Sensory Reconstruction of the Fossil Lorisid Mioeuoticus: Systematic and Evolutionary Implications
Holly E. Anderson, Adam Lis, Ingrid Lundeen, Mary T. Silcox, Sergi López-Torres

TL;DR
This study reconstructs the sensory anatomy of a fossil lorisid, Mioeuoticus, to understand its behavior and evolutionary position.
Contribution
The study provides new insights into the sensory evolution and phylogenetic placement of Mioeuoticus within lorisids.
Findings
Mioeuoticus exhibited modern lorisid behaviors like slow locomotion and nocturnal activity.
Its nasal turbinal arrangement suggests a basal position within Lorisidae or Lorisoidea.
Olfactory abilities of Mioeuoticus align with those of modern lorisids.
Abstract
We reconstructed the internal sensory system anatomy of the lorisid, Mioeuoticus shipmani (KNM-RU 2052), from the early Miocene of Rusinga Island, Kenya. Results suggest that Mioeuoticus developed typical modern lorisid behaviour (i.e., slow locomotion, nocturnal activity pattern) and olfactory abilities consistent with modern representatives. The fossil record of lorises and pottos (family Lorisidae) potentially dates back to the late Oligocene of Namibia, but a later moderate diversification of this family occurred during the Miocene of Africa and Asia. In the African Miocene, the family Lorisidae is represented solely by one genus: Mioeuoticus. The phyletic position of Mioeuoticus has been a source of debate, as it has been suggested to belong to either the stem of the family Lorisidae or to be further nested within lorisids, as a sister to the African potto clade (subfamily…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
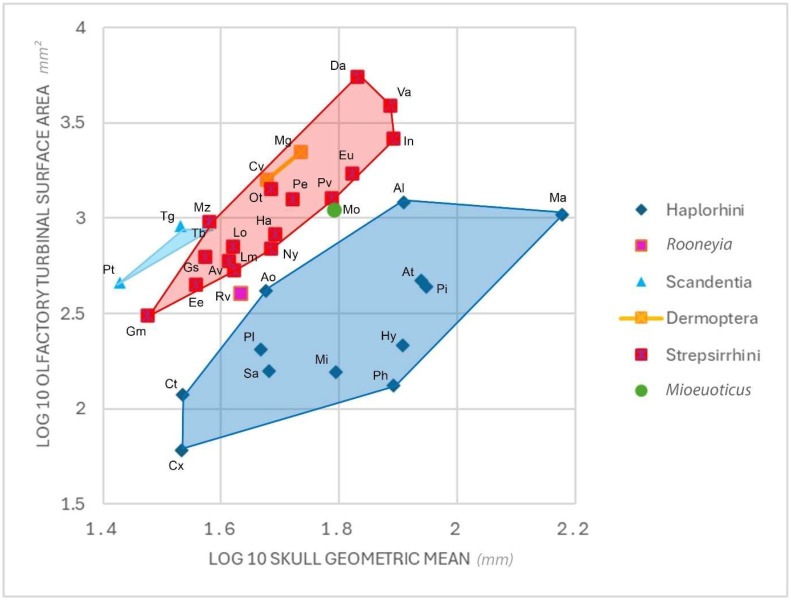 Figure 9
Figure 9- —NSERC Discovery
- —National Science Centre (Cracow, Poland)
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsPrimate Behavior and Ecology · Fish biology, ecology, and behavior · Ichthyology and Marine Biology
1. Introduction
Lorises and pottos (family Lorisidae) are nocturnal strepsirrhine primates that live in equatorial Africa, and southern and southeastern Asia. Compared to other primate families, lorisids are not a particularly speciose clade [1] but became highly specialised to a nocturnal, arboreal lifestyle and insectivorous dietary behaviours, often showing adaptations for gum extraction [2,3,4]. There has been extensive research into the ecology, behaviours, and evolutionary relationships of the modern lorisoids. The paucity of these animals in the fossil record means that our understanding of lorisid phylogeny, systematics, reconstructed behaviour, and functional anatomy in these fossil relatives is comparatively scarce and major aspects remain unresolved [1]. However, this is not to say there have not been significant discoveries from the available fossil specimens, which have significantly advanced our knowledge of lorisoid evolutionary histories, such as the potential origins of lorisoids [5] and lorisids [6], certain aspects of their locomotor ecology [7], and identification of a greater diversity of dietary behaviours in extinct forms [8,9,10,11,12]. The fossil discoveries that have been made clarify the interrelationships of extant lorisid genera and reveal seemingly good convergence between fossils and molecular divergence date estimates in certain parts of the phylogenetic tree [1].
Despite these advances, many questions remain as to the reconstructed behaviour in fossil lorisid forms. Whereas most fossil lorisid material comprises dentognathic or, occasionally, postcranial material [13,14], an almost complete cranium is known from the early Miocene of Rusinga Island, Kenya, belonging to Mioeuoticus shipmani [15]. The phylogenetic position of M. shipmani is currently uncertain, with Walker et al. [7] placing it as a sister group to all African lorisids (i.e., subfamily Perodicticinae), while Harrison [14] argued that the genus Mioeuoticus belongs to a more basal lineage at the stem of lorisids. In either case, Mioeuoticus represents an early member of the family Lorisidae and understanding its behaviour provides a deep evolutionary perspective in the context of the clade. The reconstructed behaviour of M. shipmani remains largely unexplored, but it is not completely unknown. Walker et al. [7] found that the semicircular canals (SCCs) in M. shipmani had a similar relative size to those seen in modern slow lorises (genus Nycticebus). The SCCs provide information on the body’s position in space, contributing in particular to the stabilisation of gaze in locomotion [16]. Among modern primates, slower-moving species tend to have smaller SCCs for a given body mass compared to faster-moving taxa [16]. The similarities between Mioeuoticus and Nycticebus found by Walker et al. [7] suggest, therefore, that Mioeuoticus was most likely a slow-moving arboreal quadruped, in a similar fashion to modern-day lorisids. However, besides the reconstructed locomotor behaviour, nothing is known about other aspects of its behaviour and, in particular, its sensory capabilities.
The main aim of this project is to reconstruct the sensory capacities of the early Miocene lorisid Mioeuoticus. The relative timing of the evolution of these characteristics may have important implications for the phyletic position of the early Miocene lorisid Mioeuoticus.
