Liposome-Mediated MicroRNA Delivery: An Additional Layer of Gene Network Regulation and Nuclear Reprogramming
Navid Ghasemzadeh, Fatemeh Pourrajab, Ali Dehghani Firoozabadi, Maryam Rahnama

TL;DR
This study explores using liposomes to deliver microRNAs for cell reprogramming, showing how they can regulate genes linked to pluripotency.
Contribution
The study introduces liposome-mediated miRNA delivery as a non-transduction method for cell reprogramming.
Findings
miR-302a and miR-34a regulate pluripotency factors OCT4, SOX2, and NANOG.
Lipoplexes with miR-302a increased OCT4 expression, while miR-34a decreased it.
LP-miRs can induce pluripotency precursors and influence cell lineage conversion.
Abstract
Developing miRNA-mediated cell engineering introduces a novel technology for cell reprogramming and generating patient-specific tissues for therapeutic use, facilitating basic research on human adult stem cells. Furthermore, optimizing a reprogramming method without transduction minimizes the risk of tumorigenesis, especially for reprogrammed cells. This study aimed to explore the use of liposomes as vehicles for delivering miRNAs to cells, focusing on their role in regulating gene networks and facilitating nuclear reprogramming. This study utilized cationic liposomal nanoparticles preserved under different conditions to introduce miRNAs into hMSCs. Using qPCR, the effective induction of pluripotency factors (OCT4, SOX2, and NANOG) was examined. Results indicated that miR-302a and miR-34a regulate pluripotency by interacting with key transcription factors, including OCT4, SOX2, and…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
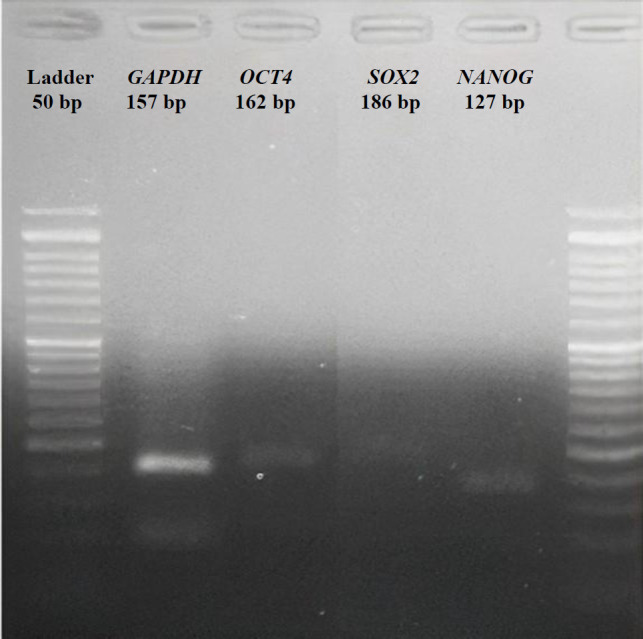 Fig. 1
Fig. 1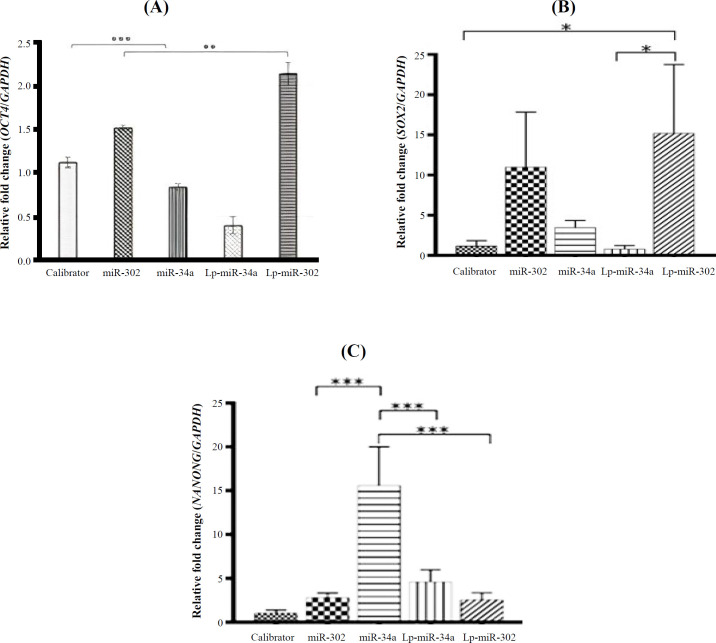 Fig. 5
Fig. 5|
|
|
|
|
|---|---|---|---|
| hsa-miR-302a-5p | MIMAT0000683 | 5’UAAGUGCUUCCAUGUUUUGGUGA3’ | Chr4 |
| hsa-miR-34a-5p | MIMAT0000255 | 5’UGGCAGUGUCUUAGCUGGUUGU3’ | Chr1 |
|
|
|
|
|
|
|---|---|---|---|---|
|
| 5'-GCCAGGGTCTCTCTTTCTG-3' | 5'-AAGCTGCCCACCTAACTT-3' | 162 | |
|
| 5'-GCACAACTCGGAGATCAGCAA-3' | 5'- GGCAGCGTGTACTTATCCTTCTTC-3' | 186 | |
|
| 5'-GAACTCTCCAACATCCTGAACCTC-3' | 5'-CCTTCTGCGTCACACCATTGC-3' | 127 | |
|
| 5'-GAGCCACATCGCTCTGACAC-3' | 5'-CATGTAGTTGAGGTCAATGAAGG-3' | 157 |
