Optimal duration of ex vivo lung perfusion for heat stress-mediated therapeutic reconditioning of damaged rat donor lungs
Roumen Parapanov, Anne Debonneville, Manon Allouche, Jérôme Lugrin, Tanguy Lugon-Moulin, Etienne Abdelnour-Berchtold, Lucas Liaudet, Thorsten Krueger

TL;DR
Applying heat stress during lung perfusion can protect damaged rat donor lungs, with optimal benefits seen after 4.5 hours of recovery.
Contribution
The study identifies the optimal duration of ex vivo lung perfusion following heat stress to maximize therapeutic benefits in damaged donor lungs.
Findings
Heat stress during EVLP improves lung function and reduces damage markers in rat lungs.
EVLP for 4.5 hours after heat stress yields the best physiological and molecular outcomes.
Prolonged EVLP beyond 4.5 hours leads to deterioration in lung function.
Abstract
Transient heat stress (HS) application during experimental ex vivo lung perfusion (EVLP) of warm ischaemic (WI) rat lungs produces a range of therapeutic benefits. Here, we explored whether different EVLP durations after HS application would influence its therapeutic effects. In protocol 1, WI rat lungs were exposed to HS (41.5°C, 60–90 min EVLP), and EVLP was maintained for 3, 4.5 or 6 h (n = 5/group), followed by physiological measurements (compliance, oedema, oxygenation capacity). In protocol 2, WI rat lungs treated with (HS groups) or without HS (control groups) were maintained for 3 or 4.5 h EVLP (n = 5/group), followed by physiological evaluation and measurements (lung tissue) of heat shock proteins (HSP70, HSP27, HS90, GRP78), endogenous proteins (surfactant protein-D, CC16, platelet endothelial cell adhesion molecule-1), anti-apoptotic (Bcl2, Bcl-xL) and pro-apoptotic proteins…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
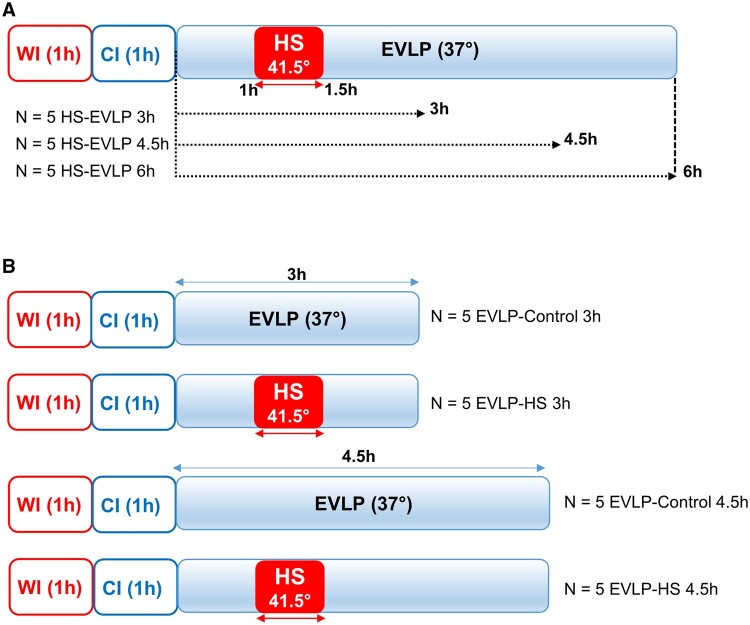 Figure 1
