Anti-Inflammatory, Cytotoxic, and Anti-Tuberculosis Properties of Selected Fabaceae Medicinal Plants through in vitro Studies
Mercy Mpho Khala, Anneke van der Spoel van Dijk, Polo-Ma-Abiele Hildah Mfengwana

TL;DR
This study investigates three South African medicinal plants for their anti-inflammatory and anti-TB properties, finding some potential for use in managing inflammation but not directly treating TB.
Contribution
The study provides new in vitro evidence on the anti-inflammatory and cytotoxic properties of three indigenous Fabaceae plants, despite no observed anti-TB activity.
Findings
E. elephantina and T. burchellianum aqueous extracts showed anti-inflammatory activity at 200 μg/ml.
E. elephantina extracts were non-cytotoxic at 200 μg/ml, while L. lanceolata and T. burchellianum were cytotoxic.
None of the plant extracts exhibited anti-TB activity against LAM and EAI strains.
Abstract
Tuberculosis is among the top ten causes of death in South Africa and worldwide and remains a high priority of the World Health Organization. South Africa has a rich tradition for using medicinal plants to treat various diseases, including TB; however, the safety and efficacy of these plants require thorough investigation and confirmation. This study examined the anti-inflammatory and anti-TB activities of three indigenous medicinal plants against two strains of TB. Water and methanol root extracts of E. elephantina (Burch.), L. lanceolata, and T. burchellianum were tested in vitro for their activity against LAM and EAI lineages. Additionally, the phytochemical screening, anti-inflammatory properties, and cytotoxicity activities of these extracts were evaluated. Aqueous extracts of E. elephantina and T. burchellianum exhibited anti-inflammatory activity at a concentration of 200…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
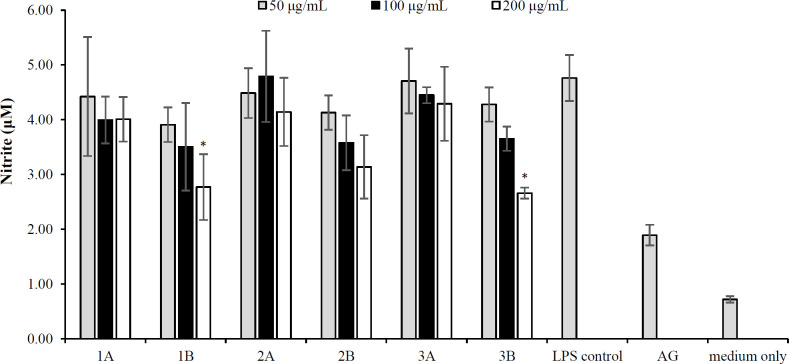 Fig. 1
Fig. 1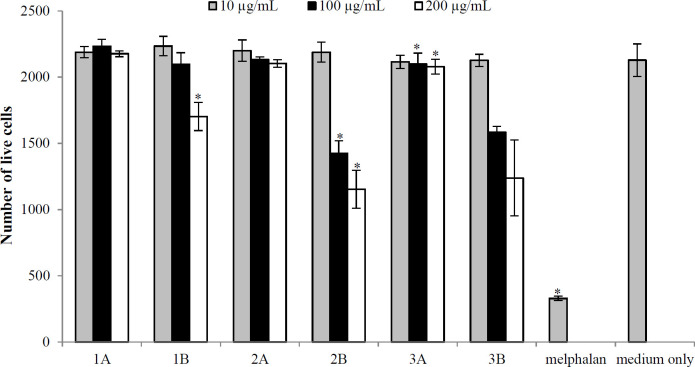 Fig. 2
Fig. 2|
|
|
|
|
|
|---|---|---|---|---|
|
| ++ | ++ | - | ++ |
|
| + | + | - | + |
|
| + | + | - | + |
|
|
|
|
|
|
|
|---|---|---|---|---|---|
| 50 | MeOH | + | + | ||
|
| H2O | + | + | ||
| LAM | 100 | MeOH | + | + | |
| H2O | + | + | |||
|
| 200 | MeOH | + | + | |
| H2O | + | + | |||
| 50 | MeOH | + | + | ||
| H2O | + | + | |||
|
| EAI | 100 | MeOH | + | + |
| H2O | + | + | |||
| 200 | MeOH | + | + | ||
| H2O | + | + |
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsNatural Antidiabetic Agents Studies · Ethnobotanical and Medicinal Plants Studies · Natural product bioactivities and synthesis
INTRODUCTION
Tuberculosis is a contagious disease that spreads through the air and is caused by M. tuberculosis^[^^1^^]^. It ranks among the top ten deadliest diseases globally^[^^2^^]^. TB is transmitted when an individual develops active TB. In other words, it begins to exhibit symptoms and expels the Mycobacterium into the air by coughing, laughing, or talking, allowing another person to inhale the bacteria. The disease primarily affects the lungs, resulting in pulmonary TB^[^^3^^]^. TB is a curable disease that can be treated with a six-month drug regime, which includes isoniazid, rifampicin, ethambutol, and pyrazinamide^[^^4^^]^. However, these first-line medications often have adverse side effects that often lead patients to discontinue treatment. Given this limitation and the increasing prevalence of M. tuberculosis strains, which are widespread and resistant to multiple drugs, there is an urgent need for further research focusing on developing new medications that can effectively treat and cure these resistant strains while minimizing side effects^[^^5^^]^.