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsRNA Interference and Gene Delivery · Advanced biosensing and bioanalysis techniques · MicroRNA in disease regulation
INTRODUCTION
MicroRNAs are small non-coding RNAs that play a crucial role in gene regulation and nuclear reprogramming. These molecules can regulate the stability of nuclear transcripts and induce epigenetic alterations that silence or activate transcription at specific loci^[^^1^^,^^2^^]^. The miRNAs have been considered valuable tools for therapeutic strategies, as they can induce tissue-specific functions^[^^1^^,^^3^^]^. One promising method for delivering miRNAs to target cells is the use of liposomes, spherical vesicles composed of a lipid bilayer.
Liposomes can encapsulate and deliver miRNAs to target cells, offering several advantages over viral delivery systems, including low immunogenicity and toxicity^[4]^. The systemic delivery of miRNA via miRNA-loaded liposomes has emerged as an attractive target for therapeutic intervention in treating human diseases^[^^5^^]^. Targeting miRNAs may be a promising strategy to enhance the survival rate of engrafted stem cells and improve therapeutic outcomes^[^^6^^,^^7^^]^.
Research has demonstrated the reprogramming of the differentiated somatic cells into iPSCs through the ectopic expression of the pluripotency factors Oct4, Sox2, Klf4, and c-Myc^[^^8^^]^. However, a recently introduced approach for cell reprogramming/ engineering has involved a brief exposure of somatic cells to ectopic pluripotency/apoptosis-associated miRNAs^[^^9^^,^^10^^]^. Data exhibited that miRNAs can influence the dynamic equilibrium of various functional states of somatic cells, leading to the conversion of cells into specific cell types^[11]^. It has been well-known that the reprogrammed somatic cells overexpress OCT4 or miR-302a, causing the transition from a differentiated state to a more plastic state. However, the exact molecular mechanisms underlying this transition have not yet been understood. The shift from a differentiated state to a plastic state requires the acquisition of stem cell characteristics. At this stage, the chromatin is epigenetically in the “open” state, and cells are permissive to fate transition in response to external stimuli^[^^6^^,^^12^^]^.