Figure 1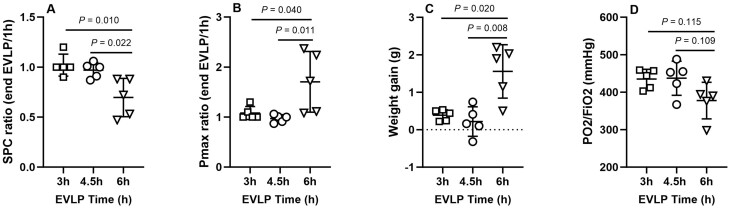 Figure 2
Figure 2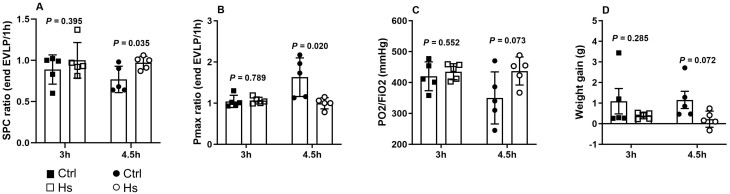 Figure 3
Figure 3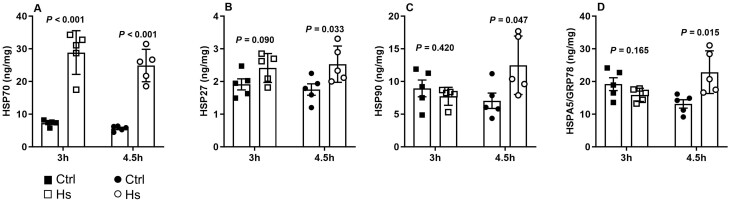 Figure 4
Figure 4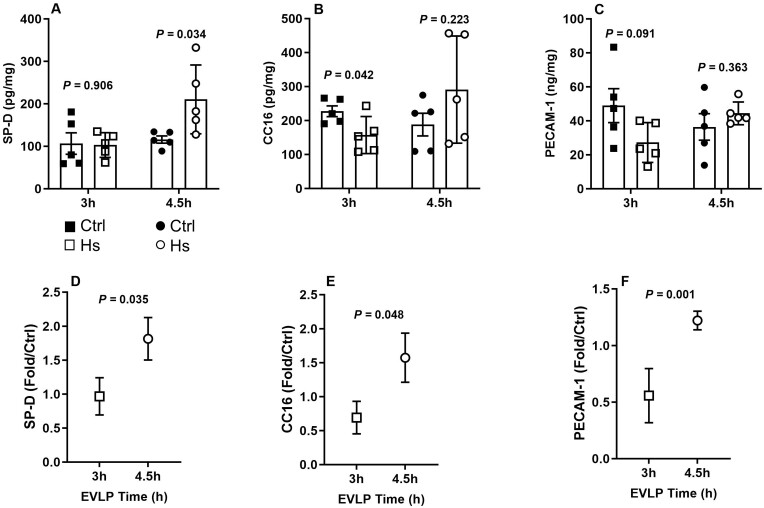 Figure 5
Figure 5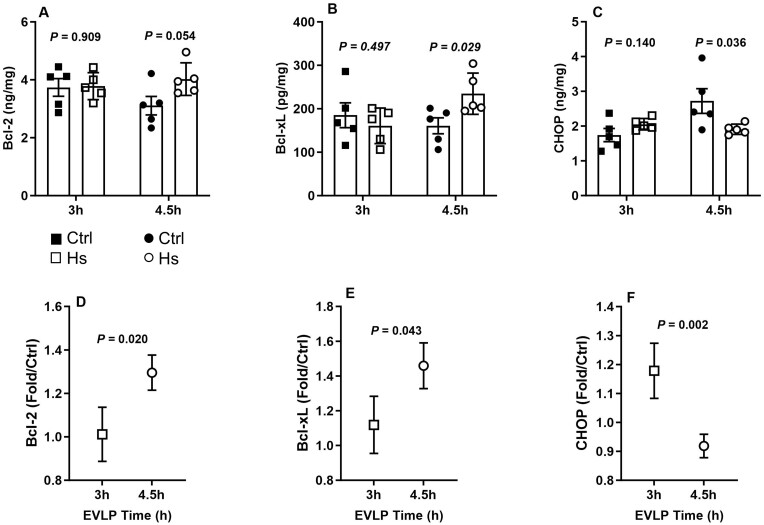 Figure 6
Figure 6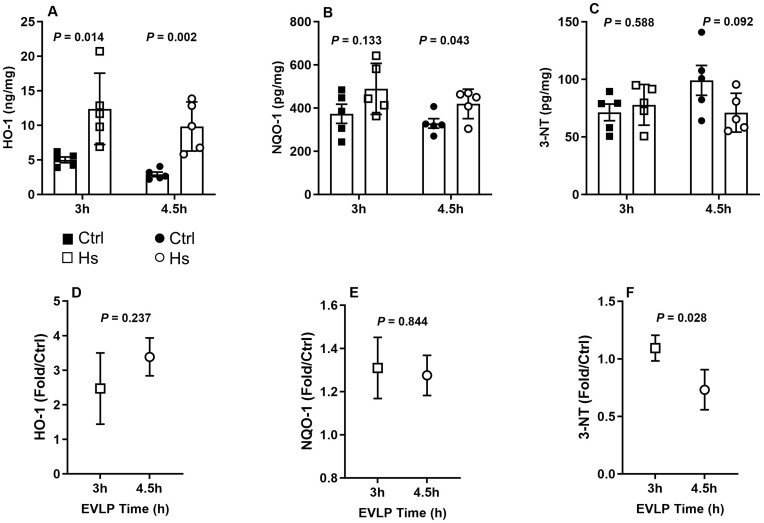 Figure 7
Figure 7- —Swiss National Science Foundation10.13039/501100001711
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsOrgan Transplantation Techniques and Outcomes · Transplantation: Methods and Outcomes · Cardiac Ischemia and Reperfusion
INTRODUCTION
Lung transplantation (LTx) is the unique treatment for end-stage lung diseases, but it is limited by the shortage of transplantable lungs. To cope with this problem, many lung transplant centres implemented ex vivo lung perfusion (EVLP) for the physiological evaluation of marginal donor lungs [1, 2]. EVLP may also allow the administration of therapies to improve lung function and reduce the risk of primary graft dysfunction after LTx, as demonstrated in experimental studies [3, 4]. Beyond pharmacological treatments, EVLP could also serve to stimulate endogenous protective responses following the ex vivo application of a physiological stress. One such stimulus can be realized by exposing lungs to a transient non-lethal mild heat stress (HS), which confers resistance to future stressful stimuli by triggering a heat shock response through the induction of heat shock proteins (HSPs) acting as molecular chaperones [5]. In this respect, we recently demonstrated that an ex vivo mild HS application to damaged rat lungs reduced cellular injury and physiological deterioration after EVLP and LTx [6, 7].
