Genome-Wide Analysis of the Cis-Prenyltransferase (CPT) Gene Family in Taraxacum kok-saghyz Provides Insights into Its Expression Patterns in Response to Hormonal Treatments
Liyu Zhang, Huan He, Jiayin Wang, Pingping Du, Lili Wang, Guangzhi Jiang, Lele Liu, Lu Yang, Xiang Jin, Hongbin Li, Quanliang Xie

TL;DR
This study identifies and analyzes the CPT gene family in a rubber-producing plant, revealing how these genes respond to hormones and contribute to rubber biosynthesis.
Contribution
The study provides a genome-wide analysis of the TkCPT/CPTL gene family and its expression patterns under hormonal treatments in Taraxacum kok-saghyz.
Findings
Eight CPT and two CPTL genes were identified with five conserved structural domains.
Fragment duplication is the main cause of gene family amplification in TKS.
TkCPT5 and TkCPT6 show significant expression increases under ethylene and methyl jasmonate treatments.
Abstract
Taraxacum kok-saghyz (TKS) is a natural rubber (NR)-producing plant with great development prospects. Accurately understanding the molecular mechanism of natural rubber biosynthesis is of great significance. Cis-prenyltransferase (CPT) and cis-prenyltransferase-like (CPTL) proteins catalyze the elongation of natural rubber molecular chains and play an essential role in rubber biosynthesis. In this study, we performed a genome-wide identification of the TkCPT/CPTL family, with eight CPT and two CPTL members. We analyzed the gene structures, evolutionary relationships and expression patterns, revealing five highly conserved structural domains. Based on systematic evolutionary analysis, CPT/CPTL can be divided into six subclades, among which the family members are most closely related to the orthologous species Taraxacum mongolicum. Collinearity analyses showed that fragment duplications…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
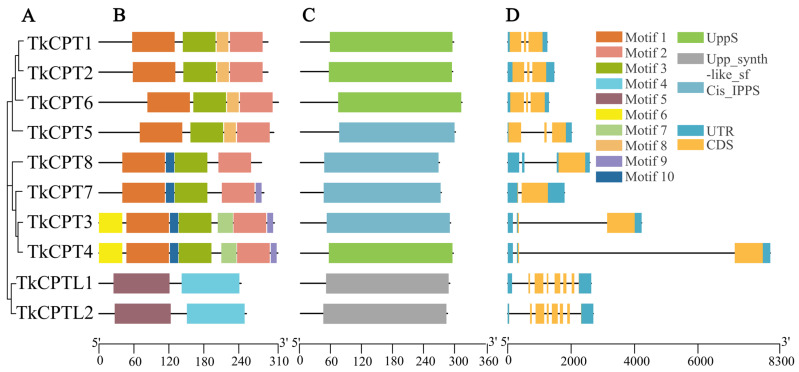 Figure 1
Figure 1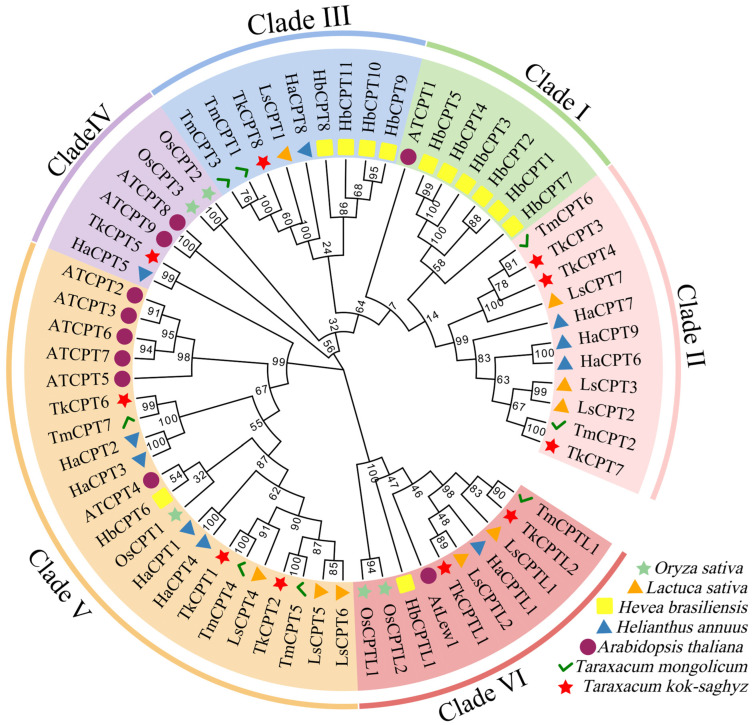 Figure 2
Figure 2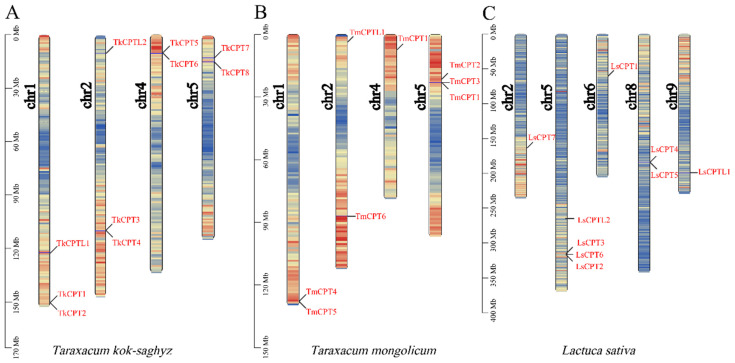 Figure 3
Figure 3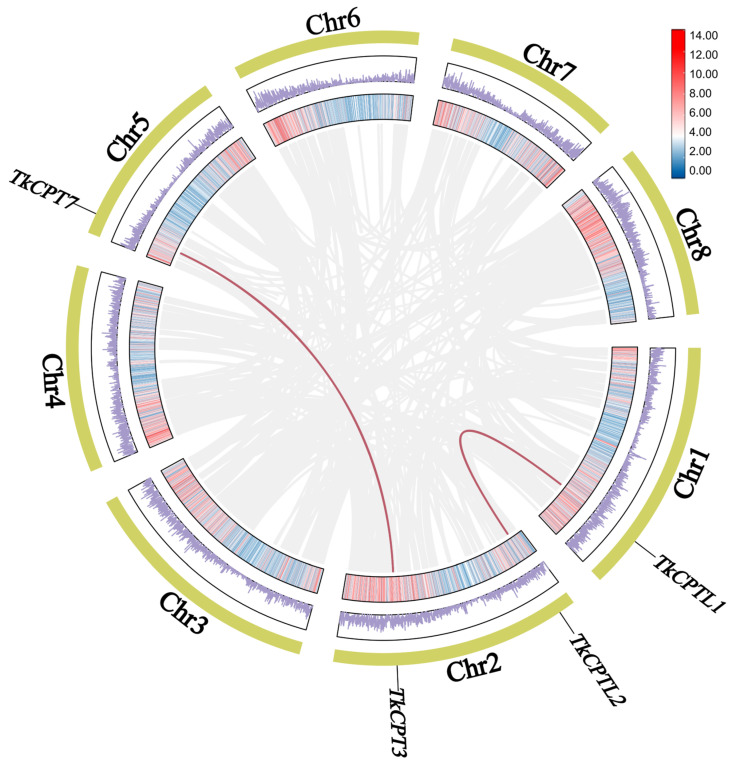 Figure 4
Figure 4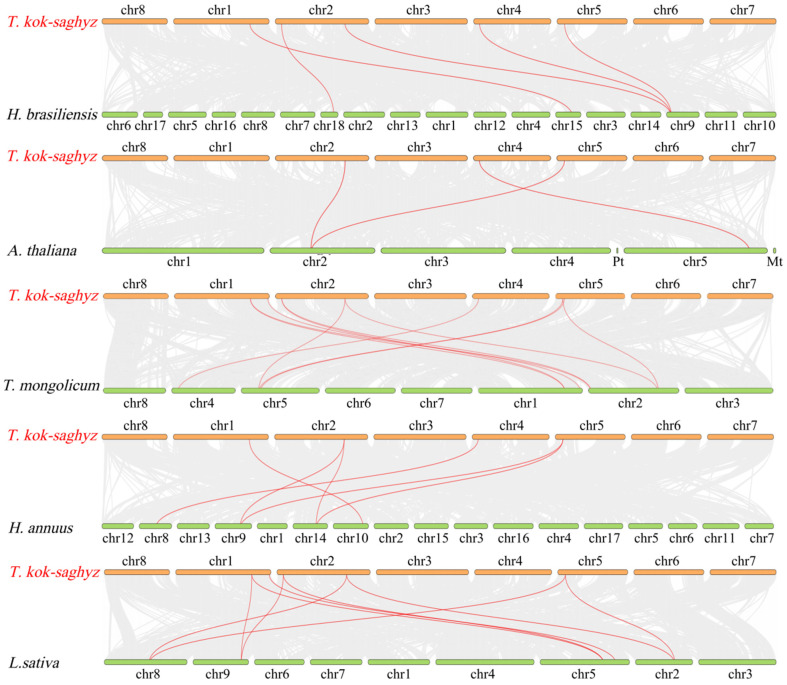 Figure 5
Figure 5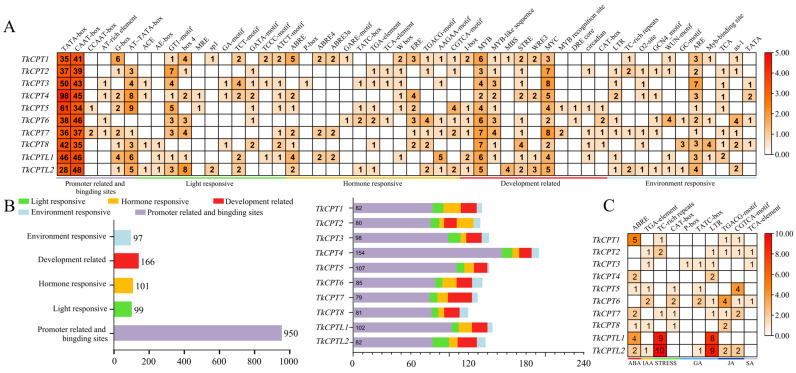 Figure 6
Figure 6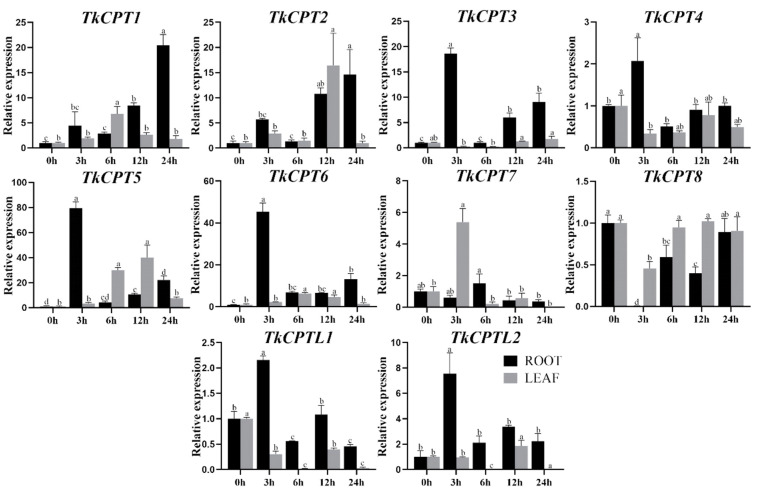 Figure 7
Figure 7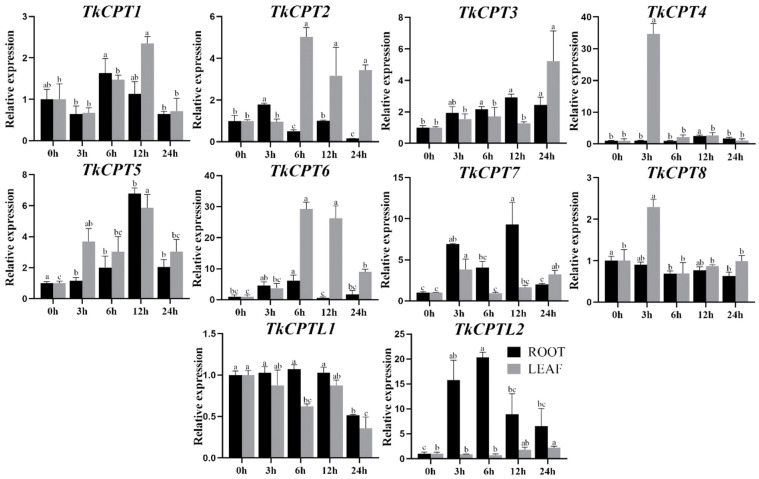 Figure 8
Figure 8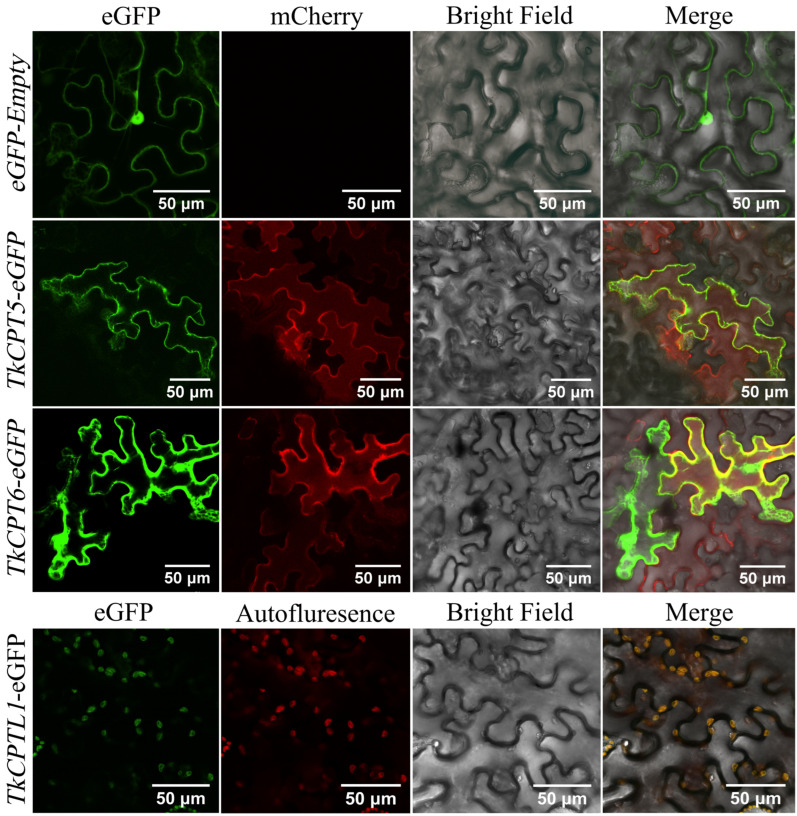 Figure 9
Figure 9 Figure 10
Figure 10 Figure 11
Figure 11 Figure 12
Figure 12 Figure 13
Figure 13 Figure 14
Figure 14 Figure 15
Figure 15 Figure 16
Figure 16 Figure 17
Figure 17 Figure 18
Figure 18 Figure 19
Figure 19- —the National Natural Science Foundation of China
- —Xinjiang Production and Construction Corps Guiding Technology Program Project
