Changes in Inflammatory Cytokines, Vascular Markers, Cell Cycle Regulators, and Gonadotropin Receptors in Granulosa Cells of COVID-19 Infected Women
Sina Vakili, Amirabbas Rostami, Bahia Namavar Jahromi, Morteza Jafarinia

TL;DR
This study finds that women with recent COVID-19 infection show changes in genes related to inflammation, blood vessels, and cell cycles in their reproductive cells, which could affect fertility.
Contribution
The study is the first to report gene expression changes in granulosa cells of women with recent SARS-CoV-2 infection undergoing ART.
Findings
Inflammatory cytokines IL-1B, TNF-α, IL-6, and IL-8 were significantly upregulated in granulosa cells of women with recent COVID-19.
Vascular genes VEGF and ANGPT1 were overexpressed, while FOXL2 was downregulated and Cyclin D1/D2 were upregulated in infected women.
LH and FSH receptor expression remained unchanged between the groups.
Abstract
Background: COVID-19 infection can negatively affect multiple organ systems, including the reproductive system. Previous research has indicated altered levels of inflammatory markers in the reproductive tissues of women with chronic diseases. This study aimed to assess the expression of inflammatory, vascular, cell cycle, and gonadotropin receptor genes in the granulosa cells and oocytes of women with recent COVID-19 infection undergoing Assisted Reproductive Technology (ART), compared to healthy controls.Materials and Methods: The study involved 15 women who had tested positive for COVID-19 within three months of ART treatment and 15 age-matched healthy women as controls. Granulosa cells were collected during oocyte retrieval, and RNA was isolated to analyze gene expression using quantitative real-time PCR. The evaluated genes included inflammatory cytokines (IL-1B, TNF-α, IL-6, IL-8),…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
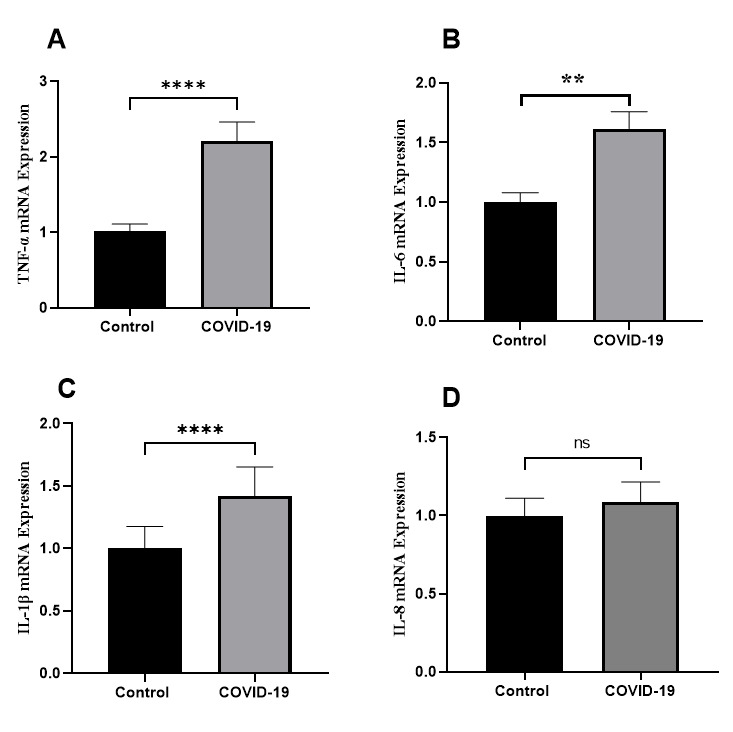 Figure-1
Figure-1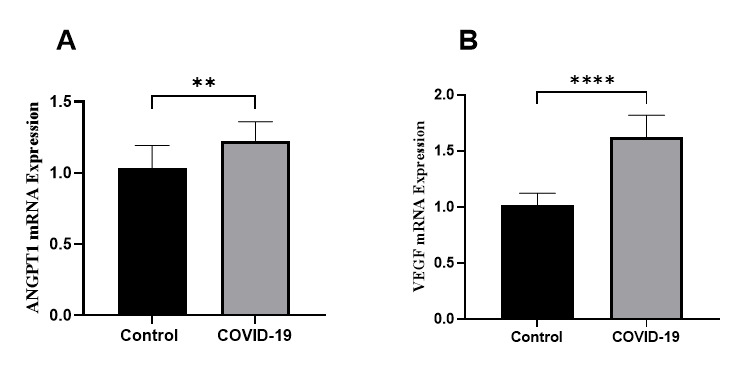 Figure-2