The molecular signals triggered by HS evolve according to certain chronological steps. During initiation of the HS response, there is a global repression of protein synthesis accompanied by the selective expression of HSPs [8]. The return to normothermic conditions after HS allows the restoration of cell homeostasis and the de novo expression of genes necessary for stress adaptation [5]. The kinetic expression of heat-responsive genes indicates that post-HS recovery time is essential for the resistance conferred by this strategy. Accordingly, understanding such kinetics appears crucial to optimize the therapeutic benefits of HS preconditioning [9]. Therefore, this study aimed to investigate the influence of various recovery times after HS application during EVLP on the activation of endogenous protective mechanisms to determine the best time window for HS preconditioning during EVLP.
MATERIAL AND METHODS
Animals
All experiments were approved by our local animal committee (Direction générale de l’agriculture, la viticulture et affaires vétérinaires de l’Etat de Vaud, authorizations Nr 3456, 21 April 2019, and Nr 3456x1, 29 October 2022). Twenty-five male Sprague-Dawley rats (350–450 g, Charles River, Saint-Germain-Nuelles, France) were used in this study and treated in accordance with ‘Guidelines for the Care and Use of Laboratory Animals’ (NIH Publication no. 96-23).
Experimental model
We used our published model of warm ischaemic rat lungs and EVLP, as previously described [6, 7]. Briefly, anaesthetized, tracheotomized and mechanically ventilated rats were sacrificed after systemic anticoagulation (intravenous heparin 600 IU) and kept in situ for 1 h at room temperature (warm ischaemic time). The pulmonary artery and left ventricle were then cannulated via a median sternotomy, lungs were flushed (PERFADEX^®^ Plus, 4°C, 25 ml) and inflated (7 ml/kg, FiO_2_ 0.5). The heart-lung block was removed, stored at 4°C in PERFADEX^®^ Plus for 1 h (cold ischaemia), weighted and mounted on the EVLP system.