1.1. Vision
The optic foramen quotient (OFQ) value calculated for any given primate acts as a proxy for the degree of retinal summation, which is a quantification of the number of photoreceptor cells that converge on bipolar cells in the eye to transmit signals to ganglion cells for transfer to the brain for interpretation and response [17]. This interpretation and response deduces how to interact with and adapt to various life-preserving factors, including predators, food, and long-distance conspecific communication [18,19,20]. Thus, the OFQ gives a solid proxy for the degree of visual acuity in a primate; a low OFQ means high central retinal summation and a high OFQ means low central retinal summation [21].
The dimensions of the intra-orbital surface can be directly related to the size of the optic nerve in primates [22]. Direct comparisons with optic nerve measurements in extant primates with known visual capacities can provide a basis to hypothesise the visual acuity of fossil primates [21]. The left intra-orbital surface of the Mioeuoticus cranium volume rendering has been significantly taphonomically distorted; however, the right intra-orbital surface appears to be well preserved with little to no apparent damage.
1.2. Olfaction
To ensure survival, mammals rely strongly on their sense of smell to convey critical information about food seeking, mate selection, hazard detection, and behavioural immune responses [23,24,25,26,27,28,29]. The olfactory system is structured as long, scrolled, spongy networks of bone within the nasal cavity, known as turbinals, turbinates, or conchae. The turbinals are lined with a specialised tissue called olfactory epithelium, which contains receptor cells that bind with molecules that detect odorants that the brain interprets as smell [30,31]. Thousands of chemically diverse odour molecules and pheromones are detected and recognised by the olfactory receptor neurons in the olfactory epithelium, which are converted into electrical signals that are sent to the brain for interpretation and behavioural response [32,33]. Many primates also have a vomeronasal organ, with the purpose of detecting pheromones critical in perceiving and processing stimuli related to social and reproductive behaviours [34,35].
The proportions, number, and complexity of these structures give strong evidence of the olfactory landscape, behavioural ecology, and phylogenetic affinities of fossil primates [36,37,38,39,40]. These data may detect a system specialised to a specific range of odorants, which can strongly indicate the primate trends in the evolution of nasal morphometry and airflow [38,41,42]. Turbinals are delicate and rarely preserved in fossil specimens, limiting opportunities to make direct observations of the olfactory periphery in extinct primates. Fortunately, the turbinals of the Mioeuoticus specimen in this study are preserved with minimal damage or distortion.
1.3. Audition and Proprioception
Mammals depend on audition for vital interactions with their environments, including food detection and attainment [43,44], hazard detection and evasion [45,46], stabilising balance [16], and participating in communal exchanges governing territoriality or mating [47,48]. The auditory system comprises two fluid-filled receptor organs that form the membranous labyrinth, which is suspended within the petrous temporal bone inside a network of bony voids known as the bony labyrinth [49]. The bony labyrinth consists of semicircular canals (SCCs), the utricle and the sacculus, and the cochlea.
As sound waves enter the inner ear, the fluid in the utricle, sacculus, and cochlear receptor organs vibrate. The hairs in the cochlea have different lengths and vibrate at numerous sound frequencies. Stimulating these microscopic hairs generates electrical impulses and sends them to the brain for interpretation and behavioural response [7,50]. The semicircular canals regulate balance and sense head position to maintain stability despite motion and activity. The orientation, relative position, and size of the semicircular canals give strong evidence of the locomotor behaviour and shape concerning phylogenetic affinities of fossil primates [51,52,53,54,55,56,57]. Measurements of semicircular canals in extant mammals with known locomotor behaviours can provide a basis for direct comparisons with which to test hypotheses about locomotion in fossil primates [58,59]. The bony labyrinths of the Mioeuoticus specimen in this study are difficult to quantify due to poor CT resolution, but some diagnostic structures appear to be preserved with minimal damage or distortion.
1.4. Phylogenetic Implications
These data may provide critical insights as to whether Mioeuoticus are stem or crown members of Lorisidae, or possibly advanced stem, or very basal crown, lorisoids. To better understand the phylogenetic and ecological significance of these observations for Mioeuoticus, we provide a comparative framework with analyses of extant and extinct specimens in the associated literature for evaluating sensory character evolution to assess the relationships between fossil and living lorisids. Understanding how fossil lorisids relate to their living relatives is critical for explaining major trends in primate evolution. A better understanding of the relationships among extinct and extant lorisids may speak to the question of the biogeographic origin of this primate family and a broader understanding of the characteristic low diversity of modern lorisids.
1.5. Institutional Abbreviations
BAA—Department of Evolutionary Anthropology, Duke University, Durham, NC, USA; MCZ—Museum of Comparative Zoology, Harvard University, Cambridge, MA, USA; AMNH—American Natural History Museum, NY, USA; YPM—Yale Peabody Museum at Yale University, New Haven, CT, USA; KNM-RU—National Museums of Kenya, Rusinga Island collection, Nairobi, Kenya.
2. Methods
The cranium of Mioeuoticus shipmani (KNM-RU 2052; housed in the Palaeontological Collections of the National Museums of Kenya, Nairobi) had been previously scanned on-site using a portable micro-CT scanner by a team from Kyoto University. The resulting scan has 575 total slices with voxel dimensions of x = 0.055, y = 0.07, and z = 0.07, providing a 16-bit TIFF image stack data set for the cranium, and the slices were rendered as 3D volumes in Avizo 3D Pro 2023 (Visualization Sciences Group, Berlin, Germany). This three-dimensional rendering was used to reconstruct the internal anatomy of the sensory systems as digital endocasts. These structures of Mioeuoticus (as evident in Figure 1) were manually segmented in each slice using the brush tool.
2.1. Vision
The visual acuity of Mioeuoticus shipmani is analysed following the OFQ method and analysis of the relationship between orbit diameter and cranial length, and activity period, as described by Kirk and Kay [21]. The length and width of the orbital and optic foramen cavities in the three-dimensional volume rendering of the Mioeuoticus cranium are taken using the ruler tool on Avizo 3D (See Supplementary Table S1); thus, the area and diameter of these two regions can be calculated. The orbit diameter is 16.52 mm and the P-I length is 60.33 mm. When compared to the bivariate plot of ln mean orbit diameter and ln mean cranial length for 60 extant primate species in Figure 5B of Kay and Kirk [17] Mioeuoticus shipmani lies firmly in the group with nocturnal strepsirrhines.
