A review of the intertidal Medon Stephens (Coleoptera, Staphylinidae, Paederinae) with description of a new species on the East Asian coasts

Abstract
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
 Figure 1
Figure 1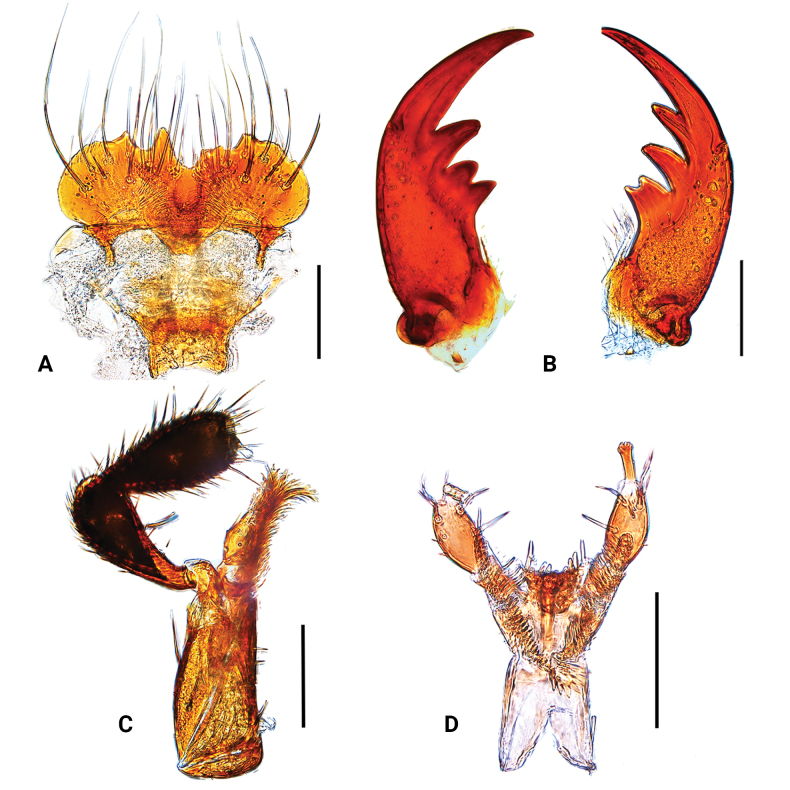 Figure 2
Figure 2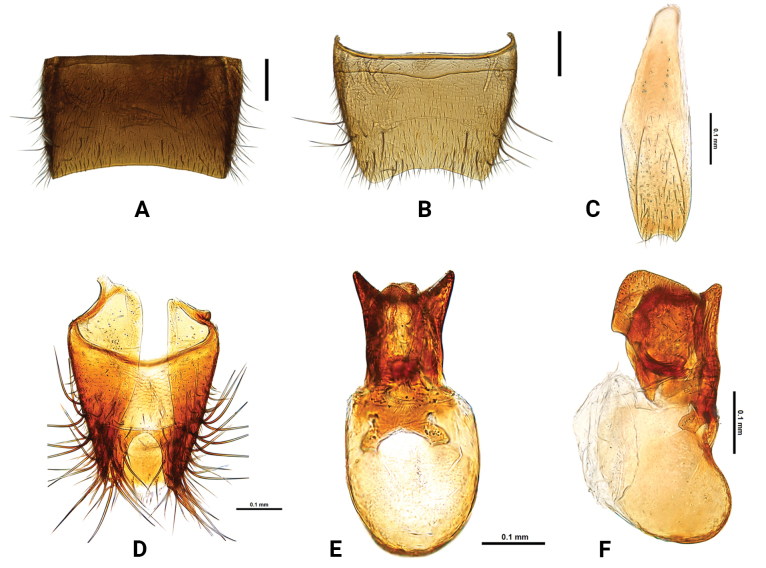 Figure 3
Figure 3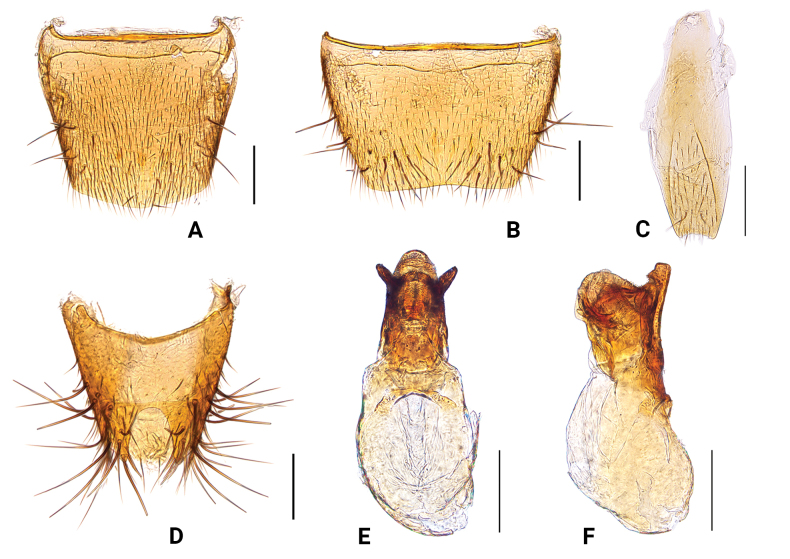 Figure 4
Figure 4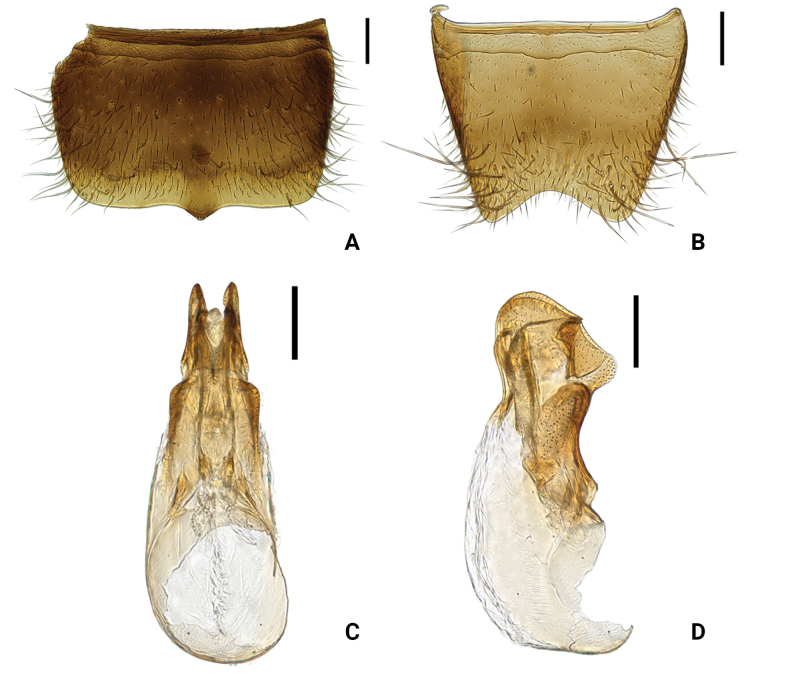 Figure 5
Figure 5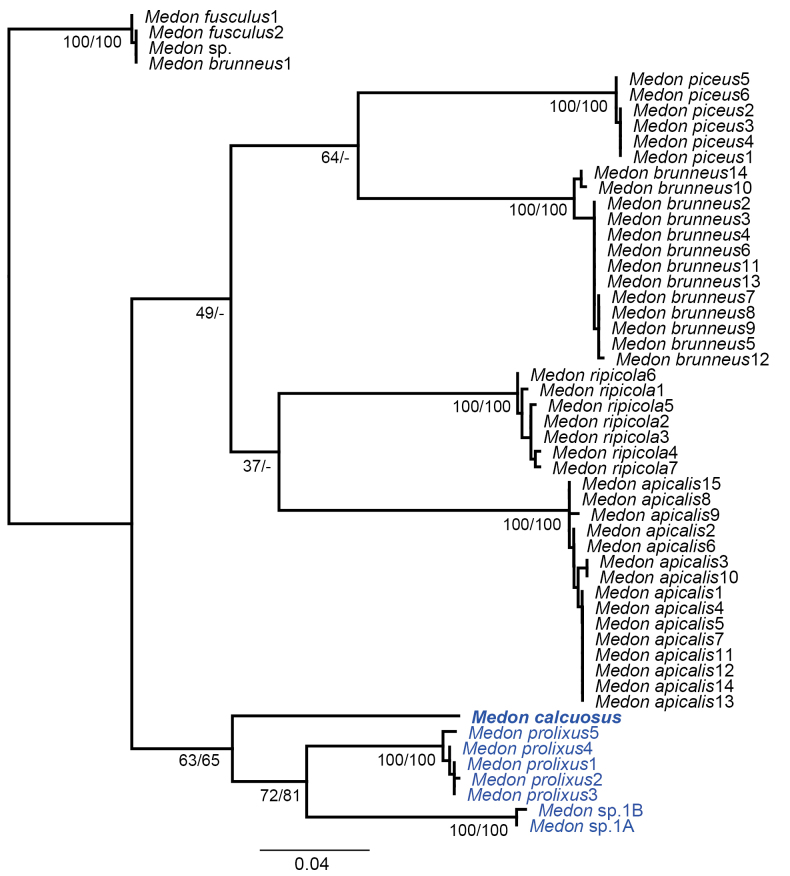 Figure 6
Figure 6| 1 | Body length more than 3.8 mm; body black with brown elytra; black tentorial spots absent on the vertex; antennomeres 8–10 subglobular, about 1.1 times as long as wide; hind wings fully developed and long, reaching to the end of abdomen; apical margin of male sternite VII protruded (Fig. |
|
| – | Body length less than 3.7 mm; body brown with black abdomen (Fig. |
|
| 2 | Apical margin of male sternite VIII subtruncate (Fig. |
|
| – | Apical margin of male sternite VIII slightly emarginate and with small sinuation at middle (Fig. |
|
|
|
| ||
|---|---|---|---|
| Body length | 3.0–3.3 mm | 2.8–3.5 mm | 4.0–4.3 mm |
| Body color | brown to reddish-brown | brown to reddish-brown | black with brown elytra |
