DNA Methylation Regulatory Axis miR‐29b‐3p/DNMT3B Regulates Liver Regeneration Process by Altering LATS1
Yinwen Zhou, Hao Wu, Qiu Wang, Bo Ma, Jiulong Sun, Guoliang Wang

TL;DR
This study reveals how DNA methylation and miR-29b-3p regulate liver regeneration through the Hippo signaling pathway.
Contribution
The novel miR-29b-3p/DNMT3B regulatory axis is identified as a key driver of liver regeneration via LATS1 modulation.
Findings
DNMT3B regulates LATS1 protein expression through DNA methylation.
miR-29b-3p modulates DNMT3B and LATS1 to influence liver regeneration.
The miR-29b-3p/DNMT3B axis promotes liver regeneration by altering Hippo signaling.
Abstract
DNA methylation is a crucial epigenetic alteration involved in diverse biological processes and diseases. Hippo signalling pathway is a key signalling regulatory network in the growth and development of tissues and organs. Nevertheless, the precise role of DNA methylation and Hippo signalling pathway during liver regeneration (PH) is still unclear. In this study, we investigated the regulatory mechanism of LATS1, a pivotal protein in the Hippo signalling pathway, on liver regeneration and explored the specific mechanism of DNA methylation regulating LATS1. To analyse the regulation of LATS1 by DNA methylation, following 2/3 partial hepatectomy (PH) in liver‐specific AAV‐8 shDNMT3B deleted mice (DNMT3B, KD) mice and sex‐matched AAV‐8 shControl (Control). We determined that DNMT3B regulates the protein expression of LATS1 by DNA methylation. miR‐29b‐3p significantly regulates the…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
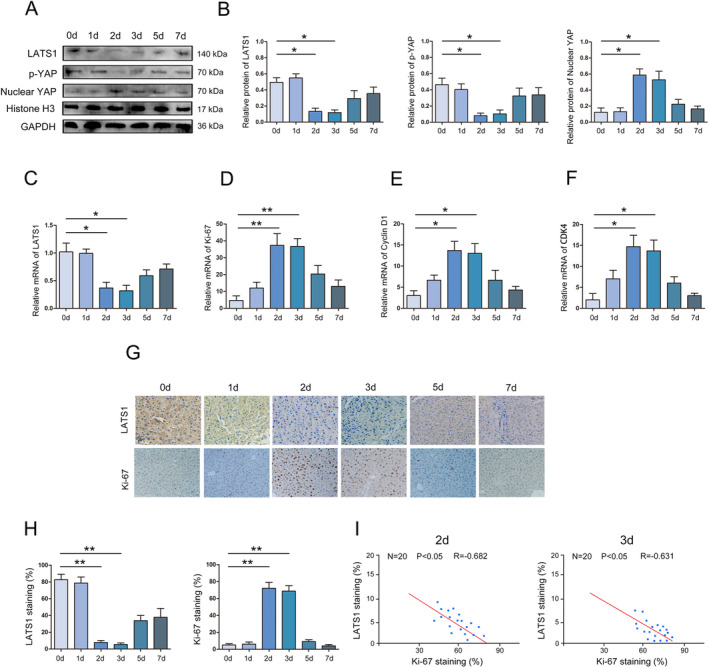 FIGURE 1
FIGURE 1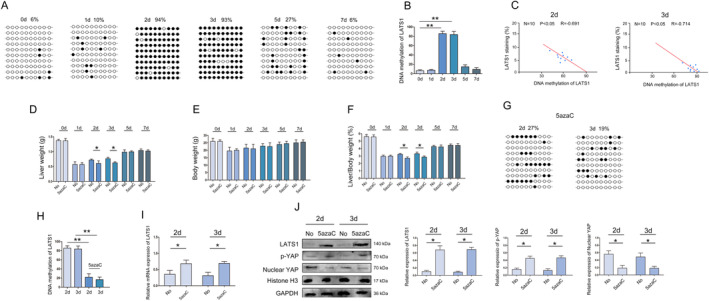 FIGURE 2
FIGURE 2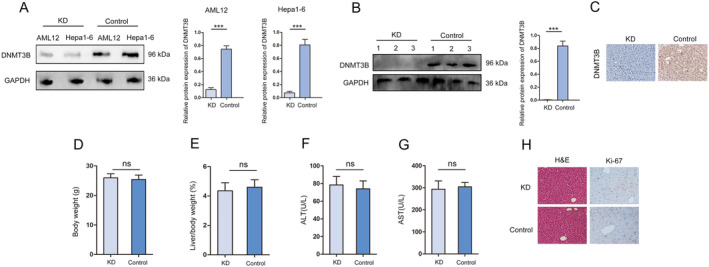 FIGURE 3
FIGURE 3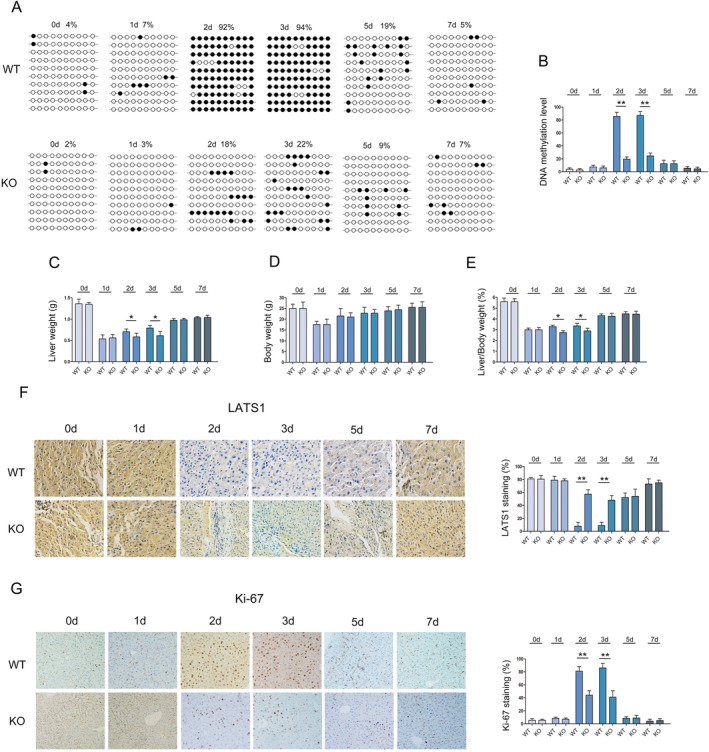 FIGURE 4
FIGURE 4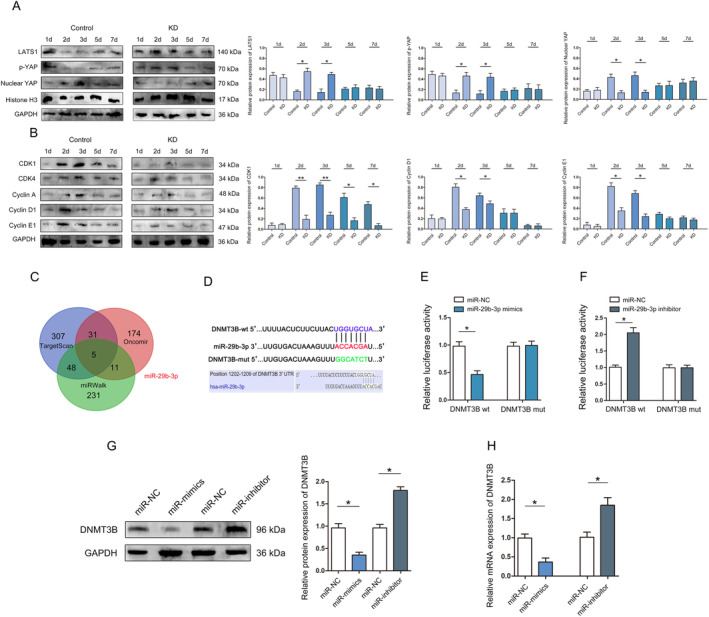 FIGURE 5
FIGURE 5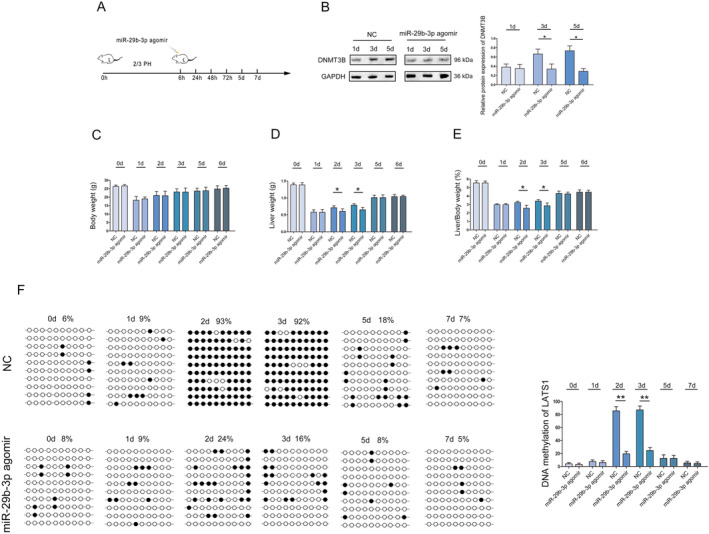 FIGURE 6
FIGURE 6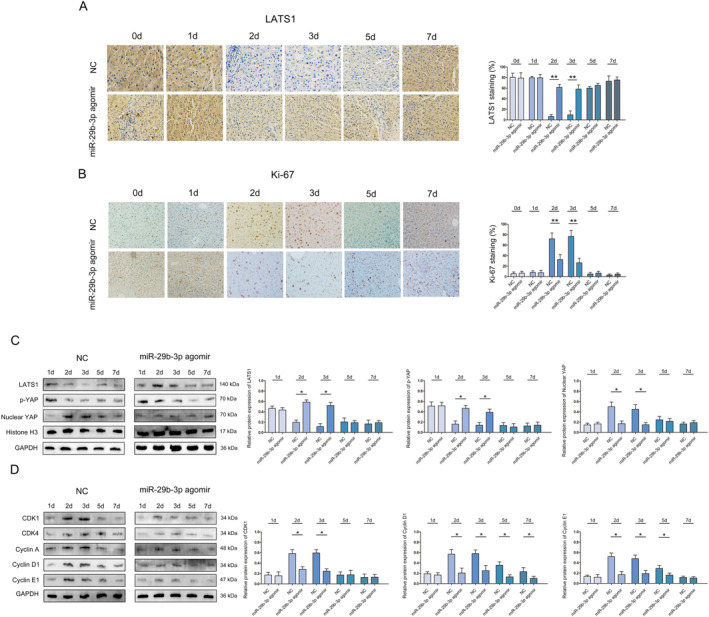 FIGURE 7
FIGURE 7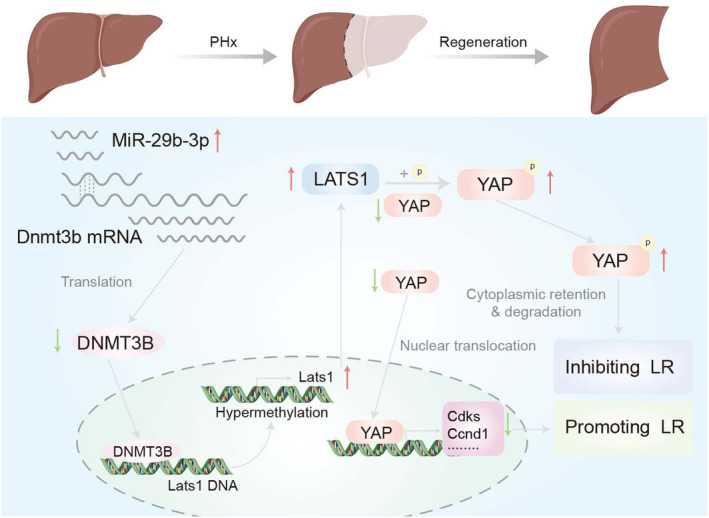 FIGURE 8
FIGURE 8- —Guizhou Provincial Health Commission Fund
- —National Natural Science Foundation of China 10.13039/501100001809
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsHippo pathway signaling and YAP/TAZ · Lipid metabolism and biosynthesis · Wnt/β-catenin signaling in development and cancer
