Typification of the Economically Important Species Thyreophagus entomophagus (Acari: Astigmata: Acaridae) Used for the Industrial Production of Predatory Mites: The Designation of a Neotype with Detailed Morphological and DNA Sequence Data
Pavel B. Klimov, Vasiliy B. Kolesnikov, Alexander A. Khaustov, Vladimir A. Khaustov, Jonas Merckx, Marcus V. A. Duarte, Dominiek Vangansbeke, Ilse Geudens, Almir Pepato

TL;DR
This paper clarifies the identity of a mite species used in agriculture, designating a new standard specimen and revealing a new species, which helps improve biological pest control methods.
Contribution
The designation of a neotype for Thyreophagus entomophagus and the discovery of a new species, Thyreophagus holda, resolve taxonomic confusion and highlight traits beneficial for mass production in biocontrol.
Findings
Thyreophagus entomophagus lacks a specialized deutonymph stage, simplifying mass production.
A previously misidentified population is a new species, Thyreophagus holda.
Asexual reproduction and absence of deutonymphs are derived traits beneficial for biocontrol.
Abstract
The mite Thyreophagus entomophagus is widely used in agriculture as a food source for breeding predatory mites, which are important for biological pest control. However, the identity of this species has been uncertain due to its incorrect identifications involving both morphology and DNA sequence data. To resolve this, we carefully examined a commercial population, selecting a new type specimen from this population to standardize its name. We also discovered that a population previously thought to belong to this species is actually a new species, Thyreophagus holda. These findings clarify that Th. entomophagus lacks a specialized life stage (deutonymph) in its life cycle, making it easier and more efficient to mass-produce. Our phylogenetic analysis shows that this trait, along with asexual reproduction, evolved after the origin of the genus Thyreophagus. We suggest that these…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
 Figure 1
Figure 1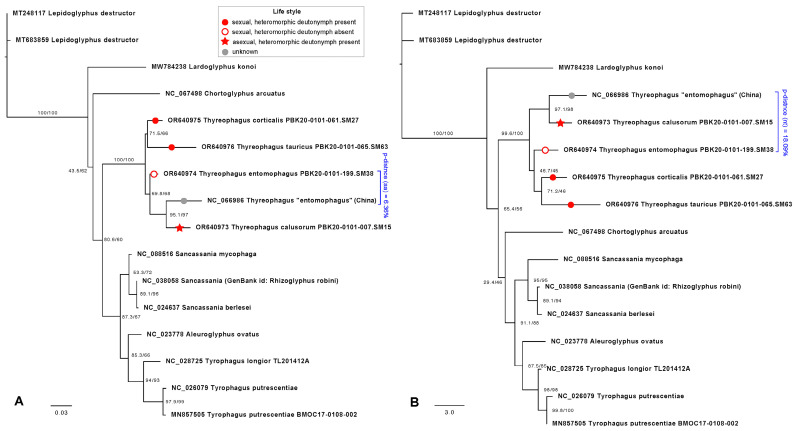 Figure 2
Figure 2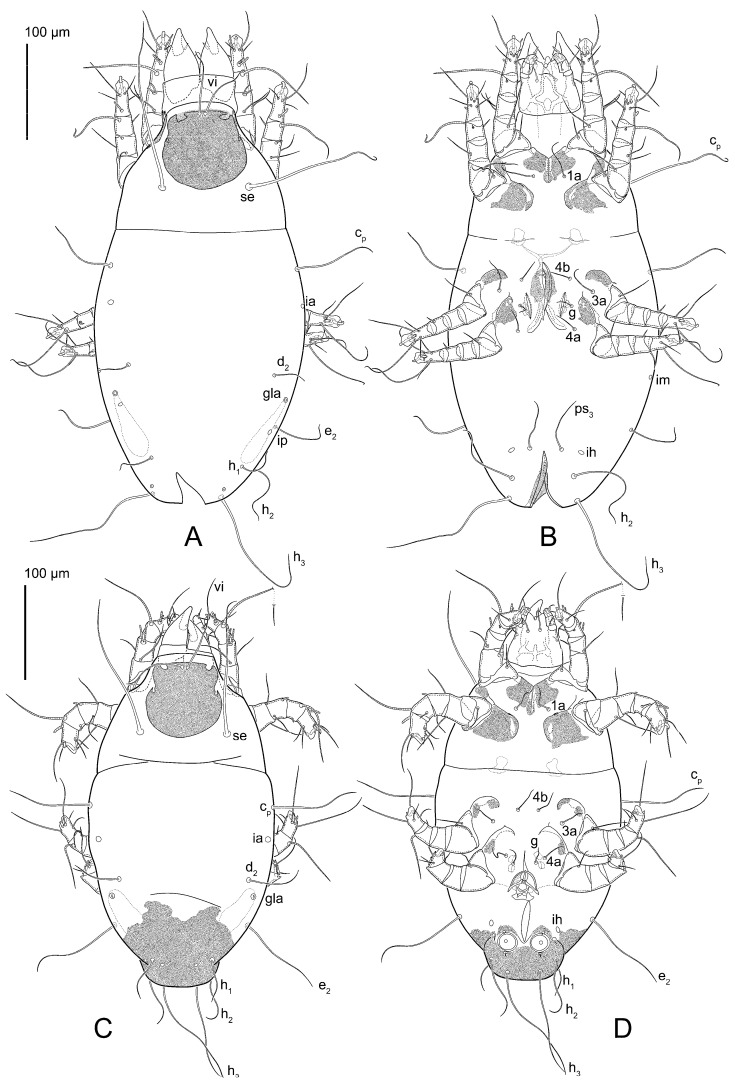 Figure 3
Figure 3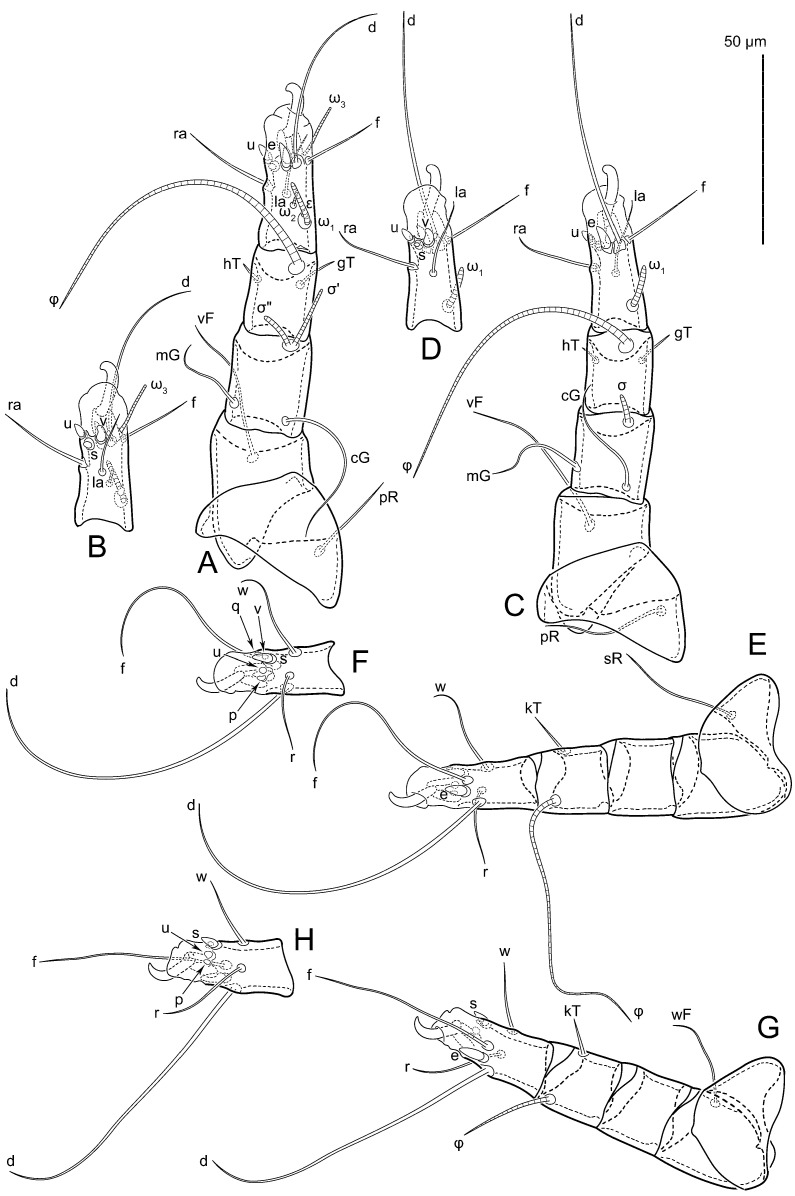 Figure 4
Figure 4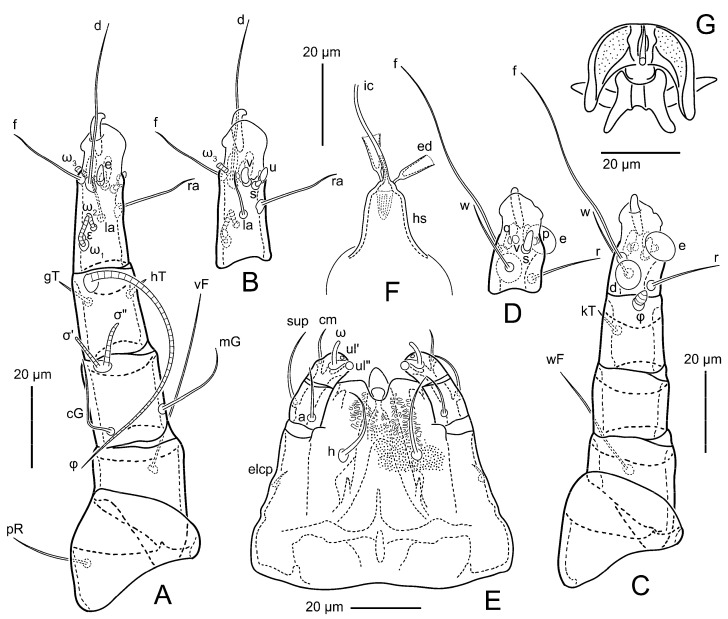 Figure 5
Figure 5