ABA and Pre-Harvest Sprouting Differences in Knockout Lines of OsPHS3 Encoding Carotenoid Isomerase via CRISPR/Cas9 in Rice
Yu-Jin Jung, Jiyun Go, Jin-Young Kim, Hyo-Ju Lee, Jong-Hee Kim, Hye-Mi Lee, Yong-Gu Cho, Kwon-Kyoo Kang

TL;DR
This paper shows how knocking out the OsPHS3 gene in rice affects plant growth and increases pre-harvest sprouting, offering insights into similar crops.
Contribution
The study reveals novel molecular mechanisms of pre-harvest sprouting in rice through OsPHS3 gene knockout and carotenoid pathway analysis.
Findings
OsPHS3 knockout lines showed reduced plant height, panicle length, and seed weight but increased pre-harvest sprouting.
KO lines had higher expression of PSY1, PSY2, and PDS genes but lower ε-LCY, β-LCY, and ZEP1 transcripts.
Carotenoid and ABA levels decreased in knockout lines compared to wild-type rice.
Abstract
We generated and characterized knockout mutant lines of the OsPHS3 gene using the CRISPR/Cas9 system. The knockout lines of the OsPHS3 gene showed that 1 bp and 7 bp deletion, early termination codons were used for protein production. Agronomic characteristics of knock-out lines were reduced in plant height, culm diameter, panicle length, seed size and weight, except for the number of tillers. In addition, we analyzed the expression levels of carotenoid biosynthesis genes by qRT-PCR. Among the genes encoding carotenoid metabolic pathway enzymes, the level of transcripts of PSY1, PSY2, PSY3, PDS and ZDS were higher in the KO lines than in the WT line. In contrast, transcription of the ε-LCY, β-LCY and ZEP1 genes were downregulated in the KO lines compared to the WT line. Also, the KO lines decreased carotenoid content and ABA amount compared to WT, while preharvest sprouts increased.…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
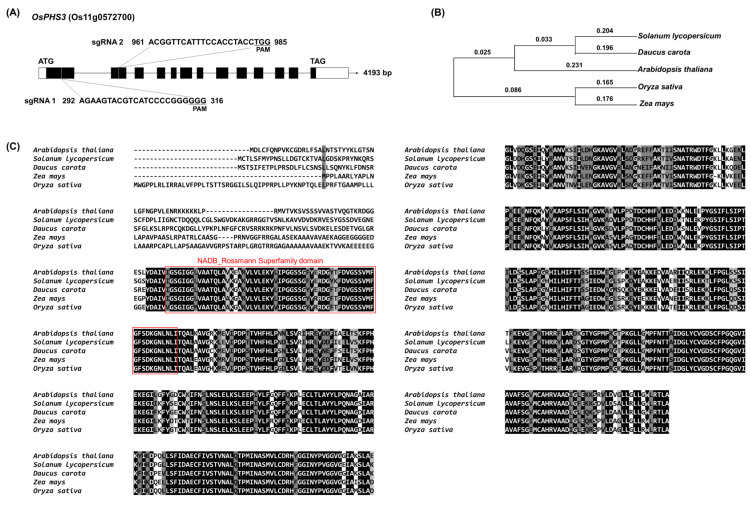 Figure 1
Figure 1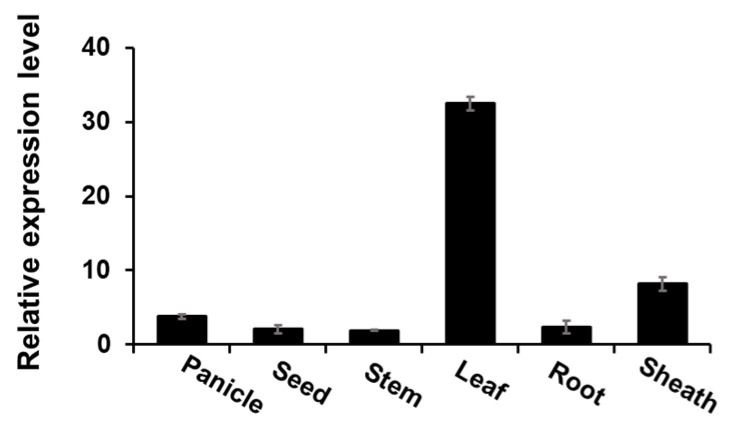 Figure 2
Figure 2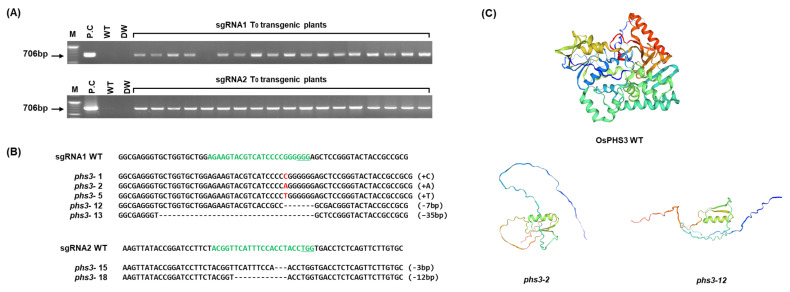 Figure 3
Figure 3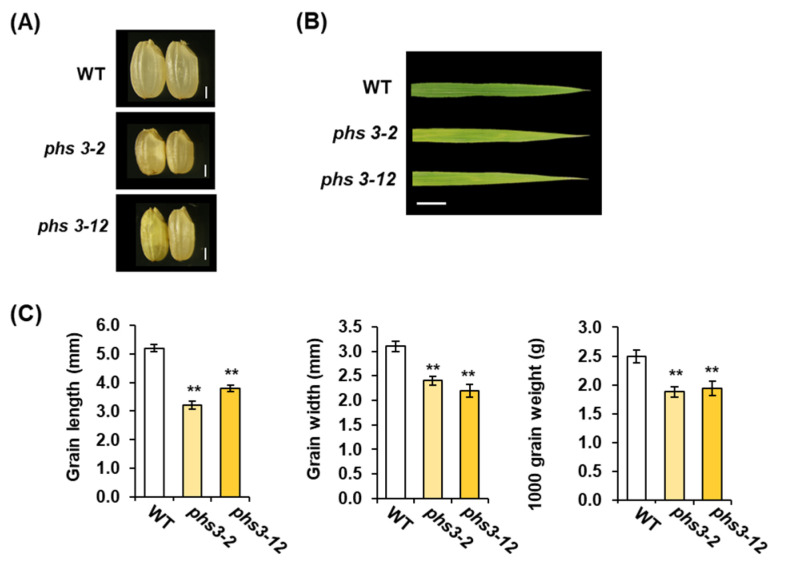 Figure 4
Figure 4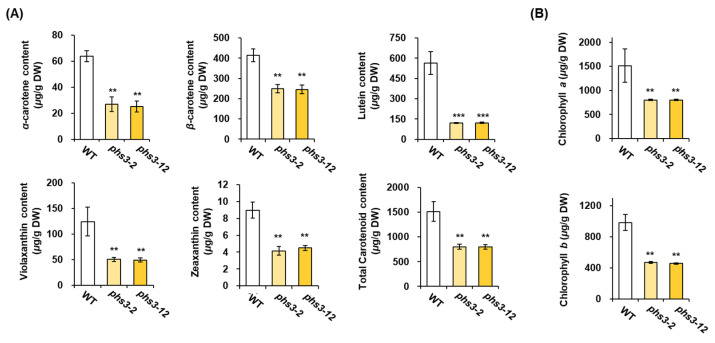 Figure 5
Figure 5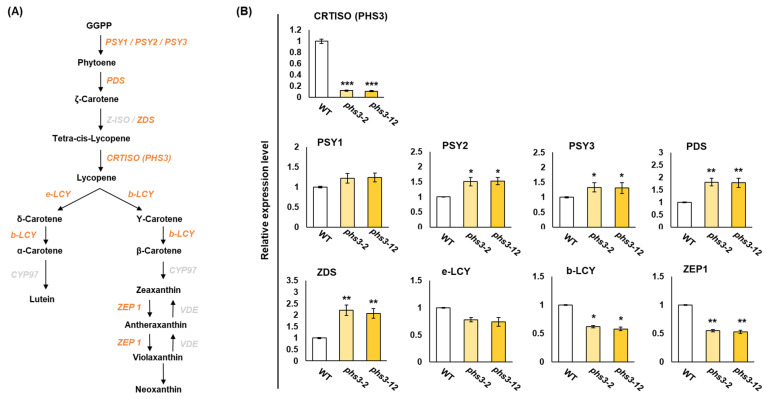 Figure 6
Figure 6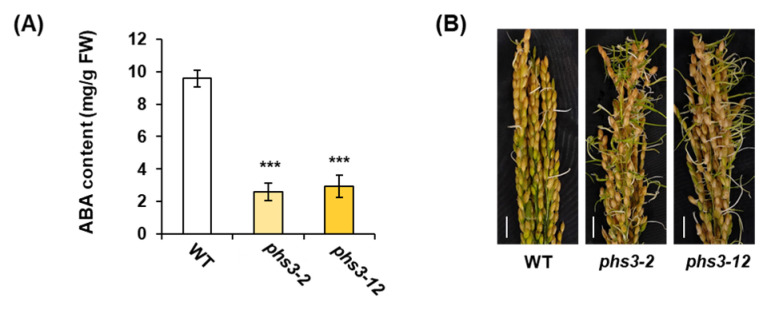 Figure 7