Inflammation plays a critical role in TB infection. When M. tuberculosis enters the body, the immune system mounts an inflammatory response to control and eradicate the disease. This response involves the activation of immune cells, primarily macrophages, which release inflammatory cytokines to recruit more immune cells to the site of infection. Chronic inflammation associated with TB can cause tissue damage, the formation of granulomas including small, organized clusters of immune cells, and, in severe cases, the development of lung cavities^[^^1^^]^. These inflammatory processes can contribute to the progression of active TB and increased risk of transmitting the bacteria to others. Balancing the immune response to prevent excessive inflammation while combating the infection is a critical challenge in TB research and treatment^[^^5^^]^.
The majority of African countries rely on traditional medicinal remedies to treat pulmonary TB due to limited access to Western medical treatments^[^^6^^]^. Medicinal plants are cost-effective and naturally occurring sources with minimal side effects. They are recognized as reservoirs of novel active compounds that offer pharmacological benefits^[^^7^^]^. The extraordinary diversity of biological activity of medicinal plants makes them useful sources for both conventional and modern medicine. Numerous chemical substances that affect the human body can be found in plants. Due to their adaptability, plants are often utilized to treat various diseases^[^^8^^]^. E. elephantina, L. lanceolata, and T. burchellianum belonging to the Fabaceae family are traditionally used for treating illnesses, including TB^[^^9-12]^. In this regard, traditional healers in Lesotho utilize the roots of various plants to treat TB, as the roots of this plant contain phytochemicals that are essential for TB treatment.
In this investigation, we selected three plant species— E. elephantina,* L. lanceolata*, and* T. burchellianum*— to explore the ethnobotanical claims regarding medicinal plants with potential anti-TB activity. Our study demonstrated the anti-inflammatory, cytotoxicity, and anti-TB effects of these plants on two strains of TB.
MATERIALS AND METHODS
Plant material and extraction
The roots of *E. *elephantina, L. lanceolate, and T. burchellianum were purchased from Random Harvest Nursery in South Africa. The roots were washed with water to remove the soil, dried at room temperature (22 °C), and crushed into fine powder. Plant extraction was performed using the maceration method. Pulverized plant materials were soaked respectively in 100% MeOH and dH_2_O on a shaker with occasionally stirring for 48 hours. The mixtures were filtered, and fresh solvents (100% MeOH and dH2O) were added to the sediment collected from the filter. This process was repeated until the collected supernatant became clear. The organic and aqueous extracts were concentrated using a rotatory evaporator and a freeze dryer, respectively. The organic extracts were then placed in an incubator at 37 °C for five days to dry.