- —New Variety Breeding Special Project of Shihezi University
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsPlant biochemistry and biosynthesis · RNA and protein synthesis mechanisms · Microbial Natural Products and Biosynthesis
1. Introduction
Taraxacum kok-saghyz L. Rodin (TKS) is a small, diploid, sexually reproducing perennial plant in the Asteraceae family. As early as 1931, it was discovered in valleys of the Tianshan Mountains in southeastern Kazakhstan [1]. T. kok-saghyz typically thrives in non-saline or slightly saline floodplain meadows and agricultural canals [2] and can be cultivated in most temperate regions [1]. Hevea brasiliensis is the only commercially viable rubber producer that yields high-quality natural rubber (NR) [3]. Nevertheless, due to the susceptibility of H. brasiliensis to pathogen infections and its low genetic diversity [4], the exploration of alternative natural rubber resources is imperative. Compared to Brazilian rubber trees, T. kok-saghyz has a higher molecular weight of natural rubber [5]. Furthermore, it also has the advantages of a wider planting area, better quality, easier harvesting, and shorter growth cycles, potentially making it a commercially viable alternative to natural rubber as a raw material [6].
Two primary metabolic pathways for the synthesis of isoprenoids in plants have been defined: the methylerythritol (MEP) pathway and the mevalonate (MVA) pathway. Natural rubber is composed of isopentenyl pyrophosphate (IPP)-derived isopentenyl monomers, which are synthesized mainly by the cytoplasmic MVA pathway [7]. The tissues in T. kok-saghyz that produce NR are known as laticifers, which contain large quantities of rubber particles (RPs) [8]. Rubber particles are usually spherical or pear-shaped [9], with many proteins attached to the lipid membrane of the rubber particles [10]. This includes proteins, such as cis-prenyltransferase-like (CPTL) protein, small rubber particle protein (SRPP), and rubber elongation factor (REF) [11,12]. Cis-prenyltransferase (CPT), the main component of the rubber transferase (RTase) complex [13], is responsible for the construction of the long hydrocarbon skeleton, which extends the short, full-chain oligopentenyl diphosphate precursor by the sequential addition of the desired number of isopentenyl diphosphate (IPP) molecules which results in the formation of a stretch of cis units [14], which catalyze the elongation of the molecular chain of natural rubber. However, CPTs are incapable of producing high-molecular-mass NR alone [15], and they must interact with other proteins such as CPTL, REF and SRPP to achieve sufficient catalytic activity [16].