Introduction
1
The liver is one of the few parenchymal organs with strong regenerative ability [1]. Hepatocytes account for approximately 65% of the total number of liver cells and are the ‘main force’ of liver regeneration (LR). Partial hepatectomy (PH) in rats or mice is the most commonly used model for studying liver regeneration [1, 2]. After partial liver lobectomy, the remaining liver lobes rapidly expand through a combination of liver cell hypertrophy and proliferation, compensating and restoring partial liver function [2]. Although the mechanism of liver regeneration has not been fully elucidated, it is clear that the Hippo signalling pathway is one of most important signalling pathways to regulate liver regeneration [3]. During liver regeneration, the cell proliferation and expression of apoptotic factors in the remaining liver cells are inevitably altered [4]. Hippo signalling pathway acts on the downstream effective factor Yes associated protein (YAP) through a series of kinase phosphorylation reactions, leading to its binding with cytoskeletal proteins and retention in the cytoplasm, thereby reducing its nuclear activity [5], through which the Hippo pathway controls organ size and volume by regulating cell proliferation and apoptosis [6, 7, 8, 9]. The regulatory role of Hippo pathway in liver regeneration has been confirmed in mouse models. Specific overexpression of YAP in the liver leads to liver enlargement, but once YAP overexpression is halted, liver size can return to normal [10, 11].
Large tumour suppressor kinase 1 (LATS1) plays an important role in the phosphorylation modification of YAP [12]. Konishi et al. found that LATS1 is inactivated during the initiation of liver regeneration, leading to non‐phosphorylation activation of YAP, upregulating the expression of connective tissue growth factor and apoptosis inhibitory protein, and promoting liver regeneration [13]. Loforese et al. found that during the termination stage of liver regeneration, LATS1 can induce phosphorylation of YAP, leading to a significant decrease in the expression level of cell proliferation factors regulated by YAP transcription [14]. The above suggests that the phosphorylation of YAP is crucial for regulating liver regeneration, while LATS1 plays an important role in the phosphorylation process of YAP.