| Antenna length | very long, extending to the middle of elytron | very long, reaching to the middle of elytron | long, reaching to just behind the pronotum |
| Antennomeres | all elongate | all elongate | not all elongate, 8–10 subquadrate |
| Eye size | very small, 0.26 times as long as temple | very small, 0.28 times as long as temple | small, 0.4 times as long as temple |
| Black tentorial spots on vertex | present | present | absent |
| Pronotum | slightly narrowed posteriorly | slightly narrowed posteriorly | parallel-sided |
| Hind wings | short, less than 2.0 times as long as elytron | short, less than 2.0 times as long as elytron | long, more than 2.0 times as long as elytron |
| Male sternite VII | subtruncate (Fig. | subtruncate | protruded posteriorly (Fig. |
| Male sternite VIII | slightly emarginate (Fig. | subtruncate posteriorly (Fig. | deeply emarginate (Fig. |
| Median lobe in dorsal aspect | broader (Fig. | narrower (Fig. | narrower (Fig. |
| Median lobe in lateral aspect | apical process shorter (Fig. | apical process longer (Fig. | apical process shorter (Fig. |
| Microhabitats | pebble/gravel beach | pebble/gravel beach | sandy beach |
| Species | Collection locality |
|
|---|---|---|
| KOREA: Gyeongbuk, Gyeongju-si |
| |
| KOREA: Gyeongbuk, Uljin-gun |
| |
| KOREA: Gyeongbuk, Uljin-gun |
| |