Secondary metabolites screening
The plant extracts were analyzed for the presence of flavonoids, tannins, alkaloids, and terpenoids using a previously described method^[13]^. In brief, to identify flavonoids, we mixed the extract of each plant with a few drops of aluminum chloride solution. The presence of flavonoids was indicated by a yellow coloration, resulting from the formation of a complex between the flavonoids and aluminum ions. To detect tannins, we added a few drops of 5% ferric chloride solution to the plant extract. The formation of a blue-black or greenish-black colored precipitate confirmed the presence of tannins, as they react with the ferric ions. Alkaloids were distinguished by treating the plant extract with the reagent of Dragendorff, which consists of bismuth nitrate and potassium iodide, indicating by a reddish-brown or orange precipitate. Finally, to identify terpenoids, we mixed the plant extract with chloroform, and a few drops of concentrated sulfuric acid were added to the mixture, resulting in the formation of a reddish-brown interface layer, confirming the presence of terpenoids.
Anti-inflammatory assay
The assay was performed as described previously with slight modifications^[14]^. Extracts from three selected plants were solubilized using DMSO to create a stock solution of 100 mg/ml. DMSO was chosen as it enhances solubility, improves cellular uptake of bioactive compounds, and maintains low toxicity at an appropriate concentration, ensuring effective delivery and stability of the extracts in cell culture assays. The mouse macrophage cell line, RAW 264.7, was seeded in RPMI1640 culture medium supplemented with 10% FBS into 96-well plates at a density of 1 × 10^5^ cells per well and allowed to attach overnight. The microtiter plates were incubated in 5% CO_2_ at 37 °C overnight. The following day, the culture medium was removed, and the samples diluted in RPMI complete medium were added to reach the final concentrations of 50, 100, and 200 μg/ml. To assess the inflammatory activity, 50 μl of LPS (final concentration of 500 μg/ml)-containing medium was added to the corresponding wells. LPS is an endotoxin that can trigger a strong immune response in cells. Aminoguanidine was used as the positive control at a concentration of 100 μM. Cells were incubated for an additional 24 hours. To quantify NO production, 50 μl of the culture medium was transferred to a new 96-well plate, and 50 μl of Griess reagent was added. Absorbance was measured at 540 nm. A standard curve using sodium nitrate dissolved in an RPMI-1640 culture medium was determined the NO concentration in samples.
In vitro cytotoxicity assay
The African green monkey kidney cell line, Vero cells, was used for cytotoxicity screening, following a formerly described method with slight modifications^[^^15^^]^. The growth medium consisted of DMEM supplemented with 10% FBS. Cells were maintained in 10 cm culture dishes and incubated in a 5% CO_2_ atmosphere at 37 °C. Cells were plated in 96-well microtiter plates at a density of 4,000 cells per well, in which each well contained 100 μl of medium. The microtiter plates were incubated in 100% humidity and 5% CO_2 at 37 °C before adding test compounds, allowing cell attachment. The cytotoxicity of extracts was assessed using the Hoechst 33342 and propidium iodide dual staining methods. The extracts were tested at 50, 100, and 200 µg/ml concentrations. To perform this process, a 400 μg/ml dilution of each plant extract was prepared in a complete medium, consisting of DMEM supplemented with 10% FBS. A three-point dilution series was tested, resulting in a serial dilution of 100, 200, and 400 µg/ml. Next, aliquots of 100-μl dilutions were prepared. The aliquots of each dilution were added to 100 μl of the attached cells in a 96-well plate, yielding the final concentrations of 50, 100, and 200 µg/ml. The cells were incubated in 5% CO_2 at 37 °C for 48 hours, and melphalan was used as positive control at 20 μM. To confirm toxicity, cell viability was assessed using the MTT assay on Vero cells. This procedure involved removing the excess medium from each well that remained after the cytotoxicity assay was completed, followed by the addition of fresh medium containing 0.5 mg/mL MTT and incubating at 37 °C for an additional 30 minutes. After removing the MTT solution, 100 μl of DMSO was added to each well to dissolve the formazan crystals. Absorbance was then measured at 540 nm using a BioTek® PowerWave XS spectrophotometer (Winooski, Vermont, USA).