The formula for the optic foramen area is then calculated as follows: A = π a b where A is the area of the orbital cavity, a is the major radius length, and b is the minor radius length of the optic foramen and orbital cavity. We then measure from the craniometric point, which is the most anterior point in the midline on the alveolar process of the maxilla, to the tip of the external occipital protuberance—known as the prosthion–inion (P-I) length. This calculation is used as a substitute and estimation for the total body size of this specimen.
These data are then added as variables into the equation by Kirk and Kay [21] to approximate optic foramen size relative to eye size, as follows:
Kirk and Kay [21] state that this equation increasingly overestimates eye size as the body size of a specimen increases, and thus to remove this bias, the OFI result for body size must be adjusted using the following equation:
We then measured the prosthion–inion (P-I) length, which runs from the craniometric point, which is the most anterior point in the midline on the alveolar process of the maxilla, to the tip of the external occipital protuberance (see Supplementary Table S1). P-I length is needed to calculate the expected OFI.
The expected OFI is calculated with two empirical regression formulas to express the OFI as a percentage of that which is expected for diurnal anthropoids (LnOFI = 3.39983 − 0.56624 × ln [P-I length]) and nocturnal strepsirrhines (LnOFI = 2.46913 − 0.58138 × ln [P-I length]) of similar P-I length. Therefore, we input the P-I length of the Mioeuoticus holotype into both of these equations and raise the result of these calculations to the power of e, where e ≈ 2.718, to determine the diurnal anthropoid regression (daOFQ) and nocturnal strepsirrhine regression (nsOFQ) for the specimen, as follows:
These data were then directly compared to the daOFQ and nsOFQ of extant specimens determined in Table 5 of Kirk and Kay [21] to assess visual acuity in Mioeuoticus (see Supplementary Table S2).
2.2. Olfaction
We provide a detailed description of the reconstructed Mioeuoticus olfactory turbinals, in which all nomenclature and definitions of turbinals follow Smith and Rossie [60]. We follow the convention of numbering ethmoturbinals using Roman numerals from anterior to posterior view. All descriptions of turbinal morphology follow an anterior-to-posterior sequence based on successive coronal CT slices. Terms used to describe turbinal shape in coronal cross-section follow Lundeen [41]. When possible, turbinals were segmented from the lateral wall of the nasal fossa at the point where the turbinal’s basal lamina make contact with the lateral wall. We also quantify the surface areas of both the olfactory turbinals and the maxilloturbinals of the Mioeuoticus specimen using the surface area tool in Avizo. The overall turbinal and maxilloturbinal data are both correlated with turbinal surface areas of a range of extant strepsirrhines and haplorhines calculated by Lundeen and Kirk [38] to hypothesise the olfactory abilities of Mioeuoticus compared to other similarly sized primates.
2.3. Audition
We provide detailed descriptions of the overall morphology of the bony labyrinth, focused mainly on the orientation, relative position, and size of the semicircular canals. The anterior, posterior, and lateral semicircular canals are described, respectively, as they appear in anterior-to-posterior view based on successive coronal CT slices. The angle between the three canal arcs is measured at the greatest width of the anterior and posterior semicircular canals on the transverse plane, following Spoor and Zonneveld [53].
The medial end of the anterior (superior) semicircular canal joins with the upper part of the posterior canal to form a bony limb, known as the common crus [61,62], whose length we additionally measure (CCL). CCL is measured from the opening of the vestibular aqueduct in the vestibular wall to the bifurcation point of the common crus—known as the crus commune apex—following inner ear morphological landmark locations defined in LeBrun [61]. The length of the common crus in the reconstructed Mioeuoticus inner ear is compared to the mean CCL in extant slow- and fast-moving primates (Pier et al. [62]: Table 4).
3. Results
The cranial measurements of the holotype of Mioeuoticus shipmani (KNM-RU 2052) are as follows: prosthion–inion length of 61.8 mm, cranial height of 26.3 mm, neurocranial width of 33.9 mm, and maxillary width of 30.5 mm. Cranial height is measured from the basion (midpoint of the anterior margin of the foramen magnum) to the highest point on the skull. Neurocranial width is measured across the most lateral aspect of each parietal (euryon to euryon). Maxillary width is measured across the most lateral aspect of each temporal bone.
3.1. Vision
Following the OFI and OFQ equations of Walker et al. [7], the daOFQ and nsOFQ were calculated for the holotype of Mioeuoticus shipmani.
3.2. Olfaction
Detailed morphological analyses and anatomical measurements for the individual turbinals in the Mioeuoticus olfactory reconstruction are presented below. These data are likely an underestimation as fragmentation of the turbinals reduces their overall surface area to less than that of the original total turbinal structure and the turbinal surface area present in the living Mioeuoticus nasal cavities, though still provide a novel physiological understanding of this previously unstudied olfactory system. The internal turbinal measurements are available in Table 1.
The cross-sectional morphology of each of the turbinals is described below from anterior to posterior on a coronal plane (Figure 2 and Figure 3), including anatomical details of the olfactory cavity, the relative position of each turbinal in the nasal fossa, and the nature of the turbinals’ contact with the fossa wall or other turbinals. It is recognised that debate still lies in whether to group the two closely associated anterior-most turbinal structures as a single ethmoturbinal (ethmoturbinal I) or divide them into two ethmoturbinals (ethmoturbinal I and II). Here, we take a conservative approach and divide these structures into two ethmoturbinals. In doing so, we form a comprehensive description of each structural subdivision that may better contribute to future evaluations of these characteristics within the appropriate comparative context. Turbinal names are abbreviated as follows (left nasal turbinals—uppercase abbreviations; right nasal turbinals—lowercase abbreviations); NT/nt—nasoturbinal, FT/ft—frontoturbinal, ET/et-I to -IV—ethmoturbinals 1 to 4, IT/it—interturbinal, MT/mt—maxilloturbinal.
The Mioeuoticus specimen has all the turbinals we may expect to find here and there is a symmetry between the left and right fossa, though these turbinals are broken and fragmented. In total, there is one pair of nasoturbinals, frontoturbinals, interturbinals, and maxilloturbinals, and three pairs of ethmoturbinals. The left and right turbinals are separated in the anterior portion of the olfactory cavity by the nasal septum. In the posterior of the olfactory cavity, a transverse septum separates a superior olfactory recess from an inferior nasopharyngeal meatus enclosing all of the turbinals.