| KOREA: Gyeongnam, Geoje-si |
| |
| KOREA: Gyeongnam, Tongyeong-si |
| |
| KOREA: Jeju Prov., Jeju-si |
| |
| KOREA: Gangwon, Sokcho-si |
| |
| KOREA: Jeonnam, Goheung-gun |
|
|
| |||
|---|---|---|---|
| 0 | – | – | |
|
| 9.795–10.111 | 0–0.948 | – |
| 11.374–11.690 | 7.741–8.215 | 0.316 |
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsColeoptera Taxonomy and Distribution · Invertebrate Taxonomy and Ecology · Insect and Arachnid Ecology and Behavior
Introduction
The genus Medon Stephens contains four coastal species along the eastern Pacific, Indian, and northern Atlantic Oceans including the Mediterranean Sea. These species inhabit decaying seaweed on sandy or pebble/gravel beaches (Frank and Ahn 2011; Kim et al. 2011; Shibata and Fujimoto 2017).
As part of an ongoing taxonomic revision of Korean coastal Staphylinidae, we collected several specimens belonging to Medon using the floatation method on pebble/gravel beaches. They are very similar to M.tomokoae Shibata & Fujimoto from Japan, although there are differences in the external form and internal structure of the male genitalia. This led us to investigate the species delimitation of these Medon species in more detail using molecular criteria based on genetic divergence and gene tree monophyly based on COI sequences.