DNA methylation is a common epigenetic modification, which is catalysed by methyltransferase to covalently bond to a methyl group at the cytosine (C) 5′ carbon position of CpG island dinucleotide in the genome, resulting in a conformational change in DNA and affecting its function [15, 16]. When DNA methylation occurs in the promoter region of a gene, it usually inhibits gene transcription [17]. During liver regeneration, the DNA methylation levels in the promoter regions of c‐myc, p53 and H‐RAS decrease, and protein expression significantly increases, promoting mitosis and proliferation of liver cells [18]. Transcriptomic analysis of PH in mice revealed that DNA hypomethylation caused by UHRF1 deficiency in liver cells can lead to sustained activation of genes that promote liver regeneration and enhance liver regeneration ability [19]. It can be seen that DNA methylation plays an important regulatory role in liver regeneration. In addition to classical protein phosphorylation modification, DNA methylation also plays a significant regulatory role in the protein expression of LATS1 [20]. A Previous study has confirmed that DNA methylation in the promoter region of LATS1 can inhibit its protein expression, leading to up regulation of cell proliferation and anti‐apoptotic factor expression, and promoting the proliferation and metastasis of breast cancer cells [21]. In liver cancer, DNA methylation in the LATS1 promoter region leads to dysfunction of the Hippo signalling pathway, inducing excessive production of monoacylglycerol lipase and promoting malignant proliferation of liver cancer cells [22]. However, the regulation of hepatocyte proliferation by DNA methylation and Hippo signalling pathway during LR and the molecular mechanism of these regulatory effects remains unclear.
In this study, we established liver‐specific DNMT3B‐deleted mice and found that DNMT3B regulates the expression level of LATS1 through DNA methylation after PH; furthermore, the results demonstrated that miR‐29b‐3p significantly regulates the expression of DNMT3B and alters LATS1 expression to inactivate the Hippo signalling pathway. In general, miR‐29b‐3p/DNMT3B regulatory axis influences LATS1 expression through DNA methylation, and regulates liver regeneration through the Hippo signalling pathway.