The miR-302a can also drive the induction of cell plasticity, leading to the de-differentiation and the acquisition of a stem cell-like/plastic state that is permissive to external stimuli for differentiation^[^^13^^]^. Conversely, the apoptosis-associated miR-34a induces neural stem cell differentiation or acts as a tumor suppressor, reducing stemness and enhancing the cytotoxic susceptibility of neuroblastoma, breast cancer stem cells, and squamous carcinoma cells^[14]^. Additionally, miR-34a acts a key role in rescuing human bone marrow mesenchymal stem cells from stressful conditions^[^^15^^]^. Although the molecular mechanisms involved in the reprogramming process are not completely clear, studies have suggested that pluripotency factors such as OCT4, SOX2, and NANOG are essential for efficient reprogramming^[^^16^^,^^17^^]^. In this study, cationic liposomes were prepared as vectors for miRNA delivery into hMSCs, and the miRNA loading efficiency, as well as the physical stability of the prepared liposomes, were examined. This study was designed to provide a method for generating human iPSCs, clarify some molecular aspects involved in reprogramming human cells, and screen drug mechanisms.
MATERIALS AND METHODS
Materials
DSPE-PEG and soybean phosphatidylcholine were purchased from Lipoid GmbH (Germany). Cholesterol, MTT, NAHCO_3_, and DiI were acquired from Sigma-Aldrich (USA) and DOTAP from Avanti Polar Lipids (USA). The mature miRNA sequences were obtained through a search in the miRBase database (http://www.mirbase.org). The ready-to-use miRNA oligonucleotides related to the miR-302a and miR-34a sequences (Table 1) were provided by Qiagen (Germany). DMEM with low glucose, Glutamax® supplement, pyruvate, PBS tablets, penicillin/ streptomycin/amphotericin B, and trypsin-EDTA were procured from Gibco (USA). Fetal bovine serum and the hMSC line S1939 were purchased from Invitrogen (USA) and Royan Institute (Iran), respectively.
Preparation of cationic liposomes and lipoplexes
Cationic liposomes were prepared using a previously well-explained method. Briefly, the liposome vesicles were prepared from lipid mixtures of DPPC, cholesterol, and DOTAP in a molar ratio of 70:20:30, with an additional 5% DSPE-PEG. Chloroform was used to dissolve the lipid phase, which was then dried to form a thin film. Thereafter, an ammonium sulfate solution was added and sonicated at 65 °C for 30 min to perform the hydration stage. By dialysis at 25 °C for 2 h, the ammonium sulfate containing small unilamellar vesicles was replaced with PBS. To prepare lipoplexes (Lp-miRNAs) or miRNA-liposome complexes, we incubated liposomes with miRNA at room temperature for 30 min^[18,19].^
Physiochemical characterization of nano-lipoplex vesicles
The protocol of nano-lipoplex vesicle characterization was performed as described previously^[18]^. Briefly, the prepared particles were diluted in deionized water. The size, hydrodynamic diameter, PDI, and surface zeta potential were determined using dynamic light scattering on a Zeta PALS instrument from Brookhaven (USA), following the manufacturer's instructions. The mean and standard deviations of the measured parameters were calculated based on three trials conducted in four replicates^[^^18^^]^.
Loading efficiency
To determine the miRNA loading, different concentrations of naked miRNA were added to cationic vesicles (10-100 μg of miRNA per 1 mg of lipid [12-15 μl]), to prepare the lipoplexes/Lp-miRNAs. Subsequently, electrophoresis using agarose gel (2%), and ethidium-bromide staining was applied to determine the miRNA loaded by cationic liposomes. The images were obtained using a gel-documentation system (UVP, Cambridge, UK). Besides, free liposome as a control and different ratios of liposome to miRNA (15, 12.5 to 0.9) were analyzed to select the most appropriate formula with the greatest capacity of miRNA loading. For this purpose, 4 μl of each suspension was mixed with 1 μl of 4× DNA-loading buffer (Biolabs)^[^^18^^]^.