Ex vivo lung perfusion and heat stress preconditioning
EVLP and HS were performed as previously described [6]. Briefly, the perfusate (Steen^®^ solution) was progressively warmed from 16° to 37° over 1 h, at increasing flow (from 2 to 7.5% estimated cardiac output). Ventilation (Flexivent FX3 ventilator, SCIREQ Inc., Montréal, Canada) was started at 35°C perfusate temperature (tidal volume 3 ml/kg, rate 7/min, room air), then increased to 6 ml/kg and 30/min at 37°C. HS was applied after 1 h EVLP by rapidly warming the perfusate at 41.5°C for 30 min, followed by return to 37°C until the end of EVLP. The temperature of the perfusate was monitored at the entry of the pulmonary artery with a TES-1303 Type-K digital thermometer (TES Electrical Electronic Corp., Taipei, Taiwan).
Experimental protocols
The experimental design is depicted in Fig. 1. In protocol 1, 3 groups of lungs (n = 5/group) were exposed to HS for 30 min after 1 h EVLP as described above (60–90 min EVLP) and then maintained under EVLP for 3 h (HS_3h), 4.5 h (HS4.5h) or 6 h (HS6h) to allow, respectively, 90, 180 and 270 min recovery time after HS. In protocol 2, we compared lungs exposed to HS and perfused 3 h (HS_3h, n = 5) or 4.5 h (HS_4.5h_, n = 5) to parallel control groups that did not undergo HS (Ctrl_3h, n = 5 and Ctrl4.5h_, n = 5). At the end of EVLP, the heart-lung block was weighed and the difference from weight before EVLP was used as an index of lung oedema, then the lungs were kept frozen at −80°C for further analyses. In both protocols, groups were alternate each day but were not randomized. The sample size was estimated from our previous study on HS-preconditioning [7], in which we measured several biomarkers in the perfusate at selected time-points during EVLP. At the 4.5 h time-point, biomarkers differed by a mean proportion of 1.84 between HS and control groups, with a mean standard deviation of 47%. Assuming comparable proportions and variance in the present study, we calculated (α: 5%, power: 80%, two-sided test) that a size of n = 5/group (HS and controls) would be appropriate for meaningful analyses.
Experimental design. (A) Protocol 1: lungs exposed to 1 h warm ischaemia (WI) and 1 h cold ischaemia (CI) were perfused in an ex vivo lung perfusion (EVLP) system for 3, 4.5 or 6 h at 37°C, with the application of a transient heat stress (HS) at 41.5° from 1 to 1.5 h EVLP. (B) Protocol 2: lungs exposed to 1 h WI and 1 h CI were perfused in an EVLP system for 3 or 4.5 h, with (EVLP-HS) or without (EVLP-control) the application of a transient HS at 41.5° from 1 to 1.5 h EVLP.
Measurements
Physiological measurements
Static pulmonary compliance and peak airway pressure (Pmax) were determined as described previously [6, 7], expressed as the ratio of the value at the end of EVLP to that at 1 h EVLP (before HS application) [6, 7]. The partial pressure of O_2_ was measured at the end of EVLP in the effluent perfusate (CG4+ cartridge, Abbott i-STAT analyzer, East Windsor, KJ).
Biological measurements
Heat shock protein expression
Lung tissue was homogenized in RIPA buffer with proteases/phosphatase inhibitors and sonicated. The concentrations of HSPs (HSP70, HSP27, HSP90 and HSPA5/GRP78) were determined using enzyme-linked immunosorbent assays (Supplementary Material, Table S1), expressed in ng/mg tissue protein, measured by the BCA assay (Thermo Scientific Pierce, Rockford, IL, USA).
Expression of lung epithelial and endothelial proteins, pro- and anti-apoptotic proteins, antioxidant proteins and 3-nitrotyrosine
Lung homogenates were assayed using specific enzyme-linked immunosorbent assays (Supplementary Material, Table S1) for the epithelial proteins Surfactant protein-D (SP-D) and Clara cell protein (CC16), the endothelial protein platelet endothelial cell adhesion molecule-1 (PECAM-1), the anti-apoptotic proteins B-cell CLL/lymphoma-2 (Bcl2) and Bcl2-like protein-1 (Bcl-xL), the pro-apoptotic proteins Bcl2-associated X protein (Bax), CCAAT/enhancer binding-protein (C/EBP) homologous protein (CHOP), as well as the antioxidant proteins NAD(P)H quinone-1-dehydrogenase (NQO-1) and heme oxygenase-1 (HO-1). The levels of lung 3-nitrotyrosine (3-NT) were determined as an index of nitroxidative stress. All data were expressed in ng/mg or pg/mg tissue protein.