Figure-2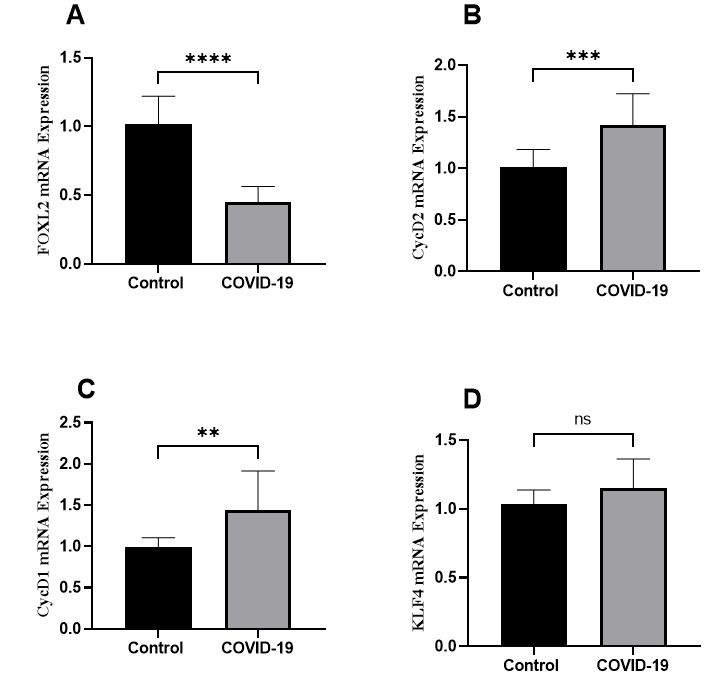 Figure-3
Figure-3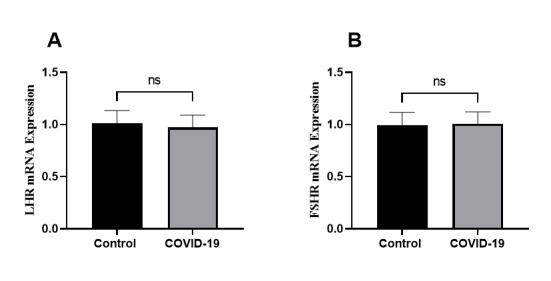 Figure-4
Figure-4Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsCOVID-19 Impact on Reproduction
Introduction
The COVID-19 pandemic, instigated by the SARS-CoV-2 virus, has become one of the most significant public health crises in contemporary history. Although the respiratory implications of the virus are well-documented, emerging research indicates that COVID-19 may also have repercussions for reproductive health [1][2][3][4]. The virus gains entry into host cells through the angiotensin-converting enzyme 2 (ACE2) receptor, which is present in several non-respiratory tissues, including the testes, ovaries, and placenta [5][6]. This presence raises alarms regarding the potential negative effects of COVID-19 on fertility and pregnancy outcomes. Numerous studies have indicated the presence of SARS-CoV-2 in various female reproductive tissues, including the ovaries, placental tissues, amniotic fluid, and breast milk of infected individuals [7][8][9]. This indicates that the virus may spread to the female reproductive system. Additionally, studies reveal elevated inflammatory cytokines in the serum and follicular fluid of women with COVID-19 who are undergoing in-vitro fertilization (IVF) [10][11][12]. Excessive inflammation caused by COVID-19 infection may hinder oocyte quality and fertility potential, as successful reproduction relies on a healthy ovulatory process and a well-balanced follicular microenvironment [13][14].
The female ovary is a complex organ that contains follicles at various stages of maturation. Each menstrual cycle involves the recruitment and selection of dominant follicles, culminating in the ovulation of a mature oocyte. This intricately regulated process relies on specific interactions among ovarian follicular cells, neuroendocrine signals, paracrine factors, and the developing oocyte [15].