CPT, cis-prenyltransferase, also known as H. brasiliensis rubber transferase (HRT), is present as prokaryotic CPT in bacteria in the form of undecylenic pentanediol, which functions as a glycosyl carrier in the biosynthesis of peptidoglycan cell walls in bacteria. In plant plastids, prokaryotic CPT is expressed in the form of polyisopentyl glycol, which affects the fluidity of the membrane of vesicles and photosynthesis in plant plastids [17]. The CPT gene family has been relatively conservative throughout the course of evolution and can be classified into three categories based on chain length: short chain (C15), medium chain (C50–C55), and long chain (C70–C120) [14]. CPT is divided into two categories based on its enzyme composition: homodimeric enzymes (short-chain and medium-chain enzymes) and heterodimeric enzymes (including long-chain enzymes and rubber synthase) [18]. It has been reported that the gene encoded by CPT was first cloned in micrococcus luteus B-P in 1989 [19]. In 2003, Asawatreratanakul isolated and identified two genes, HRT1 and HRT2, encoding cis-isopentenyl transferases from the latex of H. brasiliensis. However, expression of recombinant proteins of HRT2 by the prokaryote Escherichia coli revealed that HRT2 did not exhibit rubber-transferring enzyme activity in vitro [20]. In 2011, it was reported that the human CPT formed a heterogeneous protein complex with the Nogo-B receptor and became active in dolichol biosynthesis. In 2012, scientists expressed recombinant proteins of HRT1 and HRT2 in the protoplasts of Saccharomyces cerevisiae and Arabidopsis thaliana, exhibiting cis-isopentenyl transferase activity, but did not catalyze the formation of high-molecular-weight natural rubber [18]. The cis-isopentenyl transferase-like enzyme, also known as CPT bridge protein (CPTBP), was shown by Yamashita in 2016 to form a protein complex with CPT, CPTL, and REF, which is involved in the biosynthesis of natural rubber [21]. In the same year, similar complexes consisting of CPT and its binding protein homologues were identified in Lactuca sativa, tomato and A. thaliana [22,23], whereas the dehydropolyphenol bisphosphate synthetases of human NgBR/hCIT and yeast Nus1/Rer2 [24,25] also belong to the group of cis-isoprenyltransferases. CPTL, as a necessary component of the rubber transferase complex, can promote NR biosynthesis by interacting with CPT [26]. Other components of the rubber transferase complex, such as SRPPs, were mainly located on small rubber particles and played a role in stabilizing the structure of rubber particles [27]. REF was also abundantly located on the surface of rubber particles and can improve CPT activity [28].