Anti-TB assay
Anti-TB susceptibility assay was conducted using the BATEC MGIT™ 960 system following a previously established method^[16]^. The H37Rv strain was utilized as susceptible control. Additionally, two well-characterized strains from different lineage families, lineage 2 (EAI) and lineage 4 (LAM3), were included as susceptible isolate strains. Aseptically, a fresh MGIT tube was prepared for each strain by adding 0.8 ml of BBL MGIT™ OADC™ growth supplement to the labelled MGIT tubes. A 1:5 dilution was created by adding 1 ml of the culture solution to 4 ml of sterile saline. The tubes were vortexed for 30 seconds and allowed to settle for one minute. Afterwards, 0.5 ml of the 1:5 dilutions was added to each prepared tube. The tubes were capped and mixed by inversion. The TB strains were loaded onto the MGIT™ 960 system and monitored daily. The MeOH and dH_2_O extracts of the plants were reconstituted in 4 ml of DMSO to prepare a stock solution. The GC was prepared by adding 100 μl of inoculum to 10 ml of sterile saline to achieve a final dilution of 1:100. A 500 μl volume of the prepared growth control was inoculated into the MGIT™ GC without any plant extracts. Then, 100 μl of each plant stock solution was added to the MGIT containing 7 ml of growth medium, 0.8 ml of MGIT™ OADC™, and 0.5 ml of the TB strain inoculum to achieve a final concentration of 200, 100, and 50 μg/ml. All tubes were incubated in the MGIT™ 960 system at 37 °C and monitored for growth until the system detected a growth result. The BACTEC MGIT™ 960 instruments indicated the completion when the growth control tubes reached a growth unit value exceeding 400. A control was also included without the addition of the plant extracts. The results were reported as susceptible when the growth unit values for the tubes containing drugs (plant extract) were ≥400 and <100 and as resistant when the growth unit value of the drug tube was ≥100.
Statistical analysis
Statistical analysis was performed using Excel. Each experiment was conducted in quadruplicate. Statistical significance was determined using the two-tailed student t-test, and p<0.05 was deemed statistically significant.
RESULTS
Phytochemical screening
The results of the phytochemical screening for the presence of flavonoids, tannins, alkaloids, and terpenoids in root extracts of E. elephantina,* L. lanceolata*, and* T. burchellianum *are represented in Table 1.
Anti-inflammation activity
The RAW 264.7 mouse macrophage cell line, a widely used and well-established model, is often employed to study the anti-inflammatory effects of test samples. Cells were cultured in multi-well plates and stimulated with bacterial LPS to activate inducible NO synthase expression, producing NO. Nitrite levels in the culture medium were measured to assess changes in NO production. The extracts were screened for their anti-inflammatory properties using RAW 264.7 cells. As depicted in Figure 1, all three plants promoted NO production at levels similar to those of LPS, which was used as a negative control. However, the water extracts of E. elephantina and *T. burchellianum *at a concentration of 200 µg/ml displayed lower NO production values. These plants indicated promising anti-inflammatory activity at the highest tested concentration (200 μg/ml).
Cytotoxicity activity
The cytotoxicity of six samples was assessed using Vero cells. Figure 2 shows processed live cell data after 48 hours of treatment. The results indicated cytotoxicity of the water extracts of L. lanceolata t and T. burchellianum at a concentration of 200 μg/ml when applied to Vero cells. Cytotoxicity was evidenced by a reduction in Vero cells growth compared to the control group, treated with medium only; however, the cytotoxicity was less than that of melphalan (positive control). The other plant extracts indicated no signs of toxicity. Cell viability (the number of live cells) remained above 80% in all extracts.
Anti-TB screening
TB-susceptible strains, particularly LAM and EAI, which are prevalent in South Africa, were used in this study. The susceptibility of E. elephantina***,*** L. lanceolate, and T. burchellianum extracts was tested against these two strains, as depicted in Table 2. Based on the anti-TB results, both dH_2_O and MeOH extracts of the three plant species exhibited no inhibitory activity against LAM and EAI TB strains at any of the concentrations tested.
DISCUSSION
Extracts with anti-inflammatory activities that effectively reduce the production of pro-inflammatory mediators can play a crucial role in managing chronic inflammation associated with TB. By mitigating the systemic inflammatory response, medicinal extracts could alleviate tissue damage and the exacerbation of symptoms commonly observed in TB, potentially improving patient outcomes and enhancing the effectiveness of conventional TB treatments. Targeting inflammation and pro-inflammatory pathways is essential, as this strategy can significantly decrease inflammatory activity in the lungs, the primary host organ. An ideal TB drug should also possess the ability to penetrate the intracellular environment for effective treatment^[17]^. Hence, understanding the complex interplay between inflammation and TB is crucial for developing effective treatments and interventions to manage the disease.