Materials and Methods
2
Mice and PH Model
2.1
The classical PH models was established as previously described [23]. The tissue and blood samples of each mouse were collected at indicated timepoints for histology and biochemical analysis, respectively. Nucleic acids and proteins extracted from liver tissues were further examination. Liver specifically DNMT3B knockdown mice were generated by AAV8‐shRNA DNMT3B. Male mice aged 8 ~ 10 weeks were subjected to hepatectomy. 5azaC was dissolved in sterile PBS and intraperitoneally injected into mice at 0.25 mg/kg once a day for 5 days starting 2 days before liver resection.
Western Blot
2.2
Extracting total protein from liver samples using RIPA lysis buffer (Beyotime). 25 μg of protein/well were electrophoresed by 10% sodium dodecyl sulphate polyacrylamide gel electrophoresis (SDS‐PAGE). PVDF membranes were blocked with 5% non‐fat powdered milk at room temperature for 1 h and incubated with primary antibody protein.
LATS1 (1:2000; Abcam), DNMT3B (1:3000; Abcam), p‐YAP, (1:1000; Abcam), YAP, (1:5000; Abcam), CDK1 (1:10000; Abcam), CDK4 (1:2000; Abcam), Cyclin A, (1:2000; Abcam), CyclinD1, (1:10000; Abcam), Cyclin E1, (1:1000; Abcam), and GAPDH antibody (1:5000, Abcam) at 4°C overnight. Chemiluminescence (ECL) (Thermo Fisher) was used to detect the expression of the target proteins.
Quantitative Real‐Time Polymerase Chain Reaction (RT‐qPCR)
2.3
Total RNAwas extracted by TRIzol reagent (TaKaRa) and reversely transcribed into cDNA using PrimeScript RT Reagent (TaKaRa). RT‐qPCR was performed using SYBR Premix Ex Taq II (TaKaRa). miR‐29b‐3p expression was determined by a TaqMan MicroRNA Assay kit (Applied Biosystems; Thermo Fisher Scientific Inc). U6 and GAPDH were used as internal references. The sequences used in this study are listed in Table S1.
Histology and Immunohistochemical Staining
2.4
Haematoxylin‐eosin (HE) staining and immunohistochemical staining to assess the pathological changes and the expression of LATS1 and Ki‐67 in mouse livers. The slices were incubated with LATS1(1:500; Abcam), and Ki‐67 (1:500; Abcam) overnight at 4°C, followed by biotinylated secondary antibody at 37°C for 1 h. Colour development was carried out with DAB (3,3‐diaminobenzidine). The slides were counterstained with 1% Mayer's haematoxylin. LATS1 and Ki‐67 immunostaining were scored and examined by two independent assessors.
Reintroduction of miR‐29b‐3p Agomir in Mice Liver
2.5
miR‐29b‐3p agomir (5 nmol/mouse) or an miRNA negative control (Ribo‐bio) was injected via tail vein into C57BL/6 mice at 1 day before 2/3 PH, and liver samples were collected at the designed experimental time points [23].
Bisulfite Sanger Sequencing (BSP)
2.6
For DNA methylaiton examination, 600 ng of genomic DNA extracted from liver samples was bisulfite converted using a MethylCode Bisulfite Conversion Kit (Applied Biosystems, USA). LATS1 promoter was amplified by PCR with Taq DNA Polymerase (Invitrogen, USA). The primer sequence was designed using Methyl Primer Express Software v1.0 (Applied Biosystems, USA). The PCR products were electrophoresed, purified using Spin‐X tubes, and then cloned into the pUC‐T vector (both from CWBiotech, Beijing, China). Ten single products were sequenced for each sample.
miRNA Target Prediction
2.7
Three prediction databases, including TargetScan (http://www.targetscan.org), Oncomir (http://www.oncomir.org/) and miRWalk (http://mirwalk.umm.uni‐heidelberg.de/) were used to predict miRNAs targeting DNMT3B.
Luciferase Reporter Assay
2.8
Wild‐type DNMT3B‐3′ UTR (wt) and mutant DNMT3B‐3′ UTR (mut) containing the putative binding site of miR‐29b‐3p were amplified by GenePharma and cloned into the firefly luciferase‐expressing pMIR‐REPORT vector (Obio Technology). Luciferase reporter vector and miR‐29b‐3p mimic, miR‐29b‐3p inhibitor and miR‐NC were transiently co‐transfected using Lipofectamine 3000. Luciferase assays were performed using the Luciferase Reporter Assay System (GloMax).
Statistical Analysis
2.9
All data are presented as the mean ± standard deviation (SD). SPSS 22.0 software and GraphPad Prism version were used for statistical analysis. Statistical differences were analysed by Student's t‐test, while the significance of differences between multiple groups was determined by one‐way analysis of variance, followed by the Newman–Keuls test, and repeated measures analysis of variance. p < 0.05 indicated statistically significant.