Figure 7- —the New Breeding Technologies Development Program
- —National Research Foundation of Korea (NRF) funded by the Ministry of Education
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsPhotosynthetic Processes and Mechanisms · Antioxidant Activity and Oxidative Stress · Plant Stress Responses and Tolerance
1. Introduction
The most influential factors in rice production are abiotic stresses such as temperature, moisture, and light. Among them, frequent rainfall during the rice harvest season causes pre-harvest sprouting (PHS) phenomena, not only causes reduction in grains yield but also affects the quality of the grain [1,2]. As a mechanism to prevent PHS, seed dormancy is a property that limits seed germination under unfit ecological conditions [3,4]. Previous studies have shown that abscisic acid (ABA) biosynthesis or impaired-reactive mutations (corn and rice viviparous (vp)) and Arabidopsis ABA deficiency (aba), and ABA insensitivity (abi) are known to germinate rapidly under humid conditions [5,6]. At least 10 viviparous mutations have been reported in corn (Zea may L.), most of which (vp2, vp5, vp7, vp9, w3, y9) have blocked the biosynthesis of carotenoid precursors, resulting in poor ABA synthesis [7]. In carotenoid biosynthesis, the phytoene synthetase (PSY) enzyme produces C_40_ carotenoid phytoene in the C_20_ geranylgeranyl diphosphate (GGPP) molecule. Subsequently, the phytoene undergoes four unsaturated reactions with the production of lycopene, followed by a series of steps including cyclization and hydroxylation to produce α-carotene, β-carotene, lutein, and xanthophyll and zeaxanthin [8]. After the cleavage of the C_40_ carotenoid precursor, the intermediate xanthoxin is converted to ABA via ABA aldehyde [9,10]. 15-cis-phytoene obtained by phytoene synthase (PSY) is converted to all-trans lycopene through several steps of unsaturation and isomerization through the action of phytoene desaturase (PDS) and ζ desaturase (ZDS), 15-cis-ζ isomerase (Z-ISO), and carotenoid isomerase (CRTISO) enzymes [11]. Z-ISO has been identified in arabidopsis and corn, and this enzyme catalyzes the isomerization of the PDS product 9,15,9′-tri-cis-ζ to 9,9′-di-cis-carotene [12,13]. Afterwards, ZDS converts 9,9′-di-cis-ζ-carotene to 7,9,7′,9′-tetra-cis-lycopene (prolycopene) through 7,9,9′-tri-cis-neurosporene [11]. CRTISO, a carotenoid isomerase, converts pro-lycopene to all-trans-lycopene. All-trans-lycopene produced through the CRTISO enzyme undergoes beta-lycopene cyclization to form all-trans-α-carotene and all-trans-β-carotene [14,15]. Through a series of reactions, all-trans-α-carotene is finally converted to lutein, and all-trans-β-carotene is converted to zeaxanthin and violaxanthin, during which, intermediates are converted to abscisic acid (ABA) and strigolactones (SL) by apocarotenoid enzymes [16]. CRTISO reported that the ccr2 mutants in tomatoes and tangerines accumulate pro-lycopene and other poly-cis-carotene precursors in plastids of fruits [14,17]. In addition, zebra2 mutants grown in a light-free environment accumulated pro-lycopene, and rice seedlings grown in a light-free environment showed reduced lutein levels and multi-colored phenotypes in leaves [18,19]. In addition, the mit3 mutant showed phenotypes such as dwarfism, variegation in leaves, and an increase in the number of tillers [20]. It was considered that the increase in the number of tillers was caused by a decrease in the content of strigolactone which acts as the tiller inhibition function derived from the carotenoid pathway [20]. Recently, known gene editing technology using gene scissors has shown the potential to induce gene mutations at desired genomic DNA locations by using various types of site-specific nuclease. CRISPR/Cas9 (clustered regularly interposed short palindromic repeat (CRISPR)-associated protein (Cas) system), third-generation gene scissors, is a tool that can easily and accurately edit a target gene region [21]. Therefore, in this paper, we edited the PHS3 (Os11g0572700) gene-encoding carotenoid isomerase of ABA biosynthesis using the CRISPR/Cas9 system. Our results suggested that the knockout lines of the PHS3 gene would certainly help to not only reduction in ABA content but also PHS in rice, which will be helpful for elucidating the molecular mechanisms of PHS in other crops such as wheat and barley, which are susceptible to PHS.