Statistical analysis
Results are presented as means±SD. In protocol 1, measurements between the 3 groups were compared using one-way ANOVA followed by Tukey test. In protocol 2, comparisons between HS groups and their respective controls at 3 h and 4.5 h were done by unpaired two-tailed *t-*test after log transformation of the data in case of non-normal distribution (assessed by the Shapiro–Wilk test). Furthermore, we compared relative protein expressions (fold changes) between the HS groups and their respective controls at 3 h and 4.5 h EVLP, using unpaired two-tailed *t-*test. Statistical significance was assigned to a P < 0.05. GraphPad Prism, version 10.1.2 (GraphPad Software Inc., La Jolla, CA) was used for all statistical analyses.
RESULTS
First experimental protocol: comparison of functional outcome of lungs exposed to heat stress and different recovery times
As indicated in Fig. 2, lungs from the group HS_6h_ displayed significant reduction of static pulmonary compliance (Fig. 2A) and increase in Pmax (Fig. 2B), associated with greater oedema development (weight gain during EVLP, Fig. 2C) and a non-significant trend for reduced oxygenation capacity (P/FO_2_ in the EVLP effluent, Fig. 2D). Owing to this significant deterioration of lungs from the HS_6h_ group, further investigations were performed only in the groups HS_3h_ and HS_4.5h_ and their respective controls_._
Pulmonary physiological variables following heat stress (HS) and ex vivo lung perfusion (EVLP) durations of 3 h, 4.5 h and 6 h. (A) Static pulmonary compliance (SPC), expressed as the ratio of SPC at the end of EVLP/1 h EVLP. (B) Maximal airway pressure (Pmax) expressed as the ratio of Pmax at the end of EVLP/1h. (C) Lung oedema development, assessed by the weight gain of the lungs at the end of EVLP. (D) Lung oxygenation capacity expressed in P/F ratio. N = 5 groups
Second experimental protocol: comparison of lungs exposed or not to heat stress after 3 h and 4.5 h ex vivo lung perfusion
Physiological variables
After 3 h EVLP, HS-treated lungs (HS_3h) showed similar respiratory parameters compared to Ctrl3h_ lungs (Fig. 3A–D). In contrast, after 4.5 h EVLP, lungs from the HS_4.5h_ group demonstrated significantly better functional outcomes than lungs from the Ctrl_4.5h_ group, with significantly higher static pulmonary compliance, lower Pmax and a trend (P = 0.07) for better oxygenation and reduced oedema (Fig. 3A–D).
Lung physiological variables after 3 and 4.5 h ex vivo lung perfusion (EVLP). (A) Static pulmonary compliance (SPC), expressed as SPC ratio at the end of EVLP/1 h EVLP. (B) Maximal airway pressure (Pmax) expressed as the ratio of Pmax at the end of EVLP/1 h. (C) Lung oxygenation capacity expressed in P/F ratio. (D) Lung oedema development, assessed by the weight gain of the lungs at the end of EVLP. N = 5 groups.
Biological variables
Heat shock proteins
As shown in Fig. 4A, a significant induction of HSP70 occurred in HS_3h_ and HS_4.5h_ compared to their respective controls. In contrast, levels of HSP27 (Fig. 4B), HSP 90 (Fig. 4C) and HSPA5/GRP78 (Fig. 4D) did not differ between HS_3h_ and Ctrl_3h, whereas they were significantly increased in HS4.5h_ compared to Ctrl_4.5h_. When expressed as fold change in comparison to controls (not shown), the change tended to be greater for HSP70 and HSP27 and was significantly greater for HSP90 and HSPA5/GRP78 at 4.5 h than at 3 h.
Lung expression of heat shock proteins at the end of 3 or 4.5 h ex vivo lung perfusion (EVLP). Lung tissue concentrations of (A) HSP70, (B) HSP27, (C) HSP90 and (D) HSPA5/GRP78. N = 5 groups.