There have been no studies comprehensively reporting the genome-wide identification and analysis of the CPT/CPTL gene family in T. kok-saghyz, and its specific role is unknown. To systematically investigate the effect of the TkCPT/CPTL genes on the biosynthesis of T. kok-saghyz natural rubber, we have proposed a genome-wide characterization of the TkCPT/CPTL family members, having analyzed the physicochemical properties, basic structure, and evolutionary relationships of the family members. After treatment with ethylene and methyl jasmonate, members showed different expression patterns. The subcellular localization of some family genes was observed. These results provide new ideas for further studying the effects of exogenous plant hormones on the biosynthesis of natural rubber.
2. Results
2.1. Identification of the CPT/CPTL Gene Family in T. kok-saghyz
A total of eight TkCPTs and two TkCPTLs were identified in the TKS genome, which were named TkCPT1~TkCPT8 based on their sequential order in the chromosome. The predicted relevant physicochemical properties are shown in Table 1. The CDS length ranged from 735 bp (TkCPTL1) to 933 bp (TkCPT6), the amino acid residue length ranged from 244aa (TkCPTL1) to 307aa (TkCPT4), the molecular weight ranged from 28.55431 kDa (TkCPTL1) to 35.00990 kDa (TkCPT6), and the isoelectric point prediction ranged from 5.91 (TkCPT1) to 9.32 (TkCPT7). The isoelectricity of TkCPT1 was <7, indicating that it was an acidic protein. The instability index was 31.24 (TkCPT4) to 53.56 (TkCPT7), and the instability indexes of TkCPT3, TkCPT4 and the two TkCPTL members were <40, which indicated that the four proteins were stable. The maximum value of the aliphatic index was 104.39 (TkCPTL2), the minimum value was 80.00 (TkCPT6), and the indices of TkCPT2, TkCPTL1 and TkCPTL2 were all >100. A possible explanation for these results may be that they had better solubility and thermal stability. The results of hydrophilicity analysis showed that the total hydrophilicity indices of the TkCPT/CPTL family proteins all ranged from −0.41 to −0.1; all of them were hydrophilic proteins. The prediction of subcellular localization of the proteins using online software showed that the TkCPT family proteins were localized in the endoplasmic reticulum, whereas TkCPTL1 was localized in the chloroplasts, and TkCPTL2 was localized in the cytoplasm (Table 1).
2.2. Secondary and Three-Dimensional Structures Analysis of the TkCPT/CPTLs
The results showed that most of the secondary structures of the proteins of TkCPT/TkCPTLs family members were α-helices, followed by irregularly coiled and β-turns, with the proportion of α-helices ranging from 30.69% (TkCPT2) to 53.41% (TkCPT8), while β-turns ranged from 3.87% (TkCPT6) to 6.90% (TkCPT1), and irregularly coiled ones ranging from 25.62% (TkCPT7) to 39.31% (TkCPT2) varied; the proportion of irregular curls varied from 25.62% (TkCPT7) to 39.31% (TkCPT2) (Table 2). The number of amino acids in each structure accounted for roughly the same percentage, while the secondary structures of each TkCPT/TkCPTL family member were similar to a certain extent, such as TkCPT1, TkCPT2, TkCPT3, and TkCPT4, which were more consistent in terms of their secondary structures.
The three-dimensional structures of eight TkCPT proteins and two TkCPTL proteins were predicted using homology modeling, and the results revealed that, except for TkCPT5 and TkCPT6, there were significant similarities between different members of TkCPT/CPTLs, and thus it could be inferred that these members might have similar protein functions. The three-dimensional structures of TkCPT5 and TkCPT6 were more distinct from the other proteins, but highly similar to each other, possibly due to divergence during evolution (Figure S1).
2.3. Evolutionary Relationship, Conserved Structure, Motif Composition, and Gene Structures of TkCPT/CPTLs
Evolutionary analyses showed that TkCPT3 was more closely related evolutionarily to TkCPT4, and TkCPT1 was more closely related to TkCPT2 (Figure 1A). Conserved motif analysis of TkCPT proteins showed that motif 1, motif 2, and motif 3 were present in all TkCPTs (Figure 1B), and in particular, all family members except TkCPT6 contained motif 4. The two members of TkCPTL, on the other hand, had fewer motifs present, only motif 4 and motif 5, but were quite conserved. Analysis of the structural features of TKCPT/CPTL revealed that all proteins contain UPPS structural domains or belong to the UPPS superfamily (Figure 1C). TkCPT family members have different positions and lengths of untranslated regions (UTRs) and coding sequences (CDSs), with the number of CDSs ranging from one to three, and TkCPTL members have more CDSs, totaling six. Except for TkCPT4, members of the same evolutionary branch had similar UTR and CDS features (Figure 1D).
2.4. Evolutionary Analysis of TkCPT/CPTL Proteins
To further explore the evolutionary relationship between CPT/CPTL families, a total of six species, including Helianthus annuus, Lactuca sativa and T. mongolicum, which belong to the same family of Asteraceae, as well as the model plants A. thaliana, H. brasiliensis and Oryza sativa from the family of Gramineae, were compared with T. kok-saghyz. Eight CPT members of T. kok-saghyz, seven CPT members of T. mongolicum, nine CPT members of A. thaliana, nine CPT members of H. annuus, eleven CPT members of H. brasiliensis, seven CPT members of L. sativa, three CPT members of O. sativa, and a total of ten CPTL members in seven species, respectively, and a total of sixty-four CPT proteins were identified; CPT members showed large differences in numbers across species. These proteins could be divided into six subclades. Proteins in the same subclade may perform similar biological functions. Group IV was the smallest among all the subclades, containing only six proteins, Group I contained seven proteins, Group III contained nine proteins, all CPTL members were present in Group VI and there were no other CPT members, and group V had twenty-one proteins (Figure 2).