2. Results
2.1. Confirmation and Expression of PHS3 Gene Encoding Carotenoid Isomerase in Rice
In rice, the PHS3 gene is located on chromosome 11, the genomic sequence length is 4193 bp, consisting of 13 exons and 12 introns, and the CDS length is 1925 bp, encoding 641 amino acids (Figure 1). Also, genetic similarity between the PHS3 gene sequences of various plant species showed high homology to OsPHS3 and Zea mays (NM_001367269) and showed 87.83% identity at the amino acid sequence level. In addition, Arabidopsis thaliana (NM_100559), Solanum lycopersicum (NM_001309230), and Daucus carota (NM_001329170) showed 74.55–74.13% identity (Figure 1B). The conserved domain of the OsPHS3 protein has the NADB_Rossmann Superfamily containing amino oxidase, carotenoid isomerase, numerous dehydrogenases in metabolic pathways such as glycolysis, and GXGXGG patterns found in many oxidoreductases (Figure 1C). The expression level of the OsPHS3 gene was analyzed in each organ using qRT PCR. As a result, the OsPHS3 gene was expressed at a very high level in the leaves (Figure 2).
2.2. Generation of OsPHS3 Knockout Homozygous Mutants
To investigate the in vivo function and role of the OsPHS3 encoding carotenoid isomerase in rice, thirty-four transgenic plants were obtained by editing the first and second exons of OsPHS3 in Dongjin as a target using the CRISPR/Cas9 system (Figure 1A and Figure 3A, Supplementary Figure S1). As a result of confirming the variation type of the target site through deep sequencing in the transformation-confirmed individuals, 18 out of 34 plants had targeted mutations, showing 53% gene editing efficiency, and the ratio of homozygous allelic 55.6%, heterozygous allelic 38.9%, and bi-allelic 5.6% (Supplementary Table S1). Of these, the selected phs3-2 and phs3-12 mutations were a 1 bp (A) insertion and a 7 bp deletion (GGGGGGA), respectively (Figure 3B). The DNA sequences of OsPHS3 WT and two mutant lines were translated into amino acid sequences, and the protein structures were predicted. The results showed that 1 bp insertion and 7 bp deletion in the OsPHS3 gene caused an early stop codon, which made complete protein production impossible. However, the NADB_Rossmann Superfamily domain was well conserved in amino acid sequence and protein structure (Figure 3C, Supplemental Figure S2). Therefore, the phs3-2 and phs3-12 mutants harvested T_1_ seeds by self-fertilization. In addition, we harvested T_2_ seeds from plants without amplified bands by PCR analysis using forward primers and reverse primers present in the bar gene region to select plants with T-DNA removed (Supplemental Figure S3). Harvested seeds were named phs3-2 and phs3-12 null lines, respectively, and were used in the following analyses.
2.3. Agronomic Traits of phs3-2 and phs3-12 Null Lines
To observe the growth of phs3-2 and phs3-12 null lines in a rice field, major agricultural traits such as plant height, culm diameter, panicle length, and number of tillers were investigated. The phs3-2 and phs3-12 null lines showed decreased plant height, stem diameter, and spike length compared to WT, but increased tiller numbers compared to WT (Table 1). In addition, the seed size and seed weight of the phs3-2 and phs3-12 null lines decreased compared to WT (Figure 4C). The phs3-2 and phs3-12 null lines showed a distinctly pale yellow-green phenotype in the leaf (Figure 4B).