In this paper, we compare morphological and molecular characters among individuals of the East Asian coastal Medon species and describe M.calculosus Ahn & Jeon, sp. nov. with illustrations of diagnostic characters.
Material and methods
The depositories of the specimens examined are as follows: Chungnam National University Insect Collection (CNUIC), Daejeon, Korea, and National Museum of Nature and Science (NMNS), Tsukuba, Japan.
Habitus photographs were made using a Canon EOS 5Ds with an attached Canon MP-E65 mm f/2.8 1–5x macro lens. The produced multilayered images were merged using software (Helicon Focus 7). Microphotographs of mouthparts, male genitalia, and abdominal segments were prepared using a Leica MC170 HD microscope camera mounted on an Olympus BX51 compound microscope. All photographs were finally edited in Adobe Photoshop CS4 and CC.
A total of eight new sequences were generated from the Korean specimens (633 bp of the partial COI gene region). All the new sequences were deposited in GenBank (accession numbers: PP578207– PP578214 in Table 2). For comparison, 45 partial COI (624 bp) sequences were downloaded from GenBank for five Medon species (accession numbers: MW259761, MN249800, KR129040, HQ953660, HQ953398, HQ954439, HQ954438, KR487620, KR491265, KU919263, KU919097, KU916556, KU915842, KU915543, KU913155, KU913130, KU912779, KU912022, KU911503, KU910374, KU909268, KU909128, KU908943, KU908290, KU907897, KM451678, KM449956, KM449805, KM449734, KM449603, KM448496, KM447995, KM447725, KM447712, KM447270, KM446843, KM446508, KM445710, KM445254, KM444731, KM444711, KM441191, KM441020, KM440342, and KJ962140). DNA extraction, sequencing, and alignment were performed with reference to Song et al. (2019). Primers and amplification strategies are detailed in Yoo et al. (2021).
Intra- and interspecific distances were calculated using the uncorrected pairwise distance method in MEGA 7.0 (Kumar et al. 2016). Parsimony (PA) and maximum likelihood (ML) analyses were conducted using PAUP* 4.0 (Swofford 2002) and PhyML 3.3 (Guindon et al. 2010) with default options implemented in Geneious (ver. 2025.0.2). Clade support values were evaluated using 100 bootstrap replicates.
Taxonomy
Key to species of the East Asian coastal Medon
**: **
Genus Medon Stephens, 1833
Medon
calculosus
sp. nov.
Taxon classificationAnimaliaColeopteraStaphylinidae
41D1757A-D8D9-54BB-9689-16BB9BC005BA
https://zoobank.org/0E15F233-F763-43A4-B30B-4D83C5D46B0A
Type specimens.
Holotype • labeled as follows: “Korea: Gyeongbuk Prov., Pohang-si, Janggi-myeon, Gaewon-ri 445-12 35.855905°N, 129.524668°E, 31 X 2023, KJ Ahn, pebble beach in low-tide zone; Holotype, Medoncalculosus Ahn and Jeon, Desig. K.-J. Ahn 2024, deposited in CNUIC, Daejeon, Korea”. Paratypes • 3 exx., same data as holotype • 2 exx., Korea: Gyeongbuk Prov., Pohang-si, Janggi-myeon, Gaewon-ri 445-12 35.855905°N, 129.524668°E, 2 XI 2023, KJ Ahn, pebble beach in mid-tide zone • 2 exx., same locality as above, 24 III 2023, KJ Ahn, pebble beach in low-tide zone • 2 exx., Gyeongju-si, Yangnam-myeon, Suryeom-ri, Jigyeong-beach, 35.652814°N, 129.450041°E, 2 XI 2023, KJ Ahn, pebble beach in low-tide zone • 2 exx., Naa-ri, Naa-beach, 35°41′57.31″N, 129°28′26.73″E, 7m, 13 VII 2018, IS Yoo, JS Lee, JG Jung, in gravels covered with seaweed in high-supratidal zone, flotation • 2 exx., Yeongduk-gun, Byonggok-myeon, Byonggok-ri, 36.602581°N, 129.415695°E, 24 IX 2022, KJ Ahn, flotation on pebble beach.
Description.