Results
3
LATS1 Regulates the Process of Liver Regeneration
3.1
We performed classic 2/3 PH in mice to determine the role of Hippo in liver regeneration. Western blot was used to analyse the expression of LATS1, p‐YAP and Nuclear YAP in the liver, and the results show that the expression of LATS1 and p‐YAP dynamic change (first decreased and then increased) during liver regeneration, while the expression of Nuclear YAP dynamic change (first increased and then decreased) (Figure 1A–C). In addition, cell proliferation‐related genes MKi‐67, Cyclin D1 and CDK4 show a reverse trend compared with LATS1 during liver regeneration (Figure 1D–F). IHC further confirmed the above results (Figure 1G,H). Moreover, statistical analysis showed that LATS1 was negatively correlated with Ki‐67 expression during the critical period of LR (Figure 1I).
*The expression of Hippo signalling pathway (LATS1, p‐YAP and Nuclear YAP) in liver regeneration. (A) Western blot was used to test the expression of LATS1, p‐YAP and Nuclear YAP in liver regeneration. (B) Statistical analysis of the expression of LATS1, p‐YAP and Nuclear YAP. (C) RT‐qPCR was used to analyse the expression of LATS1 in liver regeneration. (D) RT‐qPCR was used to analyse the expression of Ki‐67 in liver regeneration. (E) RT‐qPCR was used to analyse the expression of cyclin D1 in liver regeneration. (F) RT‐qPCR was used to analyse the expression of CKD4 in liver regeneration. (G) Immunohistochemistry was used to analyse the expression of LATS1 and Ki‐67 in liver regeneration, magnification X20. (H) Statistical analysis of the expression of LATS1 and Ki‐67 in liver regeneration. (I) Statistical correlation analysis of the expression relationship between LATS1 and Ki‐67. All data are represented as the mean ± SD, *p < 0.05, *p < 0.01.
DNA Methylation Regulates the Expression of LATS1 in Liver Regeneration
3.2
The above results indicate that there is a significant difference in the expression of LATS1 during liver regeneration, and epigenetics can regulate gene expression without changing the nucleotide sequence of the gene [24]. Our previous study has also shown that DNA methylation plays a crucial role in the expression of LATS1 genes [20]. During liver regeneration, BSP detection results revealed the degree of methylation of LATS1 was gradually increased (Figure 2A,B). Moreover, statistical analysis showed that the methylation degree of LATS1 was negatively correlated with the expression of LATS1 (Figure 2C). A Previous study has reported that demethylating drugs can regulate DNA methylation levels of genes in liver regeneration [24]. Furthermore, we added demethylation drugs (5azaC) to treat mice and performed PH [25]. After demethylation, the remnant liver was sampled at 0, 1, 2, 3, 5 and 7 day time points for detection. In the early stage of liver regeneration (3 and 4 day), the liver/body weight ratio was lower (Figure 2D–F). The results showed that the methylation of LATS1 was significantly reduced and protein expression increased during liver regeneration (Figure 2G–I). The expression of p‐YAP was increased and Nuclear YAP was decreased during liver regeneration (Figure 2J).
*Detection and quantification of DNA methylation of LATS1 in regenerated liver. (A) Bisulfite sequencing analysis was performed on LATS1 promoter methylation in regenerated liver. Black dots, methylated; white dots, unmethylated. (B) Statistical analysis of DNA methylation level of LATS1 in liver regeneration. (C) Statistical correlation analysis of the relationship between LATS1 expression and DNA methylation. (D) Statistical analysis of the effect of demethylation treatment on liver weight during liver regeneration. (E) Statistical analysis of the effect of demethylation treatment on body weight during liver regeneration. (F) Statistical analysis of the effect of demethylation treatment on liver/body weight ratio during liver regeneration. (G) Bisulfite sequencing analysis was performed on LATS1 promoter methylation after demethylation treatment in regenerated liver. Black dots, methylated; white dots, unmethylated. (H) Statistical analysis of the effect of demethylation treatment on the expression of LATS1. (I) RT‐qPCR was used to analyse the expression of LATS1 after demethylation treatment in regenerated liver. (J) Western blot was used to analyse the expression of LATS1, p‐YAP and Nuclear YAP after demethylation treatment in regenerated liver. All data are represented as the mean ± SD, *p < 0.05, *p < 0.01.
DNMT3B Promotes Changes in LATS1 Methylation During Liver Regeneration
3.3
Methyltransferase is a key enzyme for DNA methylation, including DNMT1(maintain methylation), DNMT2 (methylated tRNA), DNMT3A (de novo methylation), and DNMT3B (de novo methylation) [25]. Our previous research found that the CpG island in the LATS1 promoter region can be catalysed by DNMT3B to undergo DNA methylation and inhibit the protein expression of LATS1 in HCC [20]. In addition, we further detected the expression of DNA methylation enzymes DNMT1, DNMT3A, and DNMT3B in liver regeneration, The results indicate that the expression trend of DNMT3B in liver regeneration is consistent with the change trend of LATS1 DNA methylation (Figure S1) To explore whether the DNA methylation of LATS1 regulated by DNMT3B during liver regeneration, we specifically deleted DNMT3B with AAV‐8 (KD). Shown in Figure 3A is the specific knockdown of DNMT3B in the liver and HCC cells. Western blot and immunohistochemical results show that DNMT3B was efficiently ablated in KD mouse livers (Figure 3B,C). Moreover, body weight, liver/weight ratio, ALT and AST results showed no significant difference in KD and Control mice (Figure 3D–G). The HE results showed no significant damage existed in KD mice (Figure 3H). These results indicated that L depletion of DNMT3B did not cause obvious liver injury.