Morphology observation
The SEM (KYKY EM3200, China) was used to examine the surface morphology of lipoplexes and determine the approximate size of the vesicles. Preparation of the samples and the coating method were performed as previously described^[18]^^.^ Briefly, thin and dried layers of samples were prepared and coated with a positively charged mixture of argon and gold ions, which were then physically deposited onto the surface of negatively charged samples. A sputter coater (SBC 12, KYKY, China) was utilized along with the vapor deposition method. The images were captured at a maximum voltage of 26 KV^[^^18^^]^.
Assessment of loading efficiency
The efficiency of miRNA loaded into the vesicles was calculated as described formerly^[18]^. In brief, free miRNAs were separated from lipoplexes by precipitating liposomes through centrifugation at 35,000 ×g at 4 °C. By utilizing UV spectroscopy (PG Instruments, UK) at a wavelength of 260 nm, the number of free miRNAs present in the supernatant was evaluated. The liposome-loading capacity was determined by subtracting the amount of free miRNA that was not loaded into the liposomes from the total amount of miRNA initially introduced into the solution.
Thermal stability of the prepared lipoplexes
The thermal stability method was employed to assess the physical stability of lipoplexes. The stability of bilayer vesicles containing entrapped miRNAs was measured after 48 h of incubation at 4, 25, 37, and 42 °C. As mentioned previously^[18]^, the gel (2%) electrophoresis method was used to evaluate the thermal stability of lipoplexes, particularly their ability to retain the entrapped miRNAs as a function of rigidity^[^^18^^]^.
hMSC culture
The cells were cultured in DMEM (Gibco) containing FBS (10%), a mixture of penicillin, streptomycin, and amphotericin B (100 U, 100 µg, and 2.5 µg per mL, respectively) in a 5% CO_2_ humidified incubator at 37 °C. The cells were then passaged as a monolayer in adherent culture when they reached 80-90% confluence.
hMSC treatments
We used a previously described protocol for treating hMSC^[18]^. In brief, cultures were treated with empty liposomes, liposomal miRNAs (Lp-miR-302a and Lp-miR-34a), free miRNAs, and negative control. Cells for all experiments were used when they were in their exponential growth phase, and each treatment was performed four times.
Cytotoxicity assay
Cytotoxicity assay was carried out using the MTT test method. To this end, hMSCs were seeded in 96-well plates and incubated for 24 h. The cells were then treated with different concentrations of each formulation and incubated for 48 h. Afterward, 20 μl of MTT (5 mg/mL) was added to each well and incubated for a further 3 h. Following incubation, the supernatant was removed, and 180 μl of DMSO was added to dissolve the crystals. Absorption was measured using a Synergy TMHT multi-mode microplate reader (BioTek Instruments, USA)^[^^18^^]^.
Localization of fluorescent-labeled lipoplexes
The protocol for localization of fluorescent-labeled lipoplexes was conducted as explained before^[18]^. In brief, the cells were seeded in six-well plates (5 × 10^5^ cells/well) overnight. The culture medium was then replaced with DMEM lacking FBS. Next, Lp-FAM-miRNA was added to each well at a concentration of 100 nM and incubated for 4 h. The cells were subsequently transferred into 2 mL of a fresh medium containing 10% FBS. All experiments were performed in triplicate. For DAPI staining, treated cells were washed with PBS and fixed with paraformaldehyde. A DAPI solution (0.125 μg/mL) was applied for 15 minutes for nuclear counterstaining. To visualize the uptake, a fluorescence microscope (Olympus, Japan) was utilized was used.
Cell treatment and RNA extraction
The third passage of hMSCs was treated with the prepared formulations, and RNA was extracted according to the kit instructions (RiboExTM kit; GeneAll Biotechnology, Germany). To measure the RNA quantity, we employed a Nanodrop 2000 instrument from Thermo Fisher Scientific (USA). This device determined the concentration of the samples, as well as assessed the quality and purity of total RNAs by calculating the absorbance ratios. Gel electrophoresis assays were also used to qualify and analyze the extracted RNAs^[^^18^^]^.
