colspan="1">BA+SCI</td><td align="center" rowspan="1" colspan="1">SCI+BA</td><td align="center" rowspan="1" colspan="1">p</td></tr><tr><td align="left" rowspan="1" colspan="1">Cell damage/Ischemia</td><td align="center" rowspan="1" colspan="1">0 (0-0)</td><td align="center" rowspan="1" colspan="1">3 (2-3)</td><td align="center" rowspan="1" colspan="1">1 (1-2)</td><td align="center" rowspan="1" colspan="1">2 (1-2)</td><td align="center" rowspan="1" colspan="1">Control & SCI ****<break/>Control & SCI+BA *<break/>BA+SCI & SCI *</td></tr><tr><td align="left" rowspan="1" colspan="1">Myelin damage/Vacuolization</td><td align="center" rowspan="1" colspan="1">0 (0-0)</td><td align="center" rowspan="1" colspan="1">3 (2-3)</td><td align="center" rowspan="1" colspan="1">0 (0-1)</td><td align="center" rowspan="1" colspan="1">2 (1-2)</td><td align="center" rowspan="1" colspan="1">Control & SCI ****<break/>Control & SCI+BA *<break/>BA+SCI & SCI ***</td></tr><tr><td align="left" rowspan="1" colspan="1">Ependymal cell damaged</td><td align="center" rowspan="1" colspan="1">0 (0-0)</td><td align="center" rowspan="1" colspan="1">2 (2-3)</td><td align="center" rowspan="1" colspan="1">0 (0-1)</td><td align="center" rowspan="1" colspan="1">1 (1-2)</td><td align="center" rowspan="1" colspan="1">Control & SCI ****<break/>Control & SCI+BA *<break/>BA+SCI & SCI ***</td></tr><tr><td align="left" rowspan="1" colspan="1">Hemorrhage</td><td align="center" rowspan="1" colspan="1">0 (0-0)</td><td align="center" rowspan="1" colspan="1">3 (2-3)</td><td align="center" rowspan="1" colspan="1">0 (0-1)</td><td align="center" rowspan="1" colspan="1">0 (0-1)</td><td align="center" rowspan="1" colspan="1">Control & SCI ****<break/>BA+SCI & SCI ***<break/>SCI+BA & SCI *</td></tr></tbody></table>Histological H&E staining of spinal cord sections from all groups
Conclusion
Our data demonstrated that neuronal injury is elevated after SCI via oxidative stress. Moreover, neuronal apoptosis might be triggered by mitochondrial cytochrome c release and depends on elevated caspase-3 activities. We also showed that boric acid might be effective as an antioxidative agent in SCI, especially in boric acid pre-treatment. However, the antiapoptotic role of boric acid is limited.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Smith E Fitzpatrick P Murtagh J Lyons F Morris S Synnott K Epidemiology of traumatic spinal cord injury in Ireland, 2010-2015 Neuroepidemiology 20185119242976391410.1159/000488146 · doi ↗ · pubmed ↗
- 2Halvorsen A Pettersen A Nilsen S Halle KK Schaanning EE Rekand T Epidemiology of traumatic spinal cord injury in Norway in 2012–2016: a registry-based cross-sectional study Spinal Cord 2019573313383057377010.1038/s 41393-018-0225-5 · doi ↗ · pubmed ↗
- 3Lu Y Shang Z Zhang W Pang M Hu X Dai Y Global incidence and characteristics of spinal cord injury since 2000–2021: a systematic review and meta-analysis BMC Med 2024222852983897297110.1186/s 12916-024-03514-9PMC 11229207 · doi ↗ · pubmed ↗
- 4Marcon RM Barros Filho TE Pd Oliveira RP Cristante AF Taricco MA Colares G Experimental study on the action of methylprednisolone on Wistar rats before spinal cord injury Acta Ortopédica Brasileira 2010182630
- 5Silva NA Sousa N Reis RL Salgado AJ From basics to clinical: A comprehensive review on spinal cord injury Prog Neurobiol 201411425572426980410.1016/j.pneurobio.2013.11.002 · doi ↗ · pubmed ↗
- 6Fatima G Sharma V Das S Mahdi A Oxidative stress and antioxidative parameters in patients with spinal cord injury: Implications in the pathogenesis of disease Spinal cord 201553362536652810.1038/sc.2014.178 · doi ↗ · pubmed ↗
- 7Ma Y Li P Ju C Zuo X Li X Ding T Photobiomodulation attenuates neurotoxic polarization of macrophages by inhibiting the notch 1-HIF-1α/NF-κB signaling pathway in mice with spinal cord injury Front Immunol 2022138169528169683537106510.3389/fimmu.2022.816952 PMC 8968029 · doi ↗ · pubmed ↗
- 8Chen M Chen S Lin D Carvedilol protects bone marrow stem cells against hydrogen peroxide-induced cell death via PI 3K-AKT pathway Biomed Pharmacother 2016782572632689845010.1016/j.biopha.2016.01.008 · doi ↗ · pubmed ↗