*The construction of hepatocyte‐specific DNMT3B‐deficient mice. (A) The specific knockdown of DNMT3B in the liver and HCC cells. (B) Western blot was used to analyse the expression of DNMT3B in KD and Control mice. (C) Immunohistochemistry was used to analyse the expression of DNMT3B in KD and Control mice, magnification X20. (D) Body weight ratio results from KD and Control mice. (E) Liver/body weight ratio results from KD and Control mice. (F) The serological detection of ALT. (G) The serological detection of AST. (H) HE staining to test the liver condition of KD and Control mice, magnification X20. All data are represented as the mean ± SD, **p < 0.001.
In the early stage of liver regeneration (2 and 3 days), the liver weight and liver/body weight ratio of KD mice were lower than that of Control mice (Figure 4A–C). In this period, DNA methylation of LATS1 in KD mice were lower than that of Control mice (Figure 4D,E). Additionally, the expression of LATS1 in KD mice was significantly higher than that in Control mice (Figure 4F). Moreover, the expression of Ki‐67 in KD mice was significantly lower than that in Control mice (Figure 4G).
*Detection of DNA methylation and protein of LATS1 in DNMT3B KD mice. (A) Statistical analysis of the effect of demethylation treatment on body weight during liver regeneration. (B) Statistical analysis of the effect of demethylation treatment on liver weight during liver regeneration. (C) Statistical analysis of the effect of demethylation treatment on liver/body weight ratio during liver regeneration. (D) Bisulfite sequencing analysis was performed on LATS1 promoter methylation in regenerated liver. Black dots, methylated; white dots, unmethylated. (E) Statistical analysis of DNA methylation of LATS1 in liver regeneration. (F) Immunohistochemistry was used to analyse the expression of LATS1 in KD and WT mice, magnification X20. (G) Immunohistochemistry was used to analyse the expression of Ki‐67 in KD and Control mice, magnification X20. *p < 0.05, *p < 0.01.
Western blot results show that LATS1 and p‐YAP were highly expressed in KD mouse livers, while Nuclear YAP was lower after 70% PH (Figure 5A). In addition, we found that CDK1, CDK4, cyclin A, cyclin D1 and cyclin E1 were all lower in KD compared to Control mice at early stage of liver regeneration (Figure 5B).
*Detection and quantification of miR‐29b‐3p, cyclins, CDKs and key regulator proteins of Hippo signalling pathway in regenerated liver of KD and Control mice. (A) Western blot was used to analyse the expression of LATS1, p‐YAP and Nuclear YAP in liver regeneration. (B) CDK1, CDK4, cyclin A, cyclin D1 and cyclin E1 at indicated time points in liver regeneration. (C) Venn diagram displaying DNMT3B target miRNAs. (D) The binding sites and corresponding mutant sites of miR‐29b‐3p and DNMT3B 3′ UTR. (E) A dual luciferase activity assay was performed by cotransfection of luciferase reporter containing DNMT3B 3′ UTR or the mutant reporter with miR‐29b‐3p mimics. (F) A dual luciferase activity assay was performed by cotransfection of luciferase reporter containing DNMT3B 3′ UTR or the mutant reporter with miR‐29b‐3p inhibitor. (G) Protein expression of DNMT3B detected by western blotting. (H) The expression of DNMT3B detected by RT‐qPCR. *p < 0.05, *p < 0.01.
miR‐29b‐3p Regulates DNMT3B and Affects the DNA Methylation Level of LATS1 in Liver Regeneration
3.4
A large number of studies have shown that miRNA not only plays an important role in liver tumours, but also plays a broad regulatory role in liver regeneration [26]. TargetScan (http://www.targetscan.org), Oncomir (http://www.oncomir.org/) and miRWalk (http://mirwalk.umm.uni‐heidelberg.de/) were used to analyse upstream regulatory miRNAs of DNMT3B. The results show that miR‐29b‐3p were potential target gene of DNMT3B (Figure 5C). We detected the expression of miR‐29b‐3p during liver regeneration, and the results showed a significant correlation between miR‐29b‐3p and DNMT3B expression (Figure S1). Binding site analysis found that miR‐29b‐3p contained a potential binding site in DNMT3B (Figure 5D). Co‐transfection of miR‐29b‐3p mimics significantly decreased luciferase activity in cells transfected with wt DNMT3B 3′‐UTR (Figure 5E), and co‐transfection of miR‐29b‐3p inhibitor significantly increased luciferase activity in cells transfected with wt DNMT3B 3′‐UTR (Figure 5F). In addition, western blot and qRT–PCR assays revealed that miR‐29b‐3p overexpression significantly reduced DNMT3B protein and mRNA levels (Figure 5G,H). These results suggest that miR‐29b‐3p directly targets and downregulates DNMT3B.
Furthermore, we injected mice with the miR‐29b‐3p agomir via tail vein at 6 h after 2/3 PH, and the remnant liver was sampled at 0, 24, 48, 72 h, 5 and 7 day time points for detection (Figure 6A). Western blot results showed that DNMT3B expression in miR‐29b‐3p agomir mice were lower than that of NC mice (Figure 6B). In the early stage of liver regeneration (2 and 3 days), the liver weight and liver/body weight ratio of miR‐29b‐3p agomir mice were lower than that of NC mice (Figure 6C–E).