There may also be a fourth pair of ethmoturbinals in the olfactory recess of this specimen, though this cannot be fully confirmed nor reconstructed due to the poor resolution of the scan (Figure 2). No defining structure can be seen, and so these turbinals are not described any further below.
3.2.1. Turbinals of the Left Nasal Cavity
The left nasal turbinals are well preserved overall, with all major turbinals mostly intact. The left nasal fossa contains one NT, one FT, three ETs, one IT, and one MT.
There is a minor mediolateral distortion of the NT, and ET-I, -II, and -III.
Maxilloturbinal (MT)—This turbinal is the most anterior of the assemblage, situated only 1.3 mm posterior to the piriform aperture, with a slight posterior sloping gradient on the sagittal plane. This turbinal’s anterior and inferior margins are preserved along the full length of the structure. This is a double-scrolled turbinal, with both the medially and laterally directed scrolls well preserved (as seen in slice 97 of the specimen CT scan; Figure 3A, Figure 4, and Figure 5C–H). It is difficult to assess whether the anterolateral wall of the turbinal ceases at the scrolled portion of the structure or has been destroyed and would extend to the full extent of the basal lamina.
Nasoturbinal (NT)—This turbinal is a single-scrolled structure that appears almost perfectly preserved (as seen in slices 137, 139, 146, and 156 of the specimen CT scan; Figure 3B–E, Figure 4, and Figure 5C–G). There is no apparent deformation, fragmentation, or destruction. The turbinal is orientated anteromedially in the olfactory cavity with a significant anteriorly sloping gradient, 15.4 mm posterior to the piriform aperture. The anterior-most margin of the turbinal is anterolateral to the scrolled midsection, which has a ‘teardrop’ shape in the cross-section. The turbinal has a single direct point of contact with the superior fossa wall running anteroposteriorly.
Ethmoturbinal I (ET-I)—This turbinal is positioned inferiorly to the NT and FT 13.4 mm to the piriform aperture, with a steep anterior gradient on the sagittal plane. It is on the same sagittal plane as the NT and inferolateral to the FT. Anteriorly, the ET is broken. Further towards the central structure, the ET forms a single bulla for the main portion of the turbinal, which detaches laterally and arcs into a scroll that reduces to a branching structure. This enclosed bulla decreases in volume posteriorly. Initially, the enclosed bulla has two points of contact with the lateral fossa wall. As the bulla detaches to a scroll structure, these connections merge into a singular point of contact (as seen in slices 139 and 146 of the specimen CT scan; Figure 3C,D, Figure 4 and Figure 5A–F).
Ethmoturbinal II (ET-II)—This turbinal is inferomedial to ET-I, situated 13.1 mm posterior to the piriform aperture, and orientated anteromedially in the olfactory cavity with a steep anterior gradient on the sagittal plane. Anteriorly, ET-II is a singular bulla that terminates at two points of contact with the inferior medial and lateral border of ET-I (as seen in slice 137 of the specimen CT scan; Figure 3B). The medial point of contact with ET-I disconnects posteriorly, forming an arcuate structure that extends horizontally across the nasal fossa and then curves upward to run parallel with the septal wall (as seen in slice 139 of the specimen CT scan; Figure 3C). It is unclear whether the terminal margin of the bulla is incomplete or the resolution of the scan is too poor to discern it. The connection with the superior fossa wall is lost towards the main portion of this turbinal (as seen in slices 146, 156, and 173 of the specimen CT scan; Figure 3D–F, Figure 4 and Figure 5A–F). Anteriorly, the turbinal is a branching structure that contact the superior and lateral fossa wall (as seen in slice 179 of the specimen CT scan; Figure 3G).
Frontoturbinal (FT)—This turbinal is located inferolateral to the ET-II, orientated anteromedially in the olfactory cavity with a steep anterior gradient on the sagittal plane. Situated 16.4 mm posterior to the piriform aperture, it has an arcuate morphology; a regular arc that increases in circumference from anterior to posterior (as seen in slices 146, 156, and 173 of the specimen CT scan; Figure 3D–F and Figure 5E–G). It cannot be determined whether this is due to damage and fragmentation of a scroll or if this is the original structure. The turbinal has a single direct point of contact with the lateral fossa wall running anteroposteriorly.
Ethmoturbinal III (ET-III)—Anteriorly, this turbinal is inferior to ET-II, located on the same sagittal plane in the anterior, at 13.1 mm posterior to the piriform aperture. Posteriorly, the turbinal is lateral to ET-II as it is orientated anteromedially in the olfactory cavity with a steep anterior gradient on the sagittal plane. It is difficult to discern whether this turbinal is bullar (via a superior and inferior contact with the lateral fossa wall) or arched (with a single point of contact with the fossa wall), as part of ET-III is not visible on the CT rendering. Posteriorly, this arch curves more steeply to form a single medial scroll with branches that contact the septal wall (as seen in slice 179 of the specimen CT scan; Figure 3G). More superiorly, a superior branching point of contact is also formed (as seen in slice 190 of the specimen CT scan; Figure 3H). The scroll gradually becomes less pronounced, and the medial branch extends until the turbinal becomes a single superior branching point of contact that descends along the centre of the nasal fossa and splits to form two equal curved branches in contact with the lateral and medial septal wall.
Interturbinal (IT)—This turbinal is the most posterior of the assemblage, located inferolateral to the FT, with a gradual anterior gradient on the sagittal plane. It is located 19.6 mm posterior to the piriform aperture, with an arcuate branching structure that projects from the lateral fossa wall between ethmoturbinal I and II. This turbinal arches, mirroring the curvature of the frontoturbinal (as seen in slices 156, 173, and 179 of the specimen CT scan; Figure 3E–G).
3.2.2. Turbinals of the Right Nasal Cavity
The right nasal fossa contains one nt, one ft, three et, one it, and one mt. The right nasal turbinals are well preserved overall, with all major turbinals mostly intact. There is a minor mediolateral distortion of et -I, -II, and -III.
Maxilloturbinal (mt)—The structure of this turbinal closely mirrors that of the paired MT in structure and position, located 0.7 mm posterior to the piriform aperture. The basal margins are once again mostly preserved along the turbinals full length, though the anterior margin is comminuted - as are the medially and laterally directed scrolls (as seen in slice 97 of the specimen CT scan; Figure 3A). The laterally directed scroll has begun to degrade into floating fragments.