Lung epithelial and endothelial proteins
Lungs in both Ctrl_3h_ and HS_3h_ groups displayed similar expression of SP-D (Fig. 5A), whereas expression of CC16 and PECAM-1 decreased in HS_3h_ compared to Ctrl_3h_ (Fig. 5B and C). At 4.5 h EVLP, a significant increase of SP-D expression occurred in HS_4.5h_ group (Fig. 5A), while CC16 and PECAM-1 did not significantly vary with respect to Ctrl_4.5h_ (Fig. 5B and C). The variation of protein expression in the HS groups compared to their respective controls (fold changes) indicated that the changes were significantly greater for all examined proteins after 4.5 h than after 3 h EVLP (Fig. 5D–F).
Expression of lung endogenous proteins at the end of 3 h or 4.5 h ex vivo lung perfusion (EVLP). (A) Surfactant protein D (SP-D), (B) Clara cell protein (CC16), (C) Platelet-endothelial cell adhesion molecule-1 (PECAM-1), all expressed in ng/mg or pg/mg lung tissue protein. (D) SP-D, (E) CC16, (F) PECAM-1, expressed as fold change in heat stress (HS) versus controls at each time-point. N = 5 groups.
Anti- and pro-apoptotic proteins
After 3 h EVLP, the levels of anti-apoptotic Bcl-2 and Bcl-xL (Fig. 6A and B) and pro-apoptotic CHOP (Fig. 6C) and Bax (not shown) proteins were not significantly influenced by HS. In contrast, after 4.5 h EVLP, lungs from the HS_4.5h_ group exhibited significantly higher levels of Bcl-2 and Bcl-xL, lower levels of CHOP (Fig. 6A–C) and a trend for lower Bax levels (not shown) compared to the Ctrl_4.5h_ group. This was further illustrated by the relative variations of the different proteins, showing that the influence of HS was significantly more pronounced after 4.5 h than 3 h EVLP, with a positive impact on anti-apoptotic Bcl-2 and Bcl-xL (Fig. 6D and E), and a negative impact on pro-apoptotic CHOP (Fig. 6F).
Pro- and anti-apoptotic proteins in lung tissue at 3 and 4.5 h ex vivo lung perfusion (EVLP). (A) B-cell leukemia/lymphoma-2-protein (Bcl-2), (B) B-cell lymphoma-extra large (Bcl-xL), (C) CCAAT/enhancer binding-protein (C/EBP) homologous protein (CHOP), expressed in ng/mg or pg/mg lung protein. Relative expression (fold changes in heat stress (HS) versus control lungs at 3 and 4.5 h EVLP) of (D) Bcl-2, (E) Bcl-xL, (F) CHOP. N = 5 groups.
Antioxidant proteins and 3-nitrotyrosine
While both HS_3h_ and HS_4.5h_ groups displayed a significantly increased expression of HO-1 in comparison to their respective controls (Fig. 7A), a significant increase of NQO-1 only occurred in HS_4.5h_ group (Fig. 7B). Levels of 3-NT were comparable between lungs from HS_3h_ and Ctrl_3h_ groups, whereas they tended to be lower in HS_4.5h_ compared to Ctrl_4.5h_ (Fig. 7C). Fold change analyses revealed a comparable influence of HS on HO-1 and NQO-1 after 3 and 4.5 h EVLP (Fig. 7D and E). In contrast, HS had a significantly larger effect on 3-NT after 4.5 h than 3 h EVLP (Fig. 7F).
Antioxidant proteins and 3-nitrotyrosine in lungs at the end of ex vivo lung perfusion (EVLP) for 3 or 4.5 h. Lung tissue levels of (A) HO-1 (heme oxygenase-1), (B) NQO-1 [NAD(P)H quinone dehydrogenase 1], (C) 3-NT (3-nitrotyrosine). Relative expression [fold changes heat stress (HS) versus controls] of (D) HO-1, (E) NQO-1 and (F) 3-NT at 3 and 4.5 h EVLP. N = 5 groups.
DISCUSSION
We recently demonstrated that the ex vivo application of a transient mild HS to damaged donor lungs reduced lung injury and dysfunction during EVLP and LTx, via the expression of HSPs and reduced inflammation, oxidative stress and cell death [6, 7]. Owing to the dynamic nature of the HS response [9], we postulated that different post-HS recovery times might influence its therapeutic potential. In a first set of experiments, we observed that lungs subjected to HS and maintained under EVLP for 3 and 4.5 h exhibited stable physiology, contrasting with reduced compliance, increased airway pressure and oedema formation after 6 h. We found in a previous study [7] that HS-treated lungs were protected against endothelial damage and dysfunction after 6 h EVLP, despite a slight degree of oedema and reduced compliance, in agreement with our present findings. Therefore, even though lungs remain significantly protected by HS after 6 h EVLP, they may still develop moderate physiological deterioration, which indicates that the protective effects of HS cannot be fully maintained for extended (6 h) durations of EVLP. Accordingly, we decided to restrict our subsequent investigations to EVLP durations of 3 h and 4.5 h, during which HS-treated lungs were reperfused without any apparent functional impairment.