2.5. Chromosomal Localization of TkCPT/CPTL Genes
In order to better show the evolutionary pattern of the CPT/CPTL gene family, chromosomal localization of the genomic distribution of the CPT/CPTL family genes in T. kok-saghyz as well as in T. mongolicum and L. sativa of the same Asteraceae family was performed. The TKS genome has a total of eight chromosomes, and members of the TkCPT/CPTL family were unevenly distributed on four of these chromosomes. All genes were located at the ends of chromosomes and varied in number from two to three: chr1 and chr2 have three, chr4 and chr5 have two. Gene duplication events were present in TkCPT1 and TkCPT2, TkCPT3 and TkCPT4, and TkCPT5 and TkCPT6. The evolutionary relationship subclades were very similar between the species. The CPT genes of T. mongolicum were all equally dispersed on all four chromosomes, and the genes were present in locations very similar to the distribution of TkCPT/CPTL on the chromosomes. The CPT genes of L. sativa were present on different chromosome numbers, with one LsCPT present on both chr2 and chr6, two on chr8, and three on chr5 (Figure 3), and the distribution was more dispersed, with most of them near the middle of the chromosomes.
2.6. Gene Duplication, and Collinearity Analysis of TkCPT/CPTL Genes
Tandem duplications and segmental duplications were the main causes of gene amplification in plants, and collinearity between species can be used to identify the location of orthologous genes. To explore the amplification and evolutionary mechanisms of the TkCPT/CPTL gene family, we analyzed the collinearity of the TkCPT/CPTL gene family. A total of two collinear blocks, TkCPT3/CPT7 and TkCPTL1/CPTL2, were identified. Homologous genes perform the same function (Figure 4). The prediction of familial gene duplication events revealed that both pairs of genes were caused by segmental duplication. The other members had a variety of duplication types, with TkCPT1 and TkCPT2 being tandem duplications and TkCPT4, TkCPT5, and TkCPT6 being proximal duplication types. To further analyze the selection pressure between TkCPT/CPTL genes, we calculated the Ka/Ks ratios of these two gene pairs for non-synonymous substitutions (Ka) and synonymous substitutions (Ks), where the Ka of TkCPT3/CPT7 was 0.16 and the KS was 1.24; the Ka of TkCPTL1/CPTL2 was 0.21 and the KS was 0.83, indicating that both pairs of this family had a Ka/Ks ratio less than one. These data suggested that members of this family had been selected for stronger purifying selection (Table 3).
To further understand the mechanism of gene duplication of TkCPT/CPTLs and to study the evolution of gene mutation, we constructed a gene duplicator including T. mongolicum of the same genus, L. sativa and H. annuus of the same Asteraceae family, A. thaliana, which was a model plant, and the rubber plant H. brasiliensis. The results showed that there were eleven pairs of collinear genes between T. kok-saghyz and T. mongolicum, nine pairs of collinear genes with L. sativa, three pairs of collinear genes with A. thaliana, six pairs of collinear genes with H. annuus, and five pairs of collinear genes with H. brasiliensis. It is noteworthy that TkCPT3 and TkCPT7 had collinear gene pairs in all species (Table S1), which may have existed earlier in the evolutionary process, and these findings suggest that TkCPTs had the highest gene homology with T. mongolicum and were more closely related (Figure 5).
2.7. Promoter Analysis of TkCPT/CPTL Genes
Promoter cis-acting elements are important transcription initiation binding regions that play an important role in the regulation of gene expression. To understand the possible regulatory mechanisms of TkCPT under hormone treatment and abiotic stress, we predicted cis-acting elements in the 2000 bp region upstream of the TkCPT/CPTL gene using the online database Plant CARE, using a total of 52 cis-acting elements, including TATA-box, CAAT-box, and CCAAT-box (promoter-related); ACE, GT1-motif, and box four (light-responsive element-related); ABRE element (related to abscisic acid); GARE-motif, and TATC-box (gibberellin-responsive element); TGA element (growth-hormone-responsive element); CGTCA-motif (involved in the MeJA reaction) and TGACG-motif; TCA element (involved in the salicylic acid reaction) etc. (Figure S3). All TkCPT/CPTL genes except TkCPT1, TkCPT5 and TkCPT8 possessed LTR elements, suggesting that these genes may play a role in cold and drought. These cis-acting elements were classified into five categories, including eleven environmental-stress-related elements, eleven development-related elements, thirteen hormone-responsive elements, eleven light-responsive elements, and six promoter-related elements. Among them, promoter-related and binding-site elements were the most numerous, accounting for 60.66 percent of all cis-acting elements, and light-responsive elements were the least numerous, accounting for only 6.02 per cent. Further analysis of some hormone-related cis-acting elements showed that only TkCPT3 possesses a P-box, which is related to salicylic acid associated with plant defence against pathogens, and TkCPT5 and TkCPT6 possess the CGTCA-motif or TGACG-motif, both of which were responsive to jasmonate (Figure 6), which will provide a basis for the subsequent study of the expression characteristics of TkCPT/CPTL genes after hormone treatment and function verification.
2.8. qRT-PCR Analysis of TkCPT/CPTL Genes
Quantitative real-time fluorescence PCR was used to detect the expression of TkCPT/CPTL in different tissues. The results showed that the root expression of TkCPT1, TkCPT2, TkCPT3, TkCPT5 and TkCPT6 was significantly upregulated after ethylene induction. Among them, the expression of TkCPT5 and TkCPT6 was the most significantly upregulated compared with the control, which was presumed to play a key role in the ethylene-regulated pathway, and the addition of exogenous hormones stimulated the expression of these genes. TkCPT7 and TkCPT8 were gradually downregulated and basically all TKCPT/CPTLs were significantly upregulated within 3 h of treatment, followed by a significant downregulation of expression at 6 h. TkCPT8 was unique, with downregulation of expression at 3 h and upregulation of expression at 6 h, which was different from the expression pattern of other genes, and it was speculated that it might have other biological functions. In leaves, the expression of genes such as TkCPT1 and TkCPT5 mostly increased but was less pronounced compared to that of roots, and the experimental results suggest that these genes may be involved in regulatory pathways of ethylene metabolism that regulate growth and developmental processes in specific tissues (Figure 7).