Reverse transcription and cDNA synthesis
In the reverse transcription process, cDNAs were synthesized from extracted RNAs using the RevertAid First Strand cDNA Synthesis Kit (Thermo Fisher Scientific, USA) and the BioRad T100 Thermal Cycler (USA). first-strand cDNA was amplified following the program outlined previously^[^^18^^]^: 5 min at 25 °C, 60 min at 42 °C, and 5 min at 70 °C. The RT-PCR products corresponding to each primer were then subjected to agarose gel electrophoresis (2%) to resolve the amplicons (Fig. 1).
Quantitative RT-PCR
To determine each mRNA level or the expression of each target gene (OCT4,* SOX2*,* NANOG*, and PARP1), qPCR was performed using HOT FIREPol® EvaGreen® qPCR Mix Plus kit (Solis BioDyne, Tartu, Estonia) and primer sequences (Table 2). To perform qPCR and calculate the relative expression of target genes, we utilized the StepOne™/StepOnePlus™ Software and Real-Time PCR System (Applied Biosystems, Thermo Fisher Scientific). The qPCR procedure was conducted following the methodology explained in a previous study^[18]^. Reactions were performed in duplicates, with a final volume of 20 μL using cycling parameters (3 min at 95 °C, 3 s at 95 °C, and 20 s at 60 °C, the latter two steps repeated for 40 times). The expression levels were normalized against the housekeeping gene, GAPDH, and relative changes were calculated by the ΔΔCT formula. The primer sequences used in QPCR qPCR are depicted in Table 2. due to neutralization. For both formulations, miRNA incubation increased liposome diameters and decreased PDI. The decrease in zeta potential (~60%) indicated efficient miRNA loading. SEM images showed no differences between lipoplexes, displaying a spherical shape and a homogeneous size distribution of about 100-140 nm^[^^18^^]^.
Statistical analysis
The results were presented as the means ± SD and analyzed by SPSS software. Student t-test or one-way ANOVA was performed to assess differences and significance. A p ≤ 0.05 was considered statistically significant.
DISCUSSION
Reprogramming and engineering of somatic cells hold significant importance in research and cellular therapy strategies. Studies in this field offer insights into the molecular mechanisms involved in generating reprogrammed cells and the pathogenesis of diseases^[^^7^^,^^22^^]^. Cell engineering or the induction of plasticity through applying miRNAs, is a complex process that involves numerous genes and signaling pathways^[^^23^^,^^24^^]^. Ectopic miRNAs have emerged as powerful tools for cell reprogramming and generating patient-specific cell types^[^^25^^,^^26^^]^, as they can effectively engineer cells with reduced stress^[^^18^^]^.
The endogenous expression of miR-302a is crucial for maintaining pluripotency and plays a significant role in the reprogramming and generating iPSCs by fine-tuning mRNA expression^[27]^. Conversely, miR-34a has attracted attention for its function as a tumor suppressor that inhibits stemness. Similar to miR-302a, miR-34a, as a tumor suppressor, influences cell fate through various pathways, including the regulation of cell cycle and apoptosis^[28]^.
Cationic liposomes (lipoplexes) have shown potential for the intracellular delivery of miRNAs as vectors and are often used for gene delivery. Incorporating cationic lipids impart a positive surface charge to liposomes, facilitating their interaction with the negatively charged cell membrane and resulting in more efficient and uniform cellular uptake^[5,29]^. Additionally, cholesterol, a neutral lipid, along with PEG, contributes to the stability of the liposome structure and improves transfection efficiency in vitro^[^^18^^,^^30^^]^.