*miR‐29b‐3p regulates DNMT3B and affects the DNA methylation of LATS1 in liver regeneration. (A) Schematic diagram for tail vein injection with miR‐29b‐5p agomir. (B) Western blot was used to analyse the expression of DNMT3B in liver regeneration. (C) Statistical analysis of the effect of demethylation treatment on body weight during liver regeneration. (D) Statistical analysis of the effect of demethylation treatment on liver weight during liver regeneration. (E) Statistical analysis of the effect of demethylation treatment on liver/ body weight ratio during liver regeneration. (F) Bisulfite sequencing analysis was performed on LATS1 promoter methylation in regenerated liver. Black dots, methylated; white dots, unmethylated. *p < 0.05, *p < 0.01.
miR‐29b‐3p/DNM3B Regulatory Axis Regulates the Role of Hippo in Liver Regeneration Through LATS1
3.5
After increasing the expression of miR‐29b‐3p, the methylation of LATS1 was reduced (Figure 6F). In addition, at 2 and 3 days after PH, the expression of LATS1 in miR‐29b‐3p agomir mice was significantly higher than that in NC mice (Figure 7A). Moreover, the expression of Ki‐67 in miR‐29b‐3p agomir mice was significantly lower than that in NC mice (Figure 7B).
*miR‐29b‐3p influences the role of Hippo signalling pathway in liver regeneration through LATS1. (A) Immunohistochemistry was used to analyse the expression of LATS1 in liver regeneration, magnification X20. (B) Immunohistochemistry was used to analyse the expression of Ki‐67 in liver regeneration, magnification X20. (C) Western blot was used to analyse the expression of LATS1, p‐YAP and Nuclear YAP in liver regeneration. (D) CDK1, CDK4, cyclin A, cyclin D1 and cyclin E1 at indicated time points in liver regeneration. *p < 0.05, *p < 0.01.
Western blot to detect the expression of LATS1, p‐YAP and Nuclear YAP in the process of liver regeneration at 1, 2, 3, 5 and 7 days. The results show that LATS1 and p‐YAP were highly expressed in miR‐29b‐3p agomir mouse livers, while YAP was lower after 70% PH (Figure 7C). In addition, we found that CDK1, CDK4, cyclin A, cyclin D1 and cyclin E1 were all lower in miR‐29b‐3p agomir mice compared to NC mice at early stage of liver regeneration (2 and 3 days) (Figure 7D). Overall, these results demonstrated that miR‐29b‐3p reduces mice liver regeneration after PHx by down‐regulating the expression of DNMT3B and regulate the DNA methylation of LATS1 (Figure 8).
Schematic representation of miR‐29b‐3p reduces mice liver regeneration after PHx by down‐regulating the expression of DNMT3B and regulate the DNA methylation level of LATS1 through DNMT3B.
Discussion
4
The Hippo signalling pathway is composed of a series of conserved kinases that regulate cell proliferation, apoptosis, stem cell self‐renewal and participate in multiple biological functions such as tissue development, tissue homeostasis maintenance and regeneration repair in various organisms [27, 28]. When Hippo pathway is activated, the activity of YAP/TAZ is inhibited through LATS1/2 mediated phosphorylation [29]. When Hippo pathway is inactivated, the dephosphorylated YAP/TAZ translocation enters the nucleus and binds to the transcription factor TEAD1‐4 to induce gene expression [30]. Hippo signalling pathway plays an important role in liver regeneration: on the one hand, it activates cell proliferation and the expression of anti‐apoptotic factors in the initiation and maintenance of liver regeneration to promote liver regeneration; On the other hand, reducing the expression of cell proliferation and anti‐apoptotic factors during the termination stage of liver regeneration inhibits liver regeneration [31]. Our study found that LATS1, a key protein of Hippo signalling pathway, CpG island in LATS1 promoter region can undergo DNA methylation changes (the degree of DNA methylation first increases and then gradually decreases) during liver regeneration, and significantly regulates its protein expression. Dynamic changes in cell proliferation and anti‐apoptotic factor expression regulated by YAP transcription (initially increasing and then gradually decreasing).
Methyltransferase is a key enzyme involved in DNA methylation of genes [32]. There are currently four known human methyltransferases: DNMT1, DNMT2, DNMT3A, and DNMT3B, which play important roles in de novo methylation and retention methylation processes, respectively [33]. Research has shown that DNMT3A is consistently expressed at low levels at different stages of the cell cycle, while DNMT2 is mainly involved in tRNA methylation modification rather than DNA methylation. However, DNMT1 and DNMT3B are induced to be highly expressed in the S phase, providing methylation modification guarantees for DNA replication, transcription, and protein synthesis [34]. Research has shown that DNMT3A is consistently expressed at low levels at different stages of the cell cycle, while DNMT1 and DNMT3B are induced to be highly expressed in the S phase, providing methylation modifications for DNA replication, transcription, and protein synthesis after liver injury, the number of hepatocytes decreases sharply, and various feedback signals stimulate the proliferation of hepatocytes in the G0 stage, and residual hepatocytes change from basically non‐growth state to rapid growth state through cell proliferation, so as to compensate for the loss and damage of liver tissue and restore the physiological function of the liver [35]. Research has shown that DNMT3B is highly expressed in the S phase of cell proliferation, providing methylation modification guarantees for DNA replication, transcription, and protein synthesis [36]. Liver regeneration is a process of liver cell proliferation and differentiation, where gene sequences undergo de novo methylation to produce new DNA methylation modifications, and DNMT3B is the key methyltransferase for de novo methylation [37, 38]. In our existing studies, we found that the expression of DNMT3B changed dynamically during liver regeneration (increased and then decreased gradually), and DNMT3B could significantly regulate the DNA methylation and protein expression of LATS1.