Nasoturbinal (nt)—This turbinal closely matches the structure and position of the paired NT and is also well preserved, other than anterior deformation of the laterally directed scroll along the coronal plane (as seen in slices 137, 139, 146, and 156 of the specimen CT scan; Figure 3B–E). The turbinal is located 15.6 mm posterior to the piriform aperture.
Ethmoturbinal I (et-I)—The ET-I and et-I are similar, though the bulla of et-I does not detach to form a scroll which then reduces to a branching structure; instead, it detaches immediately from a bulla to branching morphology from the middle of the structure to the posterior (as seen in slices 139 and 146 of the specimen CT scan; Figure 3C,D and Figure 5A–F). The turbinal is positioned 12.2 mm from the piriform aperture. Furthermore, the bulla is much narrower in the cross-section and more elliptical than in ET-I.
Ethmoturbinal II (et-II)—This turbinal shares the same general arrangement as ET-II and is positioned similarly 14.1 mm from the piriform aperture. The full extent of the turbinals is not observable, either due to damage or poor CT resolution. The ET-II and et-II are almost identical anteriorly, as a branching structure in contact with the superior and lateral fossa wall (as seen in slices 146, 156, and 173 of the specimen CT scan; Figure 3B–F and Figure 5A–F). However, the two points of contact of the et-II bulla with the inferior lateral border of ET-I are more lateral and superior, respectively. The bulla is also much more convex posteriorly (as seen in slice 146 of the specimen CT scan; Figure 3D and Figure 5E–G).
Frontoturbinal (ft)—Similarly to FT, the ft has an arcuate morphology—a regular arc that increases in circumference anteroposteriorly (as seen in slices 137, 139, 146, 156, and 173 of the specimen CT scan; Figure 3D–F). The arch is more convex, however, which may be a natural disparity or possibly due to mediodorsal compression and deformation. This turbinal is slightly more posterior than its counterpart, 18.4 mm from the piriform aperture.
Ethmoturbinal III (et-III)—This turbinal is almost identical to ET-III, other than that a greater portion of the morphology is not evident in the scan, either due to damage or poor CT resolution (as seen in slice 179 of the specimen CT scan; Figure 3G). It is difficult to assess whether the turbinal naturally ceases at the scrolled portion of the structure or is fragmented and would extend to a complete bulla (as seen in slice 190 of the specimen CT scan; Figure 3H and Figure 5C,D,H). It is similarly located to its opposing paired turbinal, 12 mm posterior to the piriform aperture.
Interturbinal (it)—This turbinal is almost completely identical to its right pair, though the anterior segment which extends anteromedially is slightly shorter (as seen in slices 156, 173, and 179 of the specimen CT scan; Figure 3E–G and Figure 5A–H). The positioning is almost identical too, 18.6 mm posterior to the piriform aperture. It is difficult to distinguish whether this is a natural variation or due to some minor destruction of the turbinal.
Numerous floating fragments are evident in both anterior nasal fossae. Due to severe taphonomic alterations, there is little evidence that can be derived from these fragments, and it cannot be determined which of the turbinals they originate from.
The total surface area of the Mioeuoticus specimen’s reconstructed olfactory assemblage is 1109.92 mm^2^, though it is likely that this is an underestimation as breakage and fragmentation are evident in every turbinal structure. A further estimate of total surface area which may be more representative is deduced by considering only the most complete turbinal from each pair and multiplying these data by two, the result of which is 1184 mm^2^. The surface areas of the individual olfactory components are summarised in Table 2. The maxilloturbinals, determined from the average of each paired olfactory component, comprise the most substantial portion (26%) of the total surface area. This is followed by ethmoturbinals I (21%) and nasoturbinals (15%), ethmoturbinals II (18%), ethmoturbinals III (11%), frontoturbinals (6%), the remaining floating fragments (4%), and, finally, the interturbinals (2%).
3.2.3. Vomeronasal Groove
An additional significant observation made when analysing the inferior base of the rostral cranium of the Mioeuoticus specimen is the vomeronasal groove (Figure 6). The vomeronasal groove is a trough-like structure (e.g., Figure 6A) that houses the cartilaginous tube of the vomeronasal organ, which sits at the base of the nasal cavity anteroventrally to the septum (e.g., Figure 6B) and is covered in odorant receptors [63,64,65,66].
Modern strepsirrhines possess a vomeronasal organ [34,35,41,67,68,69], though due to the endangered nature of many strepsirrhine taxa, anatomical analysis is restricted to only a small number of taxa [70,71,72,73,74,75]. Even rarer are the studies to reconstruct the vomeronasal organ of extinct strepsirrhines using modern comparatives [66], and anatomical analysis of fossil primate vomeronasal grooves [76]. This study constitutes the oldest evidence of the vomeronasal groove in a fossil crown strepsirrhine and certainly is the first record of this characteristic in fossil lorisoids.
3.3. Audition
The general morphology of the cochlea is evident, but the coiling structure cannot be discerned in the cast. The length of the cochlear canals from the apex of the spiral to the terminal base (near the middle ear and the oval window) can be measured at 3.94 mm (left) and 4.02 mm (right). The three oval semicircular canals (SCCs) are apparent in both bony labyrinths: the anterior semicircular canal (ASC), the posterior semicircular canal (PSC), and the lateral semicircular canal (LSC) (Figure 7).
None of the canal reconstructions are complete, though it is unclear whether this is due to breakage or scan resolution (Figure 7 and Figure 8). The angles between all canals are far from orthogonal, with a large average deviation from orthogonality of 12.7° (Table 3).
The ASCs and PSCs have a degree of torsion, whereas the LSCs are fairly planar. The ASCs have the greatest degree of torsion, with the dorsal edge bent significantly forward. Of the SCCs, the ASC has the greatest height and width, and the PSC has the shortest (Table 4). The posterior limb of the LSC with the ampullar limb of the PSC fuse to form a secondary common crus orthogonal to the plane of the LSC length.
4. Discussion
4.1. Olfaction
A direct comparison between several modern and extinct lorisid olfactory systems, as well as that of the fossil prosimian Rooneyia, analysed in Lundeen & Kirk [38], is undertaken here to highlight the most prominent and remarkable characteristics of the reconstructed turbinals of Mioeuoticus (Table 5). Rooneyia is included as this taxon is the only proxy we have for what might be primitive more broadly in primates, but it is not a direct ancestor since it is generally recovered as a fossil haplorrhine.