2.4. Carotenoid Profiles of phs3-2 and phs3-12 Null Lines
To analyze the carotenoid metabolisms of phs3-2 and phs3-12 null lines, HPLC analysis was performed with extracts from null lines and WT grown under light conditions. Phytoene, zeta-carotene, and pro-lycopene were accumulated in the leaves of the phs3-2 and phs3-12 null lines (Figure 5). Also, the α-carotene, β-carotene, lutein, violaxanthin, and zeaxanthin, which are carotenoid substances after carotenoid isomerase converts pro-lycopene into all-trans lycopene to form a cyclization substrate, were measured and compared (Figure 5). In all the items examined, phs3-2 and phs3-12 null lines were reduced by 2 to 4 times than those of WT.
2.5. Expression Profiling of Carotenoid Biosynthesis Genes
To investigate the expression of genes placed on the circuit due to OsPHS3 gene knockout in the carotenoid biosynthesis pathway, we performed qRT-PCR analysis on a total of nine genes including PSY1, PSY2, PSY3, PDS, ZDS, CRTISO, ε*-LCY*, β -LCY and ZEP1. As a result, gene expressions involved in PSY1, PSY2, PSY3, PDS, and ZDS in the phs3-2 and phs3-12 null lines tended to be higher than those in WT. However, in the rest of the genes, the phs3-2 and phs3-12 null lines showed lower expression levels compared to WT (Figure 6).
2.6. ABA and Pre-Harvest Sprouting in phs3-2 and phs3-12 Null Lines
To investigate whether the reduced level of carotenoids in phs3-2 and phs3-12 null lines causes a decreased level of ABA, we measured the ABA content by immunoassay. The amount of ABA in phs3-2 and phs3-12 null lines was reduced compared to the wild type. In addition, in pre-harvest sprouting experiments, the germination rates of phs3-2 and phs3-12 null lines were significantly increased compared to WT (Figure 7).
3. Discussion
PHS is a phenomenon in which rice, wheat, and barley sprout from ears before harvesting due to frequent rainfall and loading in the early ripening season [3,22]. These phenomena refer to the breakdown and dissolution of germination inhibitory substances in ears exposed to excessive humidity conditions, breaking the seed’s dormancy and progressing germination [23]. Additionally, factors closely related to PHS include external factors such as environmental stress, cultivation conditions, harvest time, day length, and temperature during ripening, as well as internal factors such as seed dormancy, seed permeability, amylase activity, and endogenous hormone level [24,25]. Among them, seed dormancy occurs through interactions with plant hormones such as ABA and external factors such as field conditions or temperature during the maturation of seeds [26]. Thus, embryonic ABA not only plays a central role in inducing and maintaining seed dormancy, but also has mechanisms that inhibit the transition from embryo to germination. The CRTISO protein in the carotenoid biosynthesis pathway isomerizes pro-lycopene to convert it into all-trans lycopene, through which precursors such as α-carotene, β carotene, lutein, and zeaxanthin are synthesized [15]. In several plants, such as Arabidopsis thaliana, Zea mays, Solanum lycopersicum, and Daucus carota, CRTISO functions as carotenoid isomerase, amino oxidase, and has NADB_Rossmann Superfamily domains during carotenoid biosynthesis [15]. In this study, we used the CRISPR/Cas9 system to knockout of the PHS3 gene encoding carotenoid isomerase in Dongjin rice and showed non-lethal and ’variegated’ phenotypes (Figure 4 and Figure 7). In the leaves grown on phs3-2 and phs3-12 null lines, α-carotene, β carotene, lutein, and zeaxanthin were dramatically reduced, and pro-lycopene accumulated (Figure 5). These results were previously consistent with those of crtiso variants of tomato and Arabidopsis. Interestingly, CRTISO activity could explain the survival of phs3 mutants by photoisomerization [27,28]. Usually, membranes in the form of chlorophyll–carotenoid–protein complexes and some carotenoid enzymes are associated with membranes, and changes in carotenoid composition or carotenoid enzymes themselves always lead to abnormal pigmentation [8,14,29]. In addition, CRTISO enzyme activity was lost in the ccr2 Arabidopsis mutant, showing a variegated phenotype [14]. Furthermore, qRT-PCR analysis of nine genes presents in the carotenoid biosynthetic pathway in the present study using phs3-2 and phs3-12 null lines showed that the expression level of genes producing PSY1, PSY2, PSY3, PDS, and ZDS was high, but the expression of all genes belonging to the next stage decreased. These results were identical to the changes in carotenoid metabolites (Figure 6). In addition, in this study, we investigated how the reduction in carotenoids affects ABA content and pre-harvest sprouting. As a result, the ABA content in phs3-2 and phs3-12 null lines was lower than that of WT, and it was sensitive to pre-harvest sprouting (Figure 7). Previous studies have suggested that ABA-insensitive arabidopsis mutants have reduced seed maturity and dormant status. However, ABA-deficient mutations in corn suggest that ABA absolute levels do not always correlate perfectly with dormancy and germination, but that other regulatory factors are involved together [30]. Therefore, viviparous mutants in rice are ideal for elucidating the complex mechanisms of pre-harvest sprouting and dormancy. Therefore, a detailed comparison of the differential expression patterns between the different kinds of osphs3 mutants and WT involved in the carotenoid biosynthesis pathway is thought to be able to identify the major factors regulating PHS. Therefore, the KO lines obtained in this study are thought to be able to assess whether ABA metabolism is highly regulated at the temporal and spatial levels during seed development. In addition, we look forward to more information about the physiological function of carotenoids and the molecular mechanisms of PHS in the future.