microRNAs (miRNAs) exert their functions by regulating the expression of specific target genes and are related to various pathophysiological activities of liver [39]. Some miRNAs have also been shown to be involved in the regulation of liver regeneration [40]. During the initiation stage of liver regeneration, the expression of miR‐21 is significantly upregulated at 1, 6, 12, 24 and 48 h, and reaches its peak at 12 and 24 h [41]. At the same time, it can promote the translation of cyclin D1 and initiate liver cell proliferation [41]. The expression of miR‐127 is significantly reduced during the initiation stage of liver regeneration [42]. Mechanism studies have shown that miR‐127 can promote the expression of Bcl‐6 and promote liver cell proliferation [42]. During the phase of liver regeneration and proliferation, miR‐221 regulates liver cell proliferation by influencing aromatic hydrocarbon nuclear transferors and subsequently regulating the expression of p27.^43^ During the termination stage of liver regeneration, miR‐23b inhibits the expression of TGF‐b1 through Smad3, and promotes the expression of TGF‐β, activing A and inhibits the proliferation of liver cells.^44^ Our results are also consistent with previous studies. Furthermore, we found that miR‐29b‐3p regulates the DNA methylation modification of LATS1 by regulating DNMT3B, inhibits the expression of LATS1, and dynamically regulates the process of liver regeneration.
In summary, this study provides evidence that miR‐29b‐3p reduces mice liver regeneration after PHx by down‐regulating the expression of DNMT3B and regulate the DNA methylation of LATS1 through DNMT3B, consequently leads to affecting the liver regeneration process by regulating the function of Hippo signalling pathway.
Author Contributions
Yinwen Zhou: conceptualization (equal), data curation (equal), methodology (equal), writing – original draft (equal), writing – review and editing (equal). Hao Wu: conceptualization (equal), formal analysis (equal), funding acquisition (equal), methodology (equal), writing – original draft (equal), writing – review and editing (equal). Qiu Wang: conceptualization (equal), investigation (equal), project administration (equal), resources (equal), software (equal), supervision (equal), validation (equal). Bo Ma: formal analysis (equal), funding acquisition (equal), investigation (equal), methodology (equal), project administration (equal), resources (equal), software (equal), supervision (equal), validation (equal), visualization (equal). Jiulong Sun: funding acquisition (equal), project administration (equal), resources (equal), software (equal), validation (equal), visualization (equal). Guoliang Wang: conceptualization (equal), funding acquisition (equal), writing – original draft (equal), writing – review and editing (equal).
Ethics Statement
The Animal Care and Use Committee of Guizhou Provincial People's Hospital granted approval for all experiments involving animals.
Consent
All authors consent to publication.
Conflicts of Interest
The authors declare no conflicts of interest.
Supporting information
Figure S1. Western blot was used to analyse the expression of DNMT1, DNMT3A and DNMT3B in liver regeneration. **p < 0.01.
Figure S2. RT‐qPCR was used to analyse the expression of miR‐29b‐3p in liver regeneration.
Table S1. The sequences of RT‐qPCR.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1G. K. Michalopoulos and B. Bhushan , “Liver Regeneration: Biological and Pathological Mechanisms and Implications,” Nature Reviews. Gastroenterology & Hepatology 18, no. 1 (2021): 40–55.32764740 10.1038/s 41575-020-0342-4 · doi ↗ · pubmed ↗
- 2S. A. Mao , J. M. Glorioso , and S. L. Nyberg , “Liver Regeneration,” Translational Research 163, no. 4 (2014): 352–362.24495569 10.1016/j.trsl.2014.01.005PMC 3976740 · doi ↗ · pubmed ↗
- 3J. O. Russell and F. D. Camargo , “Hippo Signalling in the Liver: Role in Development, Regeneration and Disease,” Nature Reviews. Gastroenterology & Hepatology 19, no. 5 (2022): 297–312.35064256 10.1038/s 41575-021-00571-w PMC 9199961 · doi ↗ · pubmed ↗
- 4J. L. Duan , B. Ruan , P. Song , et al., “Shear Stress‐Induced Cellular Senescence Blunts Liver Regeneration Through Notch‐Sirtuin 1‐P 21/P 16 Axis,” Hepatology 75, no. 3 (2022): 584–599.34687050 10.1002/hep.32209 · doi ↗ · pubmed ↗
- 5S. Ma , Z. Meng , R. Chen , and K. L. Guan , “The Hippo Pathway: Biology and Pathophysiology,” Annual Review of Biochemistry 88 (2019): 577–604.10.1146/annurev-biochem-013118-11182930566373 · doi ↗ · pubmed ↗
- 6Z. Meng , T. Moroishi , and K. L. Guan , “Mechanisms of Hippo Pathway Regulation,” Genes & Development 30, no. 1 (2016): 1–17.26728553 10.1101/gad.274027.115PMC 4701972 · doi ↗ · pubmed ↗
- 7Z. Wu and K. L. Guan , “Hippo Signaling in Embryogenesis and Development,” Trends in Biochemical Sciences 46, no. 1 (2021): 51–63.32928629 10.1016/j.tibs.2020.08.008PMC 7749079 · doi ↗ · pubmed ↗
- 8I. M. Moya and G. Halder , “Hippo‐YAP/TAZ Signalling in Organ Regeneration and Regenerative Medicine,” Nature Reviews. Molecular Cell Biology 20, no. 4 (2019): 211–226.30546055 10.1038/s 41580-018-0086-y · doi ↗ · pubmed ↗