The turbinals are described and compared from the relative posterior-to-anterior position in the olfactory system.
As aforementioned, Mioeuoticus has notably long, elongated maxilloturbinals. Compared to Loris, the MT/mt in Mioeuoticus is proportionally much longer than the other turbinals in the assemblage (though there seems to be anterior breakage of ET/et-I, and so it could appear reduced) (Lundeen [41]: Figure 9.2C,D). The maxilloturbinals of Mioeuoticus stand prominently anterior to the rest of the turbinals in the olfactory system. This contrasts the MT/mt of modern counterparts Loris, Nycticebus, Arctocebus, and Perodicticus, which sit almost directly under the other turbinals in the olfactory system (Lundeen [41]: Figure 9.2A–H). Thus, the modern counterparts have a more compact, overlaid arrangement than Mioeuoticus.
The MT/mt of Mioeuoticus and Perodicticus are broadly similar in relative surface area and shape; however, the turbinals in the Mioeuoticus specimen are too fractured to make a more detailed comparison. The maxilloturbinal total surface area (MTSA) of Mioeuoticus is a relatively small 151.36 mm^2^ in comparison to extant strepsirrhines and haplorhines studied in Lundeen & Kirk [38], e.g., the MTSA of Rooneyia is only 145.42 mm^2^, Loris is 221.20 mm^2^, Nycticebus is 221.74 mm^2^, and Perodicticus is considerably higher at 366.35 mm^2^ (the total maxilloturbinal surface area of all comparative primate specimens from Lundeen & Kirk [38] is available in Supplementary Table S4).
From the rostral-to-caudal limits of the turbinal assemblage of Mioeuoticus, the NT/nt has a similar relative surface area, position, scrolled structure, and slope gradient to that seen in Arctocebus and Nycticebus (Lundeen [41]: Figure 9.2A,B,G,H). The NT/nt is smaller than that of Loris and Perodicticus, with a shallower slope gradient on the sagittal plane. The NT/nt of these modern counterparts do, however, have a similar posterior position relative to the MT/mt in Mioeuoticus (Lundeen [41]: Figure 9.2C–F). The NT/nt of Nycticebus and Perodicticus contrast all of these specimens with a more anterior relative position to the MT/mt. There is a shallower slope gradient of the NT/nt on the sagittal plane in the olfactory system of Mioeuoticus than in that of Nycticebus, and significantly so than that of Loris, and the turbinal is not scrolled towards the posterior in these modern counterparts.
The FT/ft of Mioeuoticus are similar in relative surface area to Nycticebus, though with an arc structure rather than the scrolls seen in the modern counterpart (Lundeen [41]: Figure 9.2B). This arcuate structure is strikingly similar to that seen in Rooneyia (Lundeen & Kirk [38]: Figure 1F,G). The posterior of the FT/ft has an increasingly steep slope gradient towards the anterior in Mioeuoticus and Rooneyia, which mirrors that of the NT/nt. This characteristic is also evident, though much steeper, in Nycticebus. The FT/ft sits laterally to the NT/nt in Mioeuoticus, as opposed to a superomedial position in Loris.
The ET/et-I in the modern counterparts extend as far laterally as the MT/mt, whereas the ET/et-I of Mioeuoticus are in a significantly more posterior relative position, more posterior than the NT/nt too. ET/et-I are much larger in relative surface area in the modern specimens than in Mioeuoticus and Rooneyia. ET/et-I in Mioeuoticus are much smaller in relative surface area and length than in Loris, though these turbinals remain similar in relative size in each species. These ethmoturbinals also have weaker scrolled structures than those of Loris and Arctocebus, though it is difficult to determine if this is because of breakage in the latter. In Mioeuoticus and Rooneyia, the anterior margin of ET/et-I forms a mediolaterally compressed and laterally directed enclosed bulla that increases in volume anteriorly until it opens to a scroll structure (Lundeen & Kirk [38]: Figure 1E,F). ET/et-II and -III are strikingly similar in relative surface area, position, structure, and slope gradient on the sagittal plane in Arctocebus, Nycticebus, and Mioeuoticus, though much greater in surface area in Loris and Perodicticus.
The most striking feature in the olfactory system of Mioeuoticus is the location of the interturbinal. In lemuroids, ET/et-II originates from ET/et-I and/or the horizontal laminae and so the IT/it are located between the shared basal laminae of ET/et-I and -II. In contrast, in lorisoids, ET/et-II make contact with the lateral nasal wall directly and so the IT/it are located between ET/et-II and ET/et-III [38]. The IT/it of Mioeuoticus are unique in that, anteriorly, they are located between the shared basal laminae of ET/et-I and -II and ET/et-III, as seen in lemuroids and also in Rooneyia (Lundeen & Kirk [38]: Figure 1G). However, moving posteriorly on the coronal plane, ET/et-II contact the lateral nasal wall directly and so the interturbinal location changes. Notably, this is not between ET/et-II and ET/et-III but instead between ET/et-I and ET/et-II, so not the exact point of contact seen in modern lorisoids. This turbinal arrangement is not seen in any other known strepsirrhine, and the shared turbinal morphology with lemuroids likely denotes the primitive status of Mioeuoticus within Lorisidae, consistent with the idea that it might be a stem member of the family [14]. The IT/it of Nycticebus are similar in relative surface area to the rest of the assemblage, though it has a more elongated structure in Nycticebus than in Mioeuoticus. The IT/it of the olfactory system in Loris is similar in relative position to the IT/it of Nycticebus. However, it is more comparable in relative length to the IT/it in Mioeuoticus and much larger in relative surface area.
The most disparate characteristic in Mioeuoticus would be the absence of ET/et-IV, a feature which is present in Nycticebus, Perodicticus, Arctocebus, Loris, and Rooneyia (Lundeen [41]: Figure 9.2C,D; Lundeen & Kirk [38]: Figure 1H); though, as aforementioned, this ethmoturbinal is suspected to be present in Mioeuoticus and yet barely distinguishable due to the scan resolution. This is more likely when considering that the nasal fossae of strepsirrhines typically house 4–5 ethmoturbinals [38].