4. Materials and Methods
4.1. Plant Materials and Growing Conditions
Dongjin (Oryza sativa L., ssp. Japonica) used in the study was used for breeding transgenic plants. Transgenic plants were grown in GMO-free greenhouses after acclimation [31]. After harvesting the seeds, the moisture was dried to 14% and then stored at a temperature of 4 °C.
4.2. Selection of Gene Target Site and CRISPR/Cas9 Vector Construction
The PHS3 gene (accession no. BAF28490), which synthesizes all-trans-lycopene in the rice carotenoid biosynthesis pathway, has genomic information (Gene: Os11g0572700) from Gramene site (https://www.gramene.org/, accessed on 6 January 2020 [32]. The sgRNAs were selected using CRISPR RGEN Tools (http://www.rgenome.net/cas-designer/, accessed on 23 January 2020) with the PHS3 gene information [32,33]. The CRISPR/Cas9 vectors were constructed to express sgRNA such as OsU3:: OsPHS3-sgRNA/pBOsC according to the previously reported method [33], and the flanking sequence was confirmed by the Sanger sequencing method [34].
4.3. Generation and Selection of Transgenic Rice Plants
Rice transformation was performed according to the method of Nishimura et al. (2006) [35]. Rice seed-derived callus tissues were infected using Agrobacterium tumefaciens strain EHA105. Prior to the transplantation of the transgenic plants into the soil, a selection medium containing 6 mg/liter phosphinotrisin and 400 mg/liter carbenicillin was used. Total DNA extraction was performed by sampling 100 mg of leaves and using a DNA Quick Plant Kit (Inclone, Jeonju, Republic of Korea). Selection of T-DNA-inserted transgenic plants was performed using bar gene-specific primers (Bar-F 5′−CGTCAACCACTACATCGAGA−3′) and (Bar-R 5′−AAGTCCAGCTGCCAGAAA−3′), under PCR conditions in which pre-denaturation was performed at 95 °C for 5 min, denaturation was performed at 95 °C for 30 s, extension was repeated at 56 °C for 30 s, and extension was performed at 72 °C for 7 min. Selection of T-DNA-inserted transgenic plants was carried out using bar gene-specific primers (Bar-F 5′−CGTCAACCACTACATCGAGA−3′) and (Bar-R 5′−AAGTCCAGCTGCCAGAAA−3′), and PCR conditions were carried out according to previously reported methods [35].
4.4. Deep Sequencing Analysis of Target Genes
PCR analysis was performed using a PCR master mix (Bioneer, Daejeon, Republic of Korea) with genomic DNA (50 ng/μL) as a template to identify mutations at the target site. PCR amplicons were performed using MiniSeq (Illumina, San Diego, CA, USA) paired-end read sequencing, and next-generation sequencing (NGS) data derived from MiniSeq were performed using Cas-Analyzer (https://www.rgenome.net/cas-analyzer, accessed on 16 July 2024), which was previously reported by Park et al., 2017 [36].