The hallmark of the HS response is the induction of various HSPs [5]. Previous studies indicated that the main stress-inducible HSP70 and HSP27 are rapidly expressed (30 min–2h) after HS and then remain elevated for several hours [7, 9]. In agreement with these findings, we found that the induction of these 2 HSPs was comparable after 3 and 4.5 h, implying that extending post-HS recovery time beyond 90 min (3 h EVLP) is not necessary to obtain their full expression. In contrast, 2 additional inducible HSPs (HSP90 and HSPA5/GRP78) were significantly expressed only after 4.5 h, indicating that an EVLP model allowing 180 min post-HS recovery (EVLP 4.5 h) is necessary to optimize the expression of major inducible HSPs in physiologically intact lungs. This is an important finding, given that the efficiency of the protective functions of HSPs directly correlates with their expression levels [5].
An initial temporary downregulation of general protein transcription and translation is a characteristic response to HS, enabling the reduction of cellular misfolded protein accumulation [6, 8]. Consistently, we found that the expression of key epithelial (SP-D, CC16) and endothelial (PECAM-1) proteins was differentially influenced according to the duration of post-HS recovery. SP-D, synthesized by alveolar type II cells, plays important roles in lung innate immune defences [10], whereas CC16, secreted by Clara Cells, has potent antioxidant/anti-inflammatory roles [11]. Notably, the interest of SP-D and CC16 as biomarkers of epithelial injury and primary graft dysfunction after LTx has been highlighted by previous investigators [12, 13]. PECAM-1 is an endothelial protein essential for endothelial barrier integrity [14]. When comparing the relative expression of SP-D, CC16 and PECAM-1 in HS and control lungs, we found that it was either unchanged or reduced after 3 h but was increased after 4.5 h, consistent with an early slowdown of transcription followed by recovery. We conclude, therefore, that an EVLP duration of 4.5 h, allowing sufficient post-HS recovery time, is required to secure the appropriate expression of key pulmonary homeostatic proteins.
Apoptosis represents an important mode of cell death in lung ischaemia–reperfusion and LTx, and lung grafts undergoing warm ischaemia display a pro-apoptotic transcriptomic signature [15] and suffer enhanced apoptosis during EVLP [16]. Intrinsic (mitochondrial) apoptosis is regulated by the balanced expression of anti (Bcl-2, BcL-xL) and pro-apoptotic (Bax) proteins [17], and several HSPs can shift this balance towards a predominantly anti-apoptotic phenotype [18, 19]. Clinically, the role of several genes of the Bcl-2 family as potential biomarkers and targets of treatment for primary graft dysfunction has been recently underscored [20]. We found that HS did not alter the expression of these proteins after 3 h EVLP, contrasting with a significant enhancement of Bcl2 and Bcl-XL expression after 4.5 h. This was associated with decreased expression of CHOP, a pro-apoptotic protein induced upon endoplasmic reticulum stress [21], an effect likely related to the strong induction of HSPA5/GRP78 at 4.5 h EVLP, a regulator of endoplasmic reticulum homeostasis preventing CHOP induction and downstream apoptosis [21]. Taken together, these data indicate that extending post-HS recovery time from 90 (EVLP 3 h) to 180 min (EVLP 4.5 h) triggered important pro-survival signals in reperfused lungs.
A consistent response to HS is a steady protection against oxidative stress, largely mediated by the activation of the NRF-2 transcription factor and downstream targets such as HO-1 and NQO-1 [22, 23]. Accordingly, the expression of these 2 antioxidant proteins was promoted in lungs exposed to HS to a comparable extent after 3 or 4.5 h EVLP. Therefore, such adaptation appears as an early response to HS with no apparent time-dependent amplification. Nevertheless, we found that the levels of 3-NT, a footprint of the strong oxidant peroxynitrite [24], were attenuated after 4.5 h, but not 3 h EVLP. This suggests that a longer post-HS recovery time may favour more extensive protection against oxidative stress, and therefore, that an EVLP protocol of 4.5 h is better suited for optimal antioxidant defences.