In the present study, we observed that aqueous extracts of E. elephantina and T. *burchellianum roots had significant anti-inflammatory activity at 200 μg/ml concentration. The NO production in RAW 264.7 cells also demonstrated the anti-inflammatory activity of plant extracts. Since these cells generated reduced levels of NO in response to plant extracts, we can suggest that these extracts have anti-inflammatory properties by which can potentially inhibit inflammatory pathways or mediators involved in NO synthesis. In this study, the anti-inflammatory activity of the plant extracts was determined by measuring changes in NO production levels in the culture medium. The observed anti-inflammatory activity could be attributed to the presence of flavonoids and anthraquinone, as previous studies have displayed that cell culture containing anthraquinone exhibits anti-inflammatory properties^[8]^. These findings corroborate the report by Olaokun et al. regarding the anti-inflammatory effects of E. *elephantina on skin cells^[^^18]^. However, in our study, L. lanceolata showed no anti-inflammatory activity. This behavior could arise from the low levels of flavonoids, tannins, and terpenoids present in this plant; the color intensity of the colorimetric assay was notably weaker for L. lanceolata compared to the other two plants tested. Cell viability was concurrently assessed using the MTT assay to ensure that the test samples were non-cytotoxic. The methanol and aqueous extracts of E. elephantina showed no cytotoxicity against Vero cells (non-cancerous cells). However, the aqueous extracts of L. lanceolata and T. burchellianum roots demonstrated a concentration-dependent cytotoxicity against these cells.
The plants E. elephantina,* L. lanceolata*, and* T. burchellianum*, belonging to the Fabaceae family, are traditionally used to treat various illnesses, including TB^[^^10^^-^^12^^]^. To support the ethnobotanical claims associated with these plants, we conducted anti-TB studies on these plant species to evaluate their effectiveness in inhibiting the growth of two selected TB strains. Despite the identification of secondary metabolites, such as flavonoids, tannins, and terpenoids, none of the extracts inhibited TB growth in all the plants studied. This lack of efficacy stems from the absence of alkaloids in the tested plant extracts, as alkaloids have been reported to be responsible for anti-TB activity through structural interaction with M. tuberculosis^[^^19^^]^. These results contradict the findings by Mpofu et al.^[^^9^^]^, who reported the presence of alkaloids in E. elephantina collected from Swaziland, South Africa, and Zimbabwe.
It is well-established that geographic location influences the secondary metabolites of plants, which is due to variations in soil conditions, light, H_2_O, CO_2_, temperature, and mineral nutrients^[20]^. The ability of plants to adapt to their environment and manage stress is significantly affected by these secondary metabolites. Adverse conditions such as drought, high salinity, frost, and extreme temperatures stimulate the production of secondary metabolites in plants. According to Yang et al., plants under stress conditions produce more secondary metabolites than their unstressed counterparts^[21]^. The absence of anti-TB activity in the plants tested in this study is likely due to their collection from a nursery as they are protected from stress and harsh conditions. Nursery-grown plants are typically less subjected to significant stress compared to those growing freely in their natural habitats, such as fields and mountains. This lack of stress could explain the absence of alkaloids in the plants tested. Additionally, other factors that may influence the anti-TB activity of the extracts include the failure of the extracts to disrupt the integrity of the mycobacterial cellular structure and environmental conditions that affect the interaction of the plant extracts with the LAM and EAI TB strains.
A possible limitation contributing to the lack of anti-TB activity observed in the selected plant extracts is related to the extraction process employed. Different solvents and extraction methods could yield varying concentrations of active compounds. Therefore, future studies should explore alternative extraction techniques, such as using solvents specifically extracting phyto-chemical groups known for anti-TB properties or adjusting extraction times and temperatures, as these modifications may produce varying outcomes. Furthermore, the current extraction process may not have effectively isolated the active alkaloids or other bioactive molecules, suggesting a need for refinement in the extraction protocols to enhance the anti-TB activity of the plants. It is important to note that testing the plant extracts against a wider range of TB strains and cell lines was not feasible due to financial issues; thus, only the two prominent South African strains were selected.