In several respects, there are close similarities in the organisation of the nasal turbinals between Mioeuoticus and Rooneyia [38]. The posterior of the NT/nt in Mioeuoticus and Rooneyia are connected to the superior wall of the nasal fossa via only the superior-most basal lamina (Lundeen & Kirk [38]: Figure 1F,G). The central portion of both NT/nt takes the form of a mediolaterally compressed scroll and the posterior-most portion of the NT/nt is more restricted in cross-sectional area and positioned in the superior nasal cavity, above the ET/et-I and superomedial to the FT/ft (Lundeen & Kirk [38]: Figure 1E). The NT/nt of Rooneyia contrast in that an anterior portion of the turbinals extends laterally to a more ventral position and then diverges posteriorly just anterolateral to the MT/mt (Lundeen & Kirk [38]: Figure 1C). Posteriorly, the ET/et of both Mioeuoticus and Rooneyia are branching structures that make contact with the superior and lateral fossa wall (Lundeen & Kirk [38]: Figure 1F–H), then the connection with the superior fossa wall is lost as the structure modifies into a singular bulla in the central and anterior portion of this turbinal (Lundeen & Kirk [38]: Figure 1B–E). The ET/et-II turbinals of Mioeuoticus and Rooneyia are inferomedial to ET/et-I, orientated anteromedially with a steep anterior gradient on the sagittal plane. It is difficult to discern whether the ET/et-III of Mioeuoticus are anteriorly bullar, like those of Rooneyia (Lundeen & Kirk [38]: Figure 1G), or scrolled, as this is not clear on the CT rendering. Posteriorly, the ET/et-III in both species curve more steeply to form a single medial scroll with branches that make contact with the lateral and anterior septal walls. The only considerable divergence is that Mioeuoticus has a much larger overall total surface area (OTSA) relative to body size (1109.92 mm^2^) than Rooneyia (408.31 mm^2^).
When comparing the total olfactory turbinal surface area relative to the cranial size of Mioeuoticus with that of extant and extinct primates in Lundeen & Kirk [38] study, it is evident that Mioeuoticus has an olfactory turbinal surface area that is broadly similar to the similar-sized strepsirrhines Propithecus verreauxi, Euoticus elegantulus, and Perodicticus potto (Figure 9). The OTSA of all comparative specimens from Lundeen and Kirk [38] and Mioeuoticus specimen KNM-RU 2052 are available in Supplementary Table S4. The surface area of Mioeuoticus is slightly smaller and plots just below the lower limit of strepsirrhine distribution. However, this is certainly influenced by underestimation caused by fragmentation and breakage.
Mioeuoticus has a notably extended olfactory recess length (7.53 mm). The presence of an olfactory recess is primitive for primates [77,78]. A large olfactory recess has been considered characteristic of mammals with a well-developed sense of smell [79] and haplorrhines are reported to lack an olfactory recess [37,60]. A larger olfactory recess results in greater odorant residence time, promoting slower airflow and improving the residence time of odorants within the olfactory region [77]. Further assessment of strepsirrhine airway morphometry and functional implications can be made by analysing the position of ethmoturbinals relative to the olfactory recess, e.g., Smith et al. [37]. However, as aforementioned, the resolution of the cranial scan in this study is too poor to fully determine the presence and extent of the fourth pair of ethmoturbinal in the Mioeuoticus specimen.
Biplot showing log-transformed olfactory turbinal surface area (measured in millimetres²) and log-transformed skull geometric mean (measured in mm). Skull geometric mean of cranial length and width is measured following Muchlinski [80]. The data used in this graph are modified from the supplementary information of Lundeen and Kirk [38]. These modified raw data are also available in Supplementary Tables S3 and S4. Taxon abbreviations: Al = Alouatta; Ao = Aotus; At = Ateles; Av = Avahi; Ct = Carlito; Cv = Cynocephalus; Cx = Callithrix; Da = Daubentonia; Ee = Euoticus; Eu = Eulemur; Ga = Galeopterus; Gm = Galago moholi; Gs = Galago senegalensis; Ha = Hapalemur; Hy = Hylobates; In = Indri; Lm = Lepilemur; Lo = Loris; Ma = Mandrillus; Mi = Miopithecus; Mo = Mioeuoticus; Mz = Mirza; Ny = Nycticebus; Ot = Otolemur; Pe = Perodicticus; Ph = Presbytis; Pi = Piliocolobus; Pl= Plecturocebus; Pv = Propithecus; Pt = Ptilocercus; Rv = Roonyeia; Sa = Saimiri; Tb = Tupaia belangeri; Tg = Tupaia glis; and Va = Varecia.
4.1.1. Audition and Proprioception
Despite some features being undeterminable in the Mioeuoticus inner ear reconstruction, there are still key anatomical characteristics that give insights into the auditory capabilities and locomotor agility of the specimen, and also the phylogenetic status of the species.
The fusion of the anterior and posterior semicircular canals seen in the Mioeuoticus holotype is a common characteristic in numerous extant and extinct primates [81], and the orthogonality between this structure and the plane of the lateral semicircular canal is a condition that can also be observed in several primates, including Lorisidae [82]. The oval lateral canals and short common crura of the specimen are commonly exhibited by lorisoids, as opposed to the rounded lateral canals and long common cura which are indicative of Lemuroidea [83]. The Mioeuoticus cochlea is broad and bulky, similar in morphology to that seen in modern Loris (Figure 10i(A–H)) though significantly larger than Nycticebus and Perodicticus (Figure 10ii(A–H)). The common crus is short relative to the cochlea and semicircular canals, with a narrow utricle and vestibule, similar to that seen in Nycticebus (Figure 10iii(A–H)). The strong ovate shape of the semicircular canals is comparable between Perodicticus and Mioeuoticus, though the relative size between the canals is morphologically closer to that of Nycticebus (Figure 10iv(A–H)).
The 105° angle between the anterior and posterior semicircular canals in the Mioeuoticus specimen is remarkably consistent with the angles between the arcs at the greatest width of the anterior and posterior semicircular canals on the transverse plane (ASCm < PSCm) of platyrrhine and catarrhine anthropoids reported by LeBrun et al. [61]. According to this study, most primates exhibit an ASCm < PSCm much greater than 90°, and the platyrrhine and catarrhine anthropoids have an overlapping distribution spanning 101–110°. Following the current models correlating canal orthogonality and locomotor agility (e.g., [84,85]), we hypothesise that the significant deviation from orthogonality of the angles between all of the canals in the Mioeuoticus inner ear reconstruction may indicate slower rotational head speeds and a lower mean vestibular sensitivity to motion. These results are consistent with what would be expected for this taxon, but we take the opportunity to highlight this method has proven sometimes unreliable [86,87].