In addition, we treated the roots and leaves of TKS with methyl jasmonate to detect the expression of TkCPT/CPTL genes. In roots, TkCPT6, TkCPT7, and TkCPTL2, were upregulated at higher levels (Figure 8). The expression of TkCPT3 gradually increased with treatment time. Some genes reached the highest expression level at 3 h and 6 h equally, whereas in leaves, some genes reached the highest expression level at 3 h and 6 h. The expression of different members varied by tissue. Only TkCPT8 showed slightly reduced expression levels in roots and leaves and remained at low levels. The results showed that these genes were significantly expressed under methyl jasmonate induction and highly expressed in TKS roots. These experimental results suggested that the TkCPT/CPTL family was involved in the regulation of jasmonic acid metabolism regulatory pathways.
2.9. Subcellular Localization of TkCPT/CPTL
To further explore the functions of TkCPT and TkCPTL proteins, we performed subcellular localization to verify the specific locations where they exert their functions. The TkCPT-pCAMBIA1300-eGFP gene was co-infected with endoplasmic reticulum markers and expressed instantaneously. In the superimposed field, the fluorescence of TkCPT5 and TkCPT6 overlapped within the endoplasmic reticulum appearing as yellow, while TkCPTL1 fluorescence was concentrated on chloroplasts (Figure 9), which was consistent with the predicted results.
3. Discussion
Natural rubber is an important high-performance material with unparalleled advantages over synthetic rubber and is widely used in industries such as transportation, medicine and defense [5]. Cis-prenyltransferase (CPT) is a major component of the rubber transferase complex on the surface of rubber particles, catalyzing the extension of the molecular chain of natural rubber [27], whereas cis-prenyltransferase-like enzymes (CPTLs) interact with CPTs to enhance the stability of the CPTs in the ER membranes [29], thus boosting the activity of the CPTs [30]. To thoroughly investigate the function of the CPT/CPTL family in TKS, this study conducted a genome-wide identification of the TkCPT/CPTL family based on public genome data, and a total of ten members of the TkCPT/CPTL family as well as six other species were identified: H. annuus, L. sativa, T. mongolicum, A. thaliana, H. brasiliensis and O. sativa. A total of 64 CPT/CPTL members were divided into six subclades based on their evolutionary relationships (Figure 2). CPT members were distributed in various subclades and were most closely related to H. annuus and L. sativa of the same Asteraceae family. O. sativa is a graminaceous herbaceous plant, and the number of CPT members was sparse and concentrated. Most members of the rubber tree HbCPT are concentrated in I and III, and among the nine AtCPTs that have been published and identified [14,31], the ACPT (NP.565551) protein sequence contributes to the biosynthesis of long-chain polyisoprenoids [32,33]. LsCPT7 is present on the endoplasmic reticulum in subcellular localizations in tobacco, and the yeast two-hybrid data show that it is closely related to LsCPTL2. The yeast microsomes of LsCPTL2 promote the synthesis of short-cis polyisoprene [34]. In the evolutionary tree, LsCPT7 had evolutionary affinities with TkCPT3 and TkCPT4, and it was speculated that TkCPT3 and TkCPT4 may have an important role in rubber synthesis (Table S2).
The amino acid sequence of a protein determines how its higher structure is formed and its ultimate function. Multiple sequence comparisons (Figure S4) showed that all TkCPT protein sequences have five characteristically conserved regions, which is the same as previously reported [35], and most of the fully conserved amino acids are involved in catalysis, substrate binding or structural interactions. Some of the important residues, such as Asp (D) in the region I and Asn (W) in region III [36] are highly conserved (100%) and are key amino acid residues related to catalytic activity; Phe (F) (71%) and Ser (S) (71%) in region III, Asp (D) in region IV, Glu (E) in region V [21] and two Arg (R) (100% and 79%) residues [37] are essential for IPP binding. The two members of the TkCPTL do not have the conserved structural domains that are present in both CPT sequences, although some of the bases are also catalytic. The prediction of the protein transmembrane regions showed that all the CPT members did not have a transmembrane helical structure, and interestingly, the two TkCPTL members both have a transmembrane helical region, which might be related to the transport of their special substances or signal transduction (Figure S2 and Table S3). Variation in gene structure is an important feature of family evolution; therefore, we analyzed the basic structure of TkCPT/CPTL and identified a total of ten conserved motifs. TkCPT possesses three highly conserved motifs, which are present in all TkCPTs. Motif 6 and motif 8 are only present in TkCPT3, TkCPT4, TkCPT7. While motif 5 was present in TkCPT3, TkCPT4, TkCPT7 and TkCPT8, which might be related to the unique function of the gene itself (Figure 1B), the rest of the members of the groups had a more similar motif structure. The localization of TkCPT/CPTL families on chromosomes is more dispersed and more gene duplication events occur, which is more similar to the distribution of T. mongolicum on chromosomes, and it is likely that some gene divergence has also occurred during the evolutionary process (Figure 3). Duplicated genes are often subject to some selective pressure to adapt to the external environment. Here, we calculated the Ka/Ks ratio of the two gene pairs of TkCPT/CPTL, and both gene pairs had Ka/Ks values less than one and underwent purifying selection (Table 3). TkCPT/CPTL are amplified in a variety of modes, with fragment repeats and tandem repeats being the main modes (Table S4), which play an important role in the amplification of the number of genes in the family [38,39]. Of course, CPT/CPTL exercise different functions in different species and environments, and in humans, a subunit of cis prenyltransferase causes a congenital disorder of glycosylation [40]. In the para rubber tree genome, eleven CPT members were determined, but only HbCPT2 protein shows rubber transferase activity [41]. CPTL, in addition to its known functions related to rubber synthesis and its homologue in Arabidopsis (LEW1) were identified in a genetic screen for leaf chlorosis phenotypes. LEW1 mutants are unable to synthesize ethanol efficiently, resulting in defective O- and n-glycosylation of the protein. The results suggest that CPT-like proteins are important in alcohol synthesis in eukaryotes [42].
It has been shown that highly conserved cis-elements are present in many plants and that cis-elements play crucial roles in transcriptional regulatory signaling pathways when plants are subjected to biotic and abiotic stresses. To further elucidate the functions of these genes, we analyzed the promoter regions of the TkCPT/CPTL family. The cis-acting elements in the promoter region include various types of promoter binding site-related elements (G-box, CCAAT-box, etc.), light-responsive elements (GA-motif, ATCT-motif, etc.), growth and development-related elements (STRE, CAT-box, etc.), and hormone-related responsive elements containing ABA (ABRE), of which TkCPT1 contains more, which is hypothesized to play an important role in plant resistance to environmental stresses and pathogen infestation. There are also various hormone response elements such as IAA (TGA-element), STRESS (TC-rich repeats, CAT-box), GA (P-box, TATC-box, LTR), JA (TGACG-motif, CGTCA-motif), and SA (TCA-element) (Figure 6), which may play a role in the MYC homeopathic effectors that play important roles in the regulation of gene expression. For example, ABA enhances drought tolerance in plants by binding to MYC cis-acting elements and initiating the expression of downstream genes containing the same kind of elements. The number of ABRE and MYC cis-acting elements in TkCPT1 is quite high, which may subsequently provide new ideas for the study of key genes involved in the increase in T. kok-saghyz rubber production under drought conditions. In a previous report, CPT and CBP (CPTL) genes were found to be downregulated in expression under drought stress by transcriptome analysis of Parthenium hysterophorus rubber particles (PR) [43], and it was speculated that this might be related to their involvement in biosynthesis of polyprenol or plastidic polyprenol, which is essential for photosynthesis [44].