Male. Body length 3.0–3.3 mm. Body slender, more or less parallel-sided, flattened, and densely pubescent. Head, pronotum, elytra, antennae and legs brown to reddish-brown; abdomen black (Fig. 1). Head. Subquadrate, about 1.03 times as long as wide, widest basal 1/5 and about 1.15 times wider than pronotum, dorsal surface covered with dense pubescence; eye small, about 0.26 times as long as temple; antenna very long, extending to middle of elytron; inserted under side of front head, insertion invisible from dorsal view; all antennomeres elongate, scape longest and widest, pedicel to antennomere 7 distinctly longer than wide, antennomeres 8–10 subglobular, antennomere 11 water drop-shaped, relative length of 11 antennomeres 21:14:14:13:14:13:13:11:11:11:14; gular sutures narrowly separated and converged posteriorly. Neck moderately wide, about 1/3 as wide as head.
Male habitus AMedoncalculosus sp. nov. 3.1 mm B paler form of M.calculosus sp. nov. CM.tomokoae, 3.2 mm.
Mouthparts. Labrum (Fig. 2A) broad, anterior margin with two pairs of tooth-like processes, medial one longer and slender; mandibles (Fig. 2B) asymmetrical, with 3–4 internal teeth; maxilla (Fig. 2C) with galea and lacinia fringed with long setae; maxillary palpomere 1 small, about 3.18 times as long as 2; palpomere 2 expanded apically, little curved inwardly, about 0.97 times as long as 3; palpomere 3 droplet-shaped, widest apical third; palpomere 4 minute, about 0.88 times as long as 1; labium (Fig. 2D) with palpomere 1 small, about 0.46 times as long as 2; palpomere 2 widest near middle, about 1.87 times as long as 3; palpomere 3 small and thin, about 1.13 times as long as 1. Thorax. Pronotum more or less rectangular, about 1.17 times as long as wide; narrower than head, almost as long as head, widest apical 1/5 and narrowed posteriorly, yellow short setae densely present, impunctate central region present; prosternum well developed, median area upheaved with transverse carina, prosternal process acute and very long, hypomeral projection well developed and triangular. Elytra about 1.09 times as wide as pronotum, almost as long as pronotum, elytron 2.37 times as long as wide, distinct and shallow punctures present, covered with yellow setae, lateral margin straight and parallel. Metendosternite Y-shaped. Hind wings short but longer than elytron. Tarsal formula 5-5-5, front, middle and hind tarsomeres 1–5 each strongly widened. Abdomen. Almost parallel-sided and broadened posteriorly after segment VI. Tergites III–VI with shallow and transverse basal depression. Sternite III with basal transverse carina, medially pointed; apical margin of tergite VII subtruncate (Fig. 3A) and sternite VIII slightly emarginate (Fig. 3B); apical margin of sternite IX (Fig. 3C) slightly emarginate; tergites IX and X (Fig. 3D) covered with scattered setae, apical margin with numerous long and brown setae. Aedeagus. Median lobe symmetrical, comprising about half sclerotized part and membraneous part; more or less triangular lobe protruded in dorsal view (Fig. 3E); apical process narrow and well sclerotized in lateral view (Fig. 3F).
Medoncalculosus sp. nov., male A labrum, dorsal aspect B mandible, dorsal aspect (left and right) C maxilla, dorsal aspect D labium, ventral aspect. Scale bars: 0.1 mm.
Medoncalculosus sp. nov. A male sternite VII, ventral aspect B male sternite VIII, ventral aspect C male sternite IX, ventral aspect D male tergites IX and X, dorsal aspect E aedeagus, dorsal aspect F aedeagus, lateral aspect. Scale bars: 0.1 mm.
Female. Similar to male, but apical margin of sternite VIII rounded.
Distribution.
Korea (South).
Etymology.
The name is derived from the Latin calculus meaning ‘pebble’, referring to the species’ marine coastal habitat.
Remarks.
This species is extremely similar to M.tomokoae in external morphological characters (Figs 3A–D, 4A–D) but can be distinguished by the different external form and internal structure of the aedeagus (Figs 3E, F, 4E, F) and by the characters listed in Table 1.
MedontomokoaeA male tergite VIII, dorsal aspect B male sternite VIII, ventral aspect C male sternite IX, ventral aspect D male tergites IX and X, dorsal aspect E aedeagus, dorsal aspect F aedeagus, lateral aspect. Scale bars: 0.1 mm.