In a recent study, hMSCs were transfected with miRNAs, which were delivered by cationic liposomes (Lp-miRNAs). The cationic-liposomal formulation included cholesterol, DSPE-PEG, and DOTAP, selected as the building blocks for miRNA delivery. The prepared lipoplexes (Lp-miRNAs) effectively entered the hMSCs. To evaluate the efficacy of transfection using the prepared lipoplexes, we investigated the expression of pluripotency genes in Lp-miRNA-treated hMSCs. Our findings indicated that miR-302a and miR-34a influenced pluripotency through its partners, SOX2, and NANOG. OCT4 may act cooperatively to open chromatin configurations at pluripotency loci, accelerate the reprogramming network, and induce the expression of SOX2 and NANOG^[11]^. It is assumed that *OCT4 *is the upstream regulator of SOX2 and NANOG during the early stages of nuclear reprogramming, facilitated by epigenetic modifications that precede the induction of pluripotency. Among OCT4, SOX2, and NANOG, OCT4 is a core transcriptional factor, essential for the self-renewal of somatic cells and the maintenance of pluripotency in hPSCs^[^^6^^,^^11^^]^.
MiRNAs possess a seed region, typically spanning 2-8 nucleotides, which bind to partially complementary sequences in the 3'-UTRs of target mRNAs. This binding leads to gene silencing through translational repression or mRNA degradation^[31]^. The tumor suppressor miR-34a demonstrates a negative correlation with pluripotency genes such as OCT4, SOX2, and NANOG while showing a strong positive correlation with essential factors such as NOTCH1, AGR2, and KLF4^[32]^. Notably, the upregulation of miR-34a, which is responsive to p53 signaling, has been associated with the concurrent downregulation of critical pluripotency markers, including NANOG, OCT4, and SOX2, across various studies. This regulatory mechanism underscores the role of miR-34a in modulating stem cell pluripotency and differentiation pathways^[^^11^^,^^33^^,^^34^^]^.
Research findings have revealed that the influence of miR-34a on megakaryocytic differentiation of K562 cells is independent of p53 activity. Specifically, the induction of miR-34a expression during megakaryocyte differentiation, triggered by phorbol ester, has been exhibited to suppress cell proliferation, induce G1 phase cell-cycle arrest and enhance megakaryocyte maturation^[^^35^^]^.
Within the framework of pluripotency, miR-34a is thought to act as a suppressor of NANOG, OCT4, and SOX2, potentially contributing to the maintenance of a differentiated cellular state. The negative correlation observed between the expression of miR-34a and the levels of pluripotency factors suggests that miR-34a plays a significant role in regulating stem cell properties and the process of cellular specialization^[^^27^^]^. Tumor-suppressor miRNAs, such as miR-34a, contain a G-rich toxic 6-mer seed sequence, which is highly enriched in genes that regulate the cell cycle, cell division, DNA repair, and nucleosome assembly^[7,11]^.
Liposomal-delivered miRNA may serve as a powerful tool in gene therapy for cancers. In contrast, ectopic miR-302 can fully reprogram iPSCs by increasing the expression levels of NANOG, SOX2, and OCT4. Both NR2F2 and MBD2 are exclusively expressed in differentiated cells and are direct targets of miR-302. MBD2 acts as an epigenetic suppressor, hindering the complete reprogramming of somatic cells to iPSCs by directly binding to the NANOG promoter and preventing its transcriptional activation^[7,24]^. The five CpG dinucleotides within the NANOG promoter can be methylated and silenced by MBD2^[11]^. Furthermore, bioinformatics analysis has identified a seven-base pair sequence in the 3'-UTR of AKT1 that is complementary to the sequence of miR-302^[6]^. The upregulated miR-302 can directly target AKT1 through its 3'-UTR, thereby maintaining a high level of the pluripotent factor OCT4 in hMSCs. In both pluripotent and adult stem cells, Akt functions as a suppressor of OCT4 expression^[36,37]^. In turn, the transcription factors OCT4, SOX2, and NANOG upregulates miR-302 by binding to a conserved region in the miR-302 promoter, promoting its expression or vice versa^[6,12,38]^. This positive feedback loop between miR-302 and the pioneer transcription factors (OCT4, SOX2, and NANOG) is proposed to represent a new mechanism for understanding the induction of pluripotency in somatic cells.