Ethylene and MeJA are widely involved in regulating plant growth and development and secondary metabolite synthesis [45], exogenous ethylene promotes seed germination and rooting [46], and MeJA induces the differentiation of laticifer in H. brasiliensis, which is an important signal regulating the differentiation and development of laticifer, and exogenous JA enhances the enzyme lipoxygenase in H. brasiliensis latex [47]. We treated different tissues of TKS with ethylene and MeJA and performed qRT-PCR to detect the expression levels of TkCPT/CPTL genes and found that both hormones induced the expression of TkCPT/CPTL genes in the roots of TKS and that the expression levels of TkCPT5 and TkCPT6 were upregulated to a higher level after treatment. In addition, both hormones affect the expression of TkCPT/CPTL genes in TKS leaves, and overall, MeJA had a greater effect on gene expression in TkCPT/CPTL leaves than in roots. Interestingly, TkCPT3 and TkCPT4 did notcontain MeJA-responsive elements, but the qRT-PCR results showed a corresponding increase in their gene expression. It has been well-documented that jasmonic acid and methyl jasmonate play a key role in natural rubber biosynthesis and response to abiotic stresses in T. kok-saghyz [48,49,50]. It has been shown that MeJA can stimulate late secondary metabolism and increase NR content in TKS via the MVA pathway, and that HMGR, FPPS, GGPPS, IDI and ISPF are essential for rubber biosynthesis in the MVA pathway. The transcriptomic data in MEJA-treated TKS roots showed that the expression of these genes was high, and CPT was located downstream of the GGPPS gene, which might be related to its increased expression [12,51].
Based on this, we have a more comprehensive understanding of the TkCPT/CPTL gene families, which provides ideas for subsequent studies to verify the functions of TkCPT/CPTL.
4. Materials and Methods
4.1. Plant Materials and Processing Methods
T. kok-saghyz (Shihezi City, Xinjiang, China) was selected as the experimental material, cultivated in a growth chamber at 25 ± 2 °Cand 30% humidity, and transplanted in nutrient soil (nutrient soil: perlite: vermiculite = 3:1:1) 15 days after germination. Six-month-old T. kok-saghyz was selected as the treatment material, and the roots were immersed into Hoagland’s culture solution of 1 mmol/L methyl jasmonate and 100 μmol/L ethylene. The roots and leaves were collected at the time periods of 0 h, 3 h, 6 h, 12 h, and 24 h, the collected material was immediately placed in liquid nitrogen and stored at −80 °C for preservation, which facilitated the later experimental treatments. The natural rubber in T. kok-saghyz was extracted using the alkaline boiling method [52,53].
4.2. Identification of TkCPT/CPTL Gene Family and Physicochemical Properties of the TkCPT/CPTL Proteins
The latest genome sequence and annotation files of T. kok-saghyz [54] were obtained from the website of the National Centre for Genome Sciences Research (https://ngdc.cncb.ac.cn/, accessed on 10 March 2023), and the identified A.thaliana CPT/CPTL protein sequences were firstly downloaded from the NCBI database as seed sequences, (https://www.ncbi.nlm.nih.gov/, accessed on 10 March 2023), which were screened using local blastp comparisons. Sequences with less than 70% similarity were removed. Secondly, the Hidden Markov Model (HMM) file of the CPT conserved structural domain (PF01255) was downloaded from the PFam database [55] (http://pfam.xfam.org/, accessed on 12 March 2023), and the candidate sequences were submitted to HMM-search. Finally, using Batch CD-Search [56] (https://www.ncbi.nlm.nih.gov/Structure/cdd/wrpsb.cgi/, accessed on 12 March 2023), protein sequences that did not contain complete structural domains were removed to identify TkCPT/CPTL gene family members. The molecular weight and isoelectric point of TkCPT/CPTL were analyzed using Expasy (http://www.expasy.org, accessed on 12 March 2023) [57]. In addition, we used Cell-PLoc 2.0 to predict the subcellular localization of TkCPT/CPTL proteins (http://www.csbio.sjtu.edu.cn/bioinf/Cell-PLoc-2/, accessed on 12 March 2023) [23]. Prediction of the number of transmembrane helices for TkCPT/CPTL proteins were performed using the TMHMM-2.0 web software (https://services.healthtech.dtu.dk/services/TMHMM-2.0/, accessed on 12 March 2023).
4.3. Evolutionary Analysis, Gene Structure, Motif, and Conserved Domain Analysis
The construction of evolutionary trees was performed using the maximum likelihood (ML) method with MEGA11 software, version 11.0. To assess the reliability of the tree, a bootstrap value was set to 1000 replicates [58] and applied to calculate the p-distance (Figure S2). Landscaping was performed using the online website EvolView (https://evolgenius.info/evolview, accessed on 20 March 2023) [59]. Motifs of TkCPT/CPTL proteins were analyzed using the online database MEME (https://meme-suite.org/tools/meme, accessed on 20 March 2023), using default parameters with the maximum number of conserved motifs set to 10 [60] (Figure S5). Finally, the gene structure was visualized using Gene Structure View (Advanced) in TBtools v1.099 [61].
4.4. Protein Multiple Sequence Comparison, Secondary Structure Prediction, 3D Structure Prediction
The protein sequences of TkCPT/CPTL were compared using Clustal W software v 1.81 and displayed using GeneDoc v 2.7.0.0 software [62]. The secondary structure of TkCPT/CPTL proteins was analyzed using the SOPMA website (https://npsa-pbil.ibcp.fr/cgi-bin/npsa_automat.pl?page=npsa_sopma.html, accessed on 12 March 2023) for analysis. The SWISS-MODEL website (https://swissmodel.expasy.org/interactive, accessed on 12 March 2023) in EXpasy was used to visualize the TkCPT/CPTL protein’s three-dimensional structure [57].