4.5. Carotenoid Extraction and HPLC Analysis
Carotenoid extraction from WT and transgenic rice leaves was performed by extracting 100 mg lyophilized with acetone (0.01% BHT). The extract was centrifuged at 5640× g (Eppendorf, 5430R, Hamburg, Germany) for 30 min at 4 °C conditions and filtered through a 0.45 μm PTFE syringe filter (Whatman, New York, NY, USA). An Agilent 1260 high-performance liquid chromatography (HPLC) system (Hewlett-Packard, Waldbronn, Germany) was used according to the previously reported method [37]. Thereafter, Chemstation software (Agilent, Santa Clara, CA, USA) was used for the collection and analysis of HPLC. ESI-MS spectra were obtained using an API 4000TM Triple Quadrupole Mass Spectrometer (SCIEX, Concord, ON, Canada). The source temperature was 300 °C, and the ion spray voltage was performed at 5500 V. Gas 1 and 2 settings for nitrogen (>99.999%) were set to 60 and 70, respectively.
4.6. Gene Expression Profiling by qRT-PCR Analysis
Total RNA was extracted by sampling 100 mg of 2-week-old plant leaves using a plant total RNA mini-kit (Cat. FAPRK 001-1). Extracted total RNA was synthesized with cDNA using a reverse transcription system (Takara, www.takara-bio.com). qRT-PCR analysis was performed under conditions previously reported by a Light Cycler 480 device (Roche, Pleasanton, CA, USA) with a total volume of 20 μL using cDNA, primers, and SYBR Green Real-time PCR Master Mix (Toyobo, https://www.bio-toyobo.cn) at a concentration of 50 ng/μL [31]. The housekeeping gene OsActin (Os11g0163100) was used to quantify expression levels. Relative quantification of transcript levels was performed using the comparative Ct method [38].
4.7. Test of Pre-Harvest Sprouting
After pollinating the phs3-2 and phs3-12 null lines for pre-harvest sprouting tests, the panicles of 35-day-old plants were collected and immersed in water for 4 h. After that, the cotton swab was sufficiently immersed in water, placed on a tray, and the dipped rice panicles were placed on the cotton swab and incubated at 28 °C for 24 h under light conditions. The wet swab was replaced every 24 h to minimize contamination. The seed germination rates of the panicles were examined on days 4, 6, 8, and 11, respectively.
4.8. Determination of ABA
Determination of ABA with 2-week-old seedlings was performed as previously described [39].
4.9. Statistical Analysis
The data collected in this study were analyzed by one-way analysis of variance (ANOVA) using the statistical analysis system (SAS version 9.4). Data values were mean ± SE (n = 3) and statistical significance was set to p < 0.05, and post-analysis was performed according to Duncan’s multiple range test [40].
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Siebenmorgen T.J. Grigg B.C. Lanning S.B. Impacts of preharvest factors during kernel development on rice quality and functionality Annu. Rev. Food Sci. Technol.2013410111510.1146/annurev-food-030212-18264423464570 · doi ↗ · pubmed ↗
- 2Chang C. Zhang H. Lu J. Si H. Ma C. Genetic improvement of wheat with pre-harvest sprouting resistance in China Genes 20231483710.3390/genes 1404083737107595 PMC 10137347 · doi ↗ · pubmed ↗
- 3Gubler F. Millar A.A. Jacobsen J.V. Dormancy release, ABA and pre-harvest sprouting Curr. Opin. Plant Biol.2005818318710.1016/j.pbi.2005.01.01115752999 · doi ↗ · pubmed ↗
- 4Finkelstein R. Reeves W. Ariizumi T. Steber C. Molecular aspects of seed dormancy Annu. Rev. Plant Biol.20085938741510.1146/annurev.arplant.59.032607.09274018257711 · doi ↗ · pubmed ↗
- 5Mc Carty D.R. Genetic control and integration of maturation and germination pathways in seed development Annu. Rev. Plant Biol.199546719310.1146/annurev.pp.46.060195.000443 · doi ↗
- 6Assmann S.M. Snyder J.A. Lee Y.R.J. ABA-deficient (aba 1) and ABA-insensitive (abi 1-1, abi 2-1) mutants of Arabidopsis have a wild-type stomatal response to humidity Plant Cell Environ.20002338739510.1046/j.1365-3040.2000.00551.x · doi ↗
- 7Paek N.C. Lee B.M. Bai D.G. Smith J.D. Inhibition of germination gene expression by Viviparous-1 and ABA during maize kernel development Mol. Cells 1998833634210.1016/S 1016-8478(23)13433-99666472 · doi ↗ · pubmed ↗
- 8Cunningham F.X.Jr. Gantt E. Genes and enzymes of carotenoid biosynthesis in plants Annu. Rev. Plant Biol.19984955758310.1146/annurev.arplant.49.1.55715012246 · doi ↗ · pubmed ↗