CONCLUSION
Investigation of the potential anti-inflammatory properties of E. elephantina,* T. burchellianum*, and L. lanceolata extracts have yielded insightful results, particularly concerning TB-related inflammation. The aqueous extract of E. elephantina has emerged as a promising candidate for anti-inflammatory drug development, showing effectiveness without inducing toxicity on Vero cells across various concentrations. While this safety profile is encouraging, it warrants further validation through in vivo studies to confirm its applicability for clinical use. On the other hand, T. burchellianum exhibited cytotoxicity and anti-inflammatory effects, suggesting a narrow therapeutic window that could be optimized in future formulations. Remarkably, none of the tested plants inhibited TB directly, as they lack alkaloids known for their anti-TB properties. However, the presence of other secondary metabolites indicates alternative therapeutic pathways that may support the management of TB symptoms by modulating inflammation. Considering the chronic inflammation characteristic of TB and the compromised immune systems of affected patients, the anti-inflammatory activities of E. elephantina and T. burchellianum may offer symptomatic relief and a supportive therapeutic strategy alongside conventional TB treatments. This holistic approach could be particularly beneficial in enhancing the quality of life of TB patients and treatment outcomes for TB patients. Further research is essential to characterize the extracts of these plants and fully explore their therapeutic potential and safety profile.
*NO production in LPS-activated macrophages treated with different concentrations of plant extracts. Bar graphs show quadruple values from a single experiment, with error bars indicating the standard deviation of the mean. 1: E. elephantina; 2: L. lanceolata; 3: T. burchellianum; A: MeOH extracts; B: H2O extracts (p < 0.05).
*Cytotoxicity activity of six extracts tested against Vero cells. The error bars indicate the standard deviation of quadruple values from a single experiment. 1: E. elephantina; 2: L. lanceolata; 3: T. burchellianum; A: MeOH extracts; B: H2O extracts (p < 0.001).
Table 2: E. elephantina, L. lanceolate, and T. burchellianum extracts against LAM and EAI TB strains
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Zumla A Rao M Parida SK Keshavjee S Cassell G Wallis R Inflammation and tuberculosis: host-directed therapies J Intern Med 20152774373872471709210.1111/joim.12256 · doi ↗ · pubmed ↗
- 2World Health Organization (WHO)Global Tuberculosis Report 2021 Available at: https://www.who.int/publications/i/item/9789240037021
- 3Long R Divangahi,M Schwartzman,K Chapter 2: Transmission and pathogenesis of tuberculosis Can J Respir Crit Care Sleep Med 2022 sup 12232
- 4Babalık A Ulus IH Bakirci N Kuyucu T Arpag H Dagyildiz L Pharmacokinetics and serum concentrations of antimycobacterial drugs in adult Turkish patients Int J Tuberc Lung Dis 20131711144272412544810.5588/ijtld.12.0771 · doi ↗ · pubmed ↗
- 5Imam F Sharma M Khayyam KU Al-Harbi NO Rashid MK Ali MD Adverse drug reaction prevalence and mechanisms of action of first-line anti-tubercular drugs Saudi Pharm J 2020283316243219433310.1016/j.jsps.2020.01.011PMC 7078525 · doi ↗ · pubmed ↗
- 6Ndhlala AR Stafford GI Finnie JF Van Staden J Commercial herbal preparations in Kwa Zulu-Natal, South Africa: The urban face of traditional medicine S Afr J 201177483043
- 7Mushtaq S Abbasi BH Uzair B Abbasi R Natural products as reservoirs of novel therapeutic agents EXCLI J 20184174205110.17179/excli 2018-1174 PMC 596290029805348 · doi ↗ · pubmed ↗
- 8Maroyi A Elephantorrhiza elephantina: Traditional uses, phytochemistry, and pharmacology of an important medicinal plant species in Southern Africa Evid-Based Complementary Altern Med 2017640390510.1155/2017/6403905 PMC 544688328588639 · doi ↗ · pubmed ↗