4.5. Multi-Species CPT/CPTL Chromosome Localization and Collinearity Analysis
TkCPT/CPTL gene-related data, including chromosome location data, chromosome length and gene start and end positions was employed. Chromosomal location distribution maps of CPT/CPTL genes in T. kok-saghyz, T. mongolicum and L. sativa were obtained using the Gene Location Visualize tool from the GTF/GFF module of the TBtools software v1.099. The simple Ka/Ks Calculator, a Ka/Ks calculator in the TBtools software, was used to calculate synonymous substitutions (Ks) and synonymous substitutions (Ka) and the ratio between them, with a divergence time of T = Ks/ (2 × 1.5 × 10^−8^) × 10^−6^ Mya [63]. Comparison of TkCPT/CPTL collinearity and collinearity with other species was performed using One Step MCScanX-SuperFast in TBtools software v1.099.
4.6. Cis-Acting Element Analysis
The sequence of 2000 bp upstream of the start codon of the TkCPT/CPTL genes was obtained using TBtools software v1.099 and submitted to the PlantCARE website (http://bioinformatics.psb.ugent.be/webtools/plantcare/html/, accessed on 2 April 2023) [64], and the results were imported into the Simple Biosequence Viewer program of the TBtools software v1.099 for visualization. In the HeatMap program of TBtools software v1.099, the number of cis-acting elements and the number of different types of hormone elements were produced, and GraphPad v9.5.1.733 software was used to produce bar charts.
4.7. qRT-PCR
Pre-treated plant material was obtained and RNA was extracted using a HiPure HP Plant RNA Mini Kit (Megan, Guangzhou, China) and reverse transcription kit (Vazyme, Nanjing, China) using the cDNA kit for each sample. Each reaction was conducted with three biological replicates. Primers were designed using the primer Premier 5.0 software (https://premierbiosoft.com/, accessed on 25 December 2023) using Tkβ-actin as the internal reference gene, and the reaction procedure was as follows: 95 °C for 15 s, 95 °C for 10 s, 54–62 °C for 20 s, and 72 °C for 20 s for 40 cycles of 95 °C for 5 s, 65 °C for 1 min, and 40 °C for 30 s. The data were quantified by the 2^−∆∆Ct^ calculation method to quantify the relative expression levels of the genes, and the data were statistically analyzed and visualized using GraphPad software 9.5.1. Statistical differences between measurements at different times or with different treatments were analyzed using Duncan’s multiple range test. Differences were considered significant at a probability level of p < 0.05.
4.8. Subcellular Localization
To study the transient expression of TkCPT/CPTLs in the leaves of tobacco, the full-length CDS of TkCPT/CPTLs was PCR-amplified using primers containing KpnI and SalI restriction enzyme (Table S5) and ligated into the KpnI/SalI vector pCAMBIA1300-eGFP to construct the pCAMBIA1300-35s-eGFP, endoplasmic reticulum localization plasmid tagged with pCAMBIA1300-35S-WAK2-mCherry-HDEL (Puint, Shaanxi, China). The constructed vectors were transformed into Agrobacterium tumefaciens GV3101, injected into the bottom of four-week-old tobacco leaves, and incubated in dark culture at 25 °C for 48 h, observed using a laser confocal microscope (Nikon, Tokyo, Japan), with excitation of the GFP. The excitation wavelength of GFP was 488 nm, the excitation wavelength of RFP was 561 nm, and chloroplast auto-fluorescence was 640 nm.
5. Conclusions
In this study, we comprehensively identified and characterized the CPT/CPTL family of T. kok-saghyz. We identified eight CPT genes (TkCPT1-TkCPT8) and two CPTL genes (TkCPTL1-TkCPTL2) in the TKS genome, distributed on four chromosomes. We analyzed their physicochemical properties, gene structures, protein secondary structures, protein three-dimensional structural models, colinear and cis-acting elements in detail. The results showed that all TkCPT genes had upps-related conserved structural domains, Evolutionary tree and covariance analysis provided new perspectives for the subsequent study of the CPT/CPTL gene family in TKS. In addition, the expression patterns of TkCPT/CPTL genes under the exogenous hormone ethylene and MeJA treatments were also investigated. This study provides important information for the further elucidation of the gene function of the CPT/CPTL family in TKS and provides a theoretical basis for subsequent functional validation and study of the regulatory mechanism of natural rubber biosynthesis.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Kirschner J. Štěpánek J. ČernýT. De Heer P. van Dijk P.J. Available ex situ germplasm of the potential rubber crop Taraxacum koksaghyz belongs to a poor rubber producer, T. brevicorniculatum (Compositae–Crepidinae)Genet. Resour. Crop. Evol.20126045547110.1007/s 10722-012-9848-0 · doi ↗
- 2Krotkov G. A review of literature on Taraxacum koksaghyz Rod Bot. Rev.19451141746110.1007/BF 02861139 · doi ↗
- 3Kleinhenz M.D. Miller A.R. Streeter J.G. Temporal diversity of Taraxacum koksaghyz plants reveals high rubber yield phenotypes Biodiversitas 20161784785610.13057/biodiv/d 170262 · doi ↗
- 4Ramirez-Cadavid D.A. Cornish K. Michel F.C. Taraxacum koksaghyz (TK): Compositional analysis of a feedstock for natural rubber and other bioproducts Ind. Crop. Prod.201710762464010.1016/j.indcrop.2017.05.043 · doi ↗
- 5van Beilen J.B. Poirier Y. Guayule and Russian dandelion as alternative sources of natural rubber Crit. Rev. Biotechnol.20072721723110.1080/0738855070177592718085463 · doi ↗ · pubmed ↗
- 6Ghaffar M.A.A. Meulia T. Cornish K. Laticifer and Rubber Particle Ontogeny in Taraxacum koksaghyz (Rubber Dandelion) Roots Microsc. Microanal.2016221034103510.1017/S 1431927616006012 · doi ↗
- 7Xie Q. Ma J. Ding G. Yuan B. Wang Y. He L. Han Y. Cao A. Li R. Zhang W. Transcriptomics and proteomics profiles of Taraxacum koksaghyz roots revealed different gene and protein members play different roles for natural rubber biosynthesis Ind. Crop. Prod.202218111477610.1016/j.indcrop.2022.114776 · doi ↗
- 8Berthelot K. Lecomte S. Estevez Y. Peruch F. Hevea brasiliensis REF (Hev b 1) and SRPP (Hev b 3): An overview on rubber particle proteins Biochimie 20141061910.1016/j.biochi.2014.07.00225019490 · doi ↗ · pubmed ↗