Granulosa cells that line the follicles and the oocyte they surround engage in bidirectional communication to synchronize essential processes such as cell proliferation, differentiation, and apoptosis necessary for folliculogenesis. Disruptions in the molecular interactions between granulosa cells and oocytes can adversely affect the maturation and quality of the oocyte [16]. Previous studies have reported changes in the expression of inflammatory cytokines, growth factors, metabolic enzymes, and cell cycle regulators in the granulosa cells and oocytes of women with specific chronic conditions and obesity. These molecular changes were associated with impaired oocyte competence and reduced fertility outcomes [17]. Since COVID-19 causes a pathogenic infection with significant inflammatory response [18], it is likely that the disease alters the ovarian transcriptome too. Identifying the genes that are abnormally expressed in granulosa cells and oocytes could shed light on the mechanisms behind reproductive issues caused by COVID-19. This study aimed to thoroughly assess key genes involved in inflammatory, vascular, sex hormones, and cell cycle pathways in granulosa cells from women with COVID-19 undergoing assisted reproductive technology (ART), compared to healthy controls.
Materials and Methods
**
Study Participants
This study involved 30 women receiving assisted reproductive technology (ART) treatment split into two groups: a study group of 15 women who tested positive for COVID-19 after ovarian stimulation, and a control group of 15 healthy women matched by age. Before beginning ovarian stimulation protocols, all ART candidates should not have a SARS-CoV-2 positive test.
This SARS-CoV-2 infection should also be checked on the day of the final oocyte maturation triggering prior to oocyte retrieval. Some patients have a positive SARS-CoV-2 test when ovary is stimulated and in the day of oocyte retrieval. Treatment is discontinued for these patients due to potential risks for embryo. Oocyte retrieval is advised, although, in a small number of individuals who are susceptible to developing severe ovarian hyperstimulation syndrome (OHSS) and its associated consequences, as well as in certain unique situations like endometriosis or breast cancer patients. Based on the current recommendations in infertility treatment centers, the collected follicular fluid and the cells from the OHSS patients are discarded [9]. For this investigation, unused samples from women infected with COVID-19 were gathered. Prior to the trial, patients were informed verbally and in writing, given enough time to think over their involvement, and provided their written consent. The Shiraz University of Medical Sciences Scientific and Ethics Committee gave its clearance to this study (approval code: IR.SUMS.REC.1400.672).
Stimulation and Granulosa Cells Collection
For every patient, a conventional GnRH antagonist protocol was followed. Following a transvaginal ultrasound (TVU) scan on the second day of the menstrual cycle, ovarian stimulation was initiated with HMG (PDHoMoG®, Pooyesh Daroo, Tehran, Iran) and Follitropin Alpha (Cinal-F®, CinnaGene, Alborz, Iran). On cycle day six, or when the leading follicle reached 12 mm, the GnRH antagonist (Cetrotide®, Injection, Powder, 250 μg, Serpero pharmaceutical, Italy) was started at a dose of 250 μg per day.
The drugs were kept up until the diameter of at least two follicles measured 17-18 mm. Subcutaneous injection of 2-3 ampules of Varian Pharmed’s Variopeptyl®, Injection, 0.1 mg, Tehran, Iran, a GnRH agonist, was then administered to initiate the last stage of oocyte maturation. Oocyte retrieval guided by TVU was carried out approximately 34-36 hours following triggering. Every patient’s excited follicle was recovered. Following a routine isolation process, the acquired follicular fluid was sent to the embryology lab in order to extract granulosa cells.
RNA Isolation and Gene Expression Analysis
Granulosa cells were processed immediately after collection to maintain RNA integrity. A commercially available RNA extraction kit was used for isolation. The quality and quantity of the extracted RNA were evaluated using spectrophotometry and gel electrophoresis. Only samples with high purity (A260/A280 ratio>1.8) and intact RNA profiles were chosen for further analysis. The isolated RNA was then converted into complementary DNA (cDNA) using a reverse transcription kit. The analysis concentrated on several key genes related to inflammation (IL-1B, TNF-α, IL-6, and IL-8), vascular function (VEGF and ANGPT1), cell cycle regulation (FOXL2, Cyclin D2, Cyclin D1, and KLF4), and gonadotropin receptor signaling (luteinizing hormone receptor (LHCGR) and follicle-stimulating hormone receptor (FSHR)).
After conducting qRT-PCR, the fold changes in gene expression between the study group (women with COVID-19 positive test) and the control group (healthy individuals) were determined using the 2-ΔΔCt method.
The data were normalized against the expression of the housekeeping gene GAPDH.