Walker et al. [7] previously measured the curvature radii in this specimen. The ASC radius is 2.6 mm, the PSC radius is 3.3 mm, and the LSC radius is 2.9 mm. The prediction for locomotor agility for fossil taxa based on a predictive equation for primates calculated from the values in Table 1 of Silcox et al. [88] predicts that animals with these value ranges are between 2 and 3 on a six-point agility scale (1 = extremely slow, 2 = slow [e.g., Loris], 3 = medium slow, 4 = medium [e.g., Cebus, Cheirogaleus], 5 = medium fast, 6 = fast [e.g., Galago, Saimiri]) formed by Spoor et al. [16] (e.g., [49,85,89]). Furthermore, the lateral canal has the shortest radius of the bony labyrinth in Mioeuoticus, and this characteristic is considered to be diagnostic of less agile behaviours [90,91].
The data in this auditory analysis of the M. shipmani holotype must be considered with caution, as several publications have determined that it is difficult to predict and differentiate vestibular sensitivities of slower-moving species from faster-moving ones when only a single specimen is available, as the morphologies of various individuals within a species and across several species overlap considerably (e.g., [66,90,91]). Recent studies indicate that the results acquired from measuring the orthogonality of the semicircular canals in any organism can lead to a fair amount of variability in results, particularly in a slow-moving taxon (e.g., Gonzales et al. [59]).
4.1.2. Vision
When making direct comparisons with the extant primate daOFQs and nsOFQs from Figure 11 in our study with the data in Kirk and Kay [21], the Mioeuoticus specimen has OFQs that fall well below the range of OFQs for extant diurnal haplorhines, borders the lowest range of OFQs for extant diurnal strepsirrhines (the lowest daOFQ being −56.9 for Hapalemur griseus), sits within range of extant nocturnal cathemeral strepsirrhines, and sits above nocturnal haplorhines (Aotus). The most similar OFQ values from the extant primates analysed in Kirk and Kay [21] are the diurnal strepsirrhine Hapalemur griseus (daOFQ = −56.9, nsOFQ = 16.40) and the nocturnal strepsirrhines Galago matschiei (daOFQ = −57.1, nsOFQ = 15.2) and Perodicticus potto (daOFQ = −57.2, nsOFQ = 15.5).
Mioeuoticus is known to be a strepsirrhine lorisid [1,5,14], and the OFQs calculated here have low values, which indicates a high central retinal summation, comparable to extant strepsirrhines with a nocturnal activity pattern. Thus, it is probable that Mioeuoticus had central retinas with a large proportion of high summation rods which increased sensitivity to light at the expense of visual acuity—an adaptation to low light levels.
The similarity in OFQs seen between this specimen and Perodicticus potto could suggest that these extant nocturnal strepsirrhines have retained a primitive optic morphology and, thus, nocturnal vision, as a generally plesiomorphic character state inherited from the ancestral Mioeuoticus. Accordingly, it may be surmised that this ancestral state character may closely resemble the visual acuity and optic morphology of these extant descendants.
5. Conclusions
Our research provides a unique new understanding of the reconstructed behaviour of Mioeuoticus and how the sensory sensitivities in this fossil lorisid compare to extant members of this family. Computed tomographic (CT) analyses of the sensory systems visible in the Mioeuoticus holotype specimen suggest it had a keen olfactory sense, less agile locomotion, and visual adaptations for either a nocturnal lifestyle, like that of modern lorisids, or a cathemeral lifestyle. Because the entire radiation of modern lorisids, as well as its sister group (Galagidae), are nocturnal, the most parsimonious explanation for the activity pattern of M. shipmani is that this animal belonged to an entirely nocturnal radiation of lorisoids. The unique turbinal arrangement is not seen in any other known strepsirrhine and shares a turbinal morphology with lemuroids, which likely denotes the primitive status of Mioeuoticus within Lorisidae. Our data are consistent with Mioeuoticus being a member of the family Lorisidae, as it shares morphological features in the olfactory system, orbital structure, and inner ear anatomy with modern representatives. Our results show that the behavioural pattern seen in the modern perodicticine lorisid Perodicticus potto most closely resembles that reconstructed in Mioeuoticus shipmani, and therefore Perodicticus could be used as an appropriate analogue for the early stages of sensory evolution in lorisids.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1López-Torres S. Silcox M.T. What We Know (and Don’t Know) About the Fossil Records of Lorisids Evolution, Ecology and Conservation of Lorises and Pottos Cambridge University Press Cambridge, UK 20203346
- 2Burrows A.M. Nash L.T. Hartstone-Rose A. Selig K.R. Silcox M.T. López-Torres S. What Role Did Gum-Feeding Play in the Evolution of the Lorises?Cambridge University Press Cambridge, UK 2020154 p
- 3Nekaris A. Bearder S.K. The Lorisiform primates of Asia and mainland Africa Primates Perspect.20072362445
- 4Nekaris K.A.I. Starr C.R. Collins R.L. Wilson A. Comparative ecology of exudate feeding by lorises (Nycticebus, Loris) and pottos (Perodicticus, Arctocebus)The Evolution of Exudativory in Primates Springer New York, NY, USA 2010155168
- 5Seiffert E.R. Simons E.L. Attia Y. Fossil evidence for an ancient divergence of lorises and galagos Nature 200342242142410.1038/nature 0148912660781 · doi ↗ · pubmed ↗
- 6Pickford M. Wanas H. Soliman H. Indications for a humid climate in the Western Desert of Egypt 11–10 Myr ago: Evidence from Galagidae (Primates, Mammalia)C. R. Comptes Rendus Palevol 2006593594310.1016/j.crpv.2006.09.001 · doi ↗
- 7Walker A. Ryan T.M. Silcox M.T. Simons E.L. Spoor F. The semicircular canal system and locomotion: The case of extinct lemuroids and lorisoids Evol. Anthropol. Issues News Rev. Issues News Rev.20081713514510.1002/evan.20165 · doi ↗
- 8Fulwood E.L. Shan S. Winchester J.M. Gao T. Kirveslahti H. Daubechies I. Boyer D.M. Reconstructing dietary ecology of extinct strepsirrhines (Primates, Mammalia) with new approaches for characterizing and analyzing tooth shape Paleobiology 20214761263110.1017/pab.2021.9 · doi ↗