Agarose gel (2%) electrophoresis of RT-PCR products corresponding to each target gene (GAPDH, OCT4, SOX2, and NANOG).
*Relative fold change in the expression levels of (A) OCT4, (B) SOX2, and (C) NANOG after transfection with each miRNA formulation. Calibrator as a control, miR-302a, Lp-miR302a (lipoplex), miR-34a, Lp-miR-34a (lipoplex). The 2-ΔΔCt method was employed. Error bars represent SD (*p<0.05, *p < 0.01, *** p < 0.001).
CONCLUSION
According to the findings obtained, miRNAs are located at the genetic-epigenetic interface of cellular events, where epigenetic alterations occur before the activation of pluripotency loci. Our hypothesis posits that the liposomal distribution of ectopic miRNAs would introduce an additional layer of gene-network control, serving as a tool to influence the decision cells make regarding their fate in response to external stimuli. During the early phases of nuclear reprogramming, epigenetic changes precede the transcriptional activation of pluripotency loci. A hypothesis suggests that a feedback loop exists between pioneer pluripotency factors and miRNAs, with epigenetic alterations occurring upstream. The findings of this study illuminate the role that miRNAs play in the genetic-epigenetic interface of pluripotency networks. These networks are crucial for fine-tuning transcriptional heterogeneity and, as a result, the evolutionary flexibility exhibited by human cells. Our findings expand the existing knowledge of ectopic miR-34a and miR-302a, clarifying their functions within cells and their significance in the regulation of gene networks.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Otoukesh B Abbasi M Gorgani HOL Farahini H Moghtadaei M Boddouhi B Micro RN As signatures, bioinformatics analysis of mi RN As, mi RNA mimics and antagonists, and mi RNA therapeutics in osteosarcoma Cancer Cell Int 2020202543256573810.1186/s 12935-020-01342-4PMC 7302353 · doi ↗ · pubmed ↗
- 2Carey BW Markoulaki S Hanna JH Faddah DA Buganim Y Kim J Reprogramming factor stoichiometry influences the epigenetic state and biological properties of induced pluripotent stem cells Cell Stem Cell 201196588982213693210.1016/j.stem.2011.11.003 · doi ↗ · pubmed ↗
- 3Di Fiore R Suleiman S Pentimalli FO’toole SAO’leary JJ Ward MP Could Micro RN As be useful tools to improve the diagnosis and treatment of rare gynecological cancers? A brief overview Int J Mol Sci 202122838223391702210.3390/ijms 22083822 PMC 8067678 · doi ↗ · pubmed ↗
- 4Farooque F Wasi M Mughees MM Liposomes as Drug Delivery System: An Updated Review J Drug Deliv Ther 202111514958
- 5Gao Y Liu X Chen N Yang X Tang F Recent advance of liposome nanoparticles for nucleic acid therapy Pharmaceutics 20231511783667880710.3390/pharmaceutics 15010178 PMC 9864445 · doi ↗ · pubmed ↗
- 6Li HL Wei JF Fan LY Wang SH Zhu L Li T Pmi R-302 regulates pluripotency, teratoma formation and differentiation in stem cells via an AKT 1/OCT 4-dependent manner Cell Death Dis 201671 e 20782682107010.1038/cddis.2015.383PMC 4816169 · doi ↗ · pubmed ↗
- 7Gao QQ Putzbach WE Murmann AE Chen S Sarshad AA Peter JM 6mer seed toxicity in tumor suppressive micro RN As Nat Commun 20189145043037411010.1038/s 41467-018-06526-1PMC 6206098 · doi ↗ · pubmed ↗
- 8Kizub I Rozhok A Bilousova G Induced Pluripotent Stem Cells: Advances and Applications in Regenerative Medicine 2022