Limitations
Our study has several limitations. First, we did not assess the influence of different post-HS recovery times during EVLP after transplantation. The primary aim of our study was to address the chronology of events set in motion by HS application during the ex vivo procedure. Future experiments will be needed to determine whether the time-dependent effects of HS ex vivo translate into different outcomes after LTx for an in-depth evaluation of this topic. Second, we detailed the effects of HS at only 2 distinct time-points (3 and 4.5 h). Importantly, our objective was to determine the duration of EVLP after HS application that would maintain the lungs in a functionally intact state while ensuring an optimized expression of HSPs. Such conditions were better fulfilled after 4.5 h than after 3 h EVLP, while after 6 h, the protective effects of HS could not be fully maintained. However, even though the 4.5 h EVLP provided superior outcomes when compared to 3 and 6 h, we cannot exclude that any durations of EVLP within the 3–6-h range could have provided optimal performance.
In summary, our study presents the first evidence of the critical role of post-stress recovery time in the context of therapeutic heat stress for ex vivo reconditioning of damaged lungs. Thus, when applying a transient HS after 1 h EVLP, we could demonstrate that a post-HS recovery time of 180 min (4.5 h EVLP) provides a larger benefit than 3 h or 6 h EVLP in terms of HSP induction, expression of endogenous homeostatic proteins and the induction of pro-survival and antioxidant defences, while maintaining the lungs in a state of physiological integrity. Whether a better performance of HS reconditioning could be achieved using different durations of EVLP (between 3 and 6 h) remains to be established. These findings extend the concept of ex vivo heat stress application as a non-pharmacological therapy for the rehabilitation and reconditioning of damaged lung grafts.
Supplementary Material
ezaf027_Supplementary_Data
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Cypel M , Yeung JC, Liu M et al Normothermic ex vivo lung perfusion in clinical lung transplantation. N Engl J Med 2011;364:1431–40.21488765 10.1056/NEJ Moa 1014597 · doi ↗ · pubmed ↗
- 2Nilsson T , Wallinder A, Henriksen I et al Lung transplantation after ex vivo lung perfusion in two Scandinavian centres. Eur J Cardiothorac Surg 2019;55:766–72.30376058 10.1093/ejcts/ezy 354PMC 6421510 · doi ↗ · pubmed ↗
- 3Cypel M , Keshavjee S. Extending the donor pool: rehabilitation of poor organs. Thorac Surg Clin 2015;25:27–33.25430427 10.1016/j.thorsurg.2014.09.002 · doi ↗ · pubmed ↗
- 4Watanabe T , Cypel M, Keshavjee S. Ex vivo lung perfusion. J Thorac Dis 2021;13:6602–17.34992839 10.21037/jtd-2021-23PMC 8662477 · doi ↗ · pubmed ↗
- 5Singh MK , Shin Y, Ju S et al Heat shock response and heat shock proteins: current understanding and future opportunities in human diseases. Int J Mol Sci 2024;25:4209.38673794 10.3390/ijms 25084209 PMC 11050489 · doi ↗ · pubmed ↗
- 6Ojanguren A , Parapanov R, Debonneville A et al Therapeutic reconditioning of damaged lungs by transient heat stress during ex vivo lung perfusion. Am J Transplant 2023;23:1130–44.37217006 10.1016/j.ajt.2023.05.009 · doi ↗ · pubmed ↗
- 7Parapanov R , Debonneville A, Allouche M et al Transient heat stress protects from severe endothelial damage and dysfunction during prolonged experimental ex vivo lung perfusion. Front Immunol 2024;15:1390026.38807604 10.3389/fimmu.2024.1390026 PMC 11130382 · doi ↗ · pubmed ↗
- 8Fulda S , Gorman AM, Hori O, Samali A. Cellular stress responses: cell survival and cell death. Int J Cell Biol 2010;2010:214074.20182529 10.1155/2010/214074 PMC 2825543 · doi ↗ · pubmed ↗