Discussion
Diagnostic characters among members of the coastal Medon species (M.calculosus, M.prolixus, and M.tomokoae) include the shape of the antennomeres and male sternites VII–VIII, the size of the hind wings, body length, and the shape and structure of the male genitalia (Table 1). The male genitalia characters are especially important for species delimitation, as in other staphylinid beetles. Our morphological study showed that M.calculosus is clearly different from other previously described species.
Furthermore, the pairwise distance data support the validity of the new species: interspecific genetic divergence of the COI (633 bp) using uncorrected p-distance among eight Medon individuals, including one new species, ranged from 7.741% (between M.prolixus and Medon sp.) to 11.690% (Medon sp. and M.calculosus), while intraspecific divergence ranged from 0% to 0.790% (among M.prolixus) (Table 3). Our phylogenetic analyses also support the validity of the new species (Fig. 6). The PA tree showed polytomy in species relationships, but all seven species were supported as a lineage with 100% bootstrap value (not shown). One individual of M.brunneus (M.brunneus 1 – KR487620) and an unidentified specimen (Medon sp. – KR491265) were grouped with M.fusculus (Fig. 6). They need further investigation to confirm their identifications. Fresh specimens of M.tomokoae were not available for DNA analysis.
MedonprolixusA male sternite VII, ventral aspect B male sternite VIII, ventral aspect C aedeagus, dorsal aspect D aedeagus, lateral aspect. Scale bars: 0.1 mm.
Maximum likelihood tree of Medon based on partial COI gene sequence with bootstrap values (left) and parsimony analysis bootstrap values (right).
We found two specimens, one each from Sokcho (central Korea) and Goheung (southern Korea), that showed a genetic difference from all other included species (7.741%). They were also separated from other species in the phylogenetic analyses (Fig. 6). However, they are all females and will remain undescribed until we find a male to confirm whether it could be M.tomokoae or a new species.
Shibata and Fujimoto (2017) were the first to note that M.tomokoae was collected under stones on pebble/gravel beaches. Medoncalculosus is the second species collected from the same microhabitats in association with Halorhadinus Sawada, Myrmecopora Saulcy, Physoplectus Reitter, and Giulianium Moore, among others, in the low to mid-tide zones. In contrast, M.prolixus is readily found under seaweed/debris on sandy beaches in high-tide to splash zones.
Supplementary Material
XML Treatment for Medon calculosus
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Frank JH Ahn KJ (2011) Coastal Staphylinidae (Coleoptera): A worldwide checklist, biogeography and natural history.Zoo Keys 10: 1–98. 10.3897/zookeys.107.1651 PMC 339218822792029 · doi ↗ · pubmed ↗
- 2Guindon S Dufayar J-F Lefort V Anisimova M Hordiik W Gascuel O (2010) New algorithms and methods to estimate maximum-likelihood phylogenies: assessing the performance of Phy ML 3.0.Systematic Biology 59: 307–321. 10.1093/sysbio/syq 01020525638 · doi ↗ · pubmed ↗
- 3Kim TK Cho YB Ahn KJ (2011) Two marine littoral species of the genus Medon (Coleoptera: Staphylinidae: Paederinae) new to Korea.Korean Journal of Systematic Zoology 27: 69–73. 10.5635/KJSZ.2011.27.1.069 · doi ↗
- 4Kumar S Stecher G Tamura K (2016) MEGA 7: molecular evolutionary genetics analysis version 7.0 for bigger dataset.Molecular Biology and Evolution 33: 1870–1874. 10.1093/molbev/msw 05427004904 PMC 8210823 · doi ↗ · pubmed ↗
- 5Shibata Y Fujimoto H (2017) A new intertidal species of the genus Medon (Coleoptera, Staphylinidae, Paederinae) from Kagawa, Japan.Elytra, New Series 7: 55–60.
- 6Song JH Lee G Lee JS Ahn KJ (2019) Independent origins of coastal colonization in the tribe Athetini (Coleoptera, Staphylinidae).Zoologica Scripta 48: 337–348. 10.1111/zsc.12351 · doi ↗
- 7Swofford DL (2002) PAUP* version 4.0b 10. Phylogenetic analysis using parsimony (*and other methods). Sinauer Associates.
- 8Yoo IS Lee JSÔhara M Ahn KJ (2021) Three synonyms of the coastal Phucobius Sharp species (Coleoptera: Staphylinidae) are proposed based on morphological and molecular characters.Journal of Asia-Pacific Entomology 24: 320–328. 10.1016/j.aspen.2020.12.015 · doi ↗