Statistical Analysis
Data were analyzed using SPSS (version 24, IBM, Chicago, IL) and GraphPad Prism (version 8). Differences in baseline characteristics between the study and control groups were evaluated with an independent samples t-test for continuous variables and a chi-square test (or Fisher’s exact test when appropriate) for categorical variables, with a p-value of less than 0.05 considered statistically significant.
Gene expression differences between the COVID-19 positive group and the control group were compared using the Mann-Whitney U test for non-normally distributed data or the Student’s t-test for normally distributed data.
The relationship between comorbidities and gene expression levels was examined using Pearson’s or Spearman’s correlation coefficients, depending on the distribution of the data.
Results
**
- Study Participants
The study group consisted of 15 women who had tested positive for COVID-19 after ovarian stimulation, with ages ranging from 26 to 38 years and a mean age of 31.2 years. The control group comprised 15 healthy women matched by age who were undergoing ART between 27 to 36 years old with the mean age of 32.1. There was no statistically difference between groups regarding the mean age.
- Gene Expression in Granulosa Cells
2.1. Inflammatory Cytokines
The mRNA levels of inflammatory cytokines IL-1β, TNF-α, and IL-6 were significantly elevated in the granulosa cells of women with COVID-19 compared to the control group. Specifically, IL-1β expression was 1.4 times higher (P<0.0001), TNF-α was 2.2 times higher (P<0.0001), and IL-6 was 1.6 times higher (P<0.01). Although IL-8 expression also increased, this change did not achieve statistical significance (Figure-1)
2.2. Vascular Genes
Both the VEGF and ANGPT1 genes were found to be overexpressed in the granulosa cells of the study group. Specifically, VEGF mRNA levels were 1.6 times higher (P<0.0001), while ANGPT1 levels were 1.2 times higher (P=0.001) compared to the control group (Figure-2).
2.3. Cell Cycle
The regulators FOXL2, a vital transcription factor for granulosa cell differentiation, exhibited a downregulation in women positive for COVID-19 (P<0.0001). In contrast, the cell cycle promoters Cyclin D2 and Cyclin D1 were upregulated by 1.4-fold (P=0.0001) and 1.4-fold (P=0.001), respectively. No significant change was observed in KLF4 expression (Figure-3).
2.4. Gonadotropin Receptor
The expression of both LH (LHCGR) and FSH (FSHR) receptor genes in granulosa cells was similar between the study and control groups, showing no significant differences (Figure-4).
Discussion
**
**
The findings of this study reveal significant alterations in the molecular profiles of granulosa cells obtained from women who tested positive for COVID-19 compared to healthy controls. Notably, there were significantly increased mRNA levels of inflammatory cytokines IL-1β, TNF-α, and IL-6 in the granulosa cells of women infected with SARS-CoV-2 undergoing ART. This aligns with earlier findings that COVID-19 infection triggers a strong inflammatory response marked by the release of cytokines [19]. Inflammatory cytokines such as IL-1β and TNF-α are known to disrupt ovarian function through various mechanisms, including inducing apoptosis in granulosa cells, interfering with steroidogenesis, and hindering oocyte maturation [20]. The increased levels of cytokines observed in this study suggest a pro-inflammatory ovarian microenvironment in women with COVID-19, which may negatively affect folliculogenesis and oocyte quality. Interestingly, although IL-8 exhibited a trend toward upregulation, it did not achieve statistical significance, indicating a differential regulation of cytokines in response to SARS-CoV-2 infection. Additionally, the vascular genes VEGF and ANGPT1 were found to be overexpressed in the granulosa cells of women who tested positive for COVID-19. VEGF plays a crucial role in promoting angiogenesis during follicle development by facilitating the formation of new blood vessels [21]. Increased levels of VEGF indicate heightened vascular permeability and angiogenesis, which may disrupt the blood-follicular barrier. The overexpression of ANGPT1, a crucial mediator of vascular stabilization, protection, and remodeling, indicates that significant vascular changes are occurring in response to SARS-CoV-2 infection. Dysregulated angiogenesis and vascular instability are known to negatively impact oocyte maturation [22][23]. Furthermore, the study found that FOXL2, a master regulator of granulosa cell differentiation, was downregulated, while cell cycle genes Cyclin D1 and D2 were upregulated in the granulosa cells of women infected with COVID-19. FOXL2 is essential for maintaining granulosa cell identity and coordinating folliculogenesis with proliferation signals [24][25]. Aberrant expression of FOXL2, coupled with excessive cell cycling, may result in failed differentiation and inadequate coordination of growth with developmental signals, ultimately hindering follicle development. This study is the first to report changes in key cell cycle mediators in relation to COVID-19 infection, offering new mechanistic insights. Notably, there were no differences in the gonadotropin receptor genes LH and FSH between the groups, suggesting that SARS-CoV-2 infection may not directly affect pituitary-ovarian communication. However, the disproportionate changes observed in other molecular regulators underscore the complex interactions among the inflammatory microenvironment, intra-ovarian signaling networks, and oocyte quality in the context of COVID-19.
A limitation of this study was its cross-sectional design, which prevents the assessment of long-term or recurring effects of COVID-19 on ovarian function and fertility potential over time. However, the current data provide new insights into how SARS-CoV-2 infection disrupts various ovarian pathways at the molecular level during the acute phase. By integrating clinical parameters of participants with gene expression patterns, a comprehensive profile analysis was achieved. Future prospective studies are needed to evaluate ovarian reserve markers and the number and quality of oocytes following infection.
Conclusion
In conclusion, this research significantly enhances our understanding of the relationship between COVID-19 and female reproductive health. It reveals that SARS-CoV-2 dysregulates expression of inflammatory cytokines, growth factors, and cell cycle regulators persists in the granulosa cells, indicating long-term pathological changes. This molecular evidence supports the hypothesis that COVID-19 infection may disrupt follicular development and oocyte maturation through altered signaling within the ovary. The findings shed light on potential pathogenic mechanisms and have implications for counseling and managing fertility in women who have recovered from SARS-CoV-2. Further exploration of mitochondrial function, DNA repair capacity, meiotic resumption, and embryo development post-fertilization could provide deeper insights. Additionally, this study underscores the necessity to investigate therapeutic interventions, such as anti-inflammatory strategies, aimed at reversing COVID-19-related ovarian changes and optimizing fertility preservation in women of reproductive age who have been infected.
Conflict of Interest
None.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Phelan N Behan LA Owens L The impact of the COVID-19 pandemic on women’s reproductive health Front Endocrinol 20211264275564275510.3389/fendo.2021.642755 PMC 803058433841334 · doi ↗ · pubmed ↗
- 2Jamali E Shapoori S Farrokhi MR Vakili S Rostamzadeh D Iravanpour F Effect of Disease-Modifying Therapies on COVID-19 Vaccination Efficacy in Multiple Sclerosis Patients: A Comprehensive Review Viral Immunol 20233663687710.1089/vim.2023.003537276047 · doi ↗ · pubmed ↗
- 3Vakili S Roshanisefat S Ghahramani L Jamalnia SA Report of an Iranian COVID-19 Case in a Laparoscopic Cholecystectomy Patient: A Case Report and Insights Journal of Health Sciences & Surveillance System 2021921359
- 4Vakili S Akbari H Jamalnia S Clinical and Laboratory findings on the differences between h 1n 1 influenza and coronavirus disease-2019 (covid-19): focusing on the treatment approach Clin Pulm Med 20202748793
- 5Wu M Ma L Xue L Zhu Q Zhou S Dai J Co-expression of the SARS-Co V-2 entry molecules ACE 2 and TMPRSS 2 in human ovaries: identification of cell types and trends with age Genomics 2021113634496010.1016/j.ygeno.2021.08.012PMC 837246434418496 · doi ↗ · pubmed ↗
- 6Li M-Y Li L Zhang Y Wang X-S Expression of the SARS-Co V-2 cell receptor gene ACE 2 in a wide variety of human tissues Infect Dis Poverty 202090223910.1186/s 40249-020-00662-x PMC 718653432345362 · doi ↗ · pubmed ↗
- 7Goad J Rudolph J Rajkovic A Female reproductive tract has low concentration of SARS-Co V 2 receptors Plos one 20201512 e 0243959 e 024395910.1371/journal.pone.0243959 PMC 773559333315943 · doi ↗ · pubmed ↗
- 8D’Ippolito S Turchiano F Vitagliano A Scutiero G Lanzone A Scambia G Greco P Is there a role for SARS-Co V-2/COVID-19 on the female reproductive system Front physiol 2022138451568451563530905510.3389/fphys.2022.845156 PMC 8924447 · doi ↗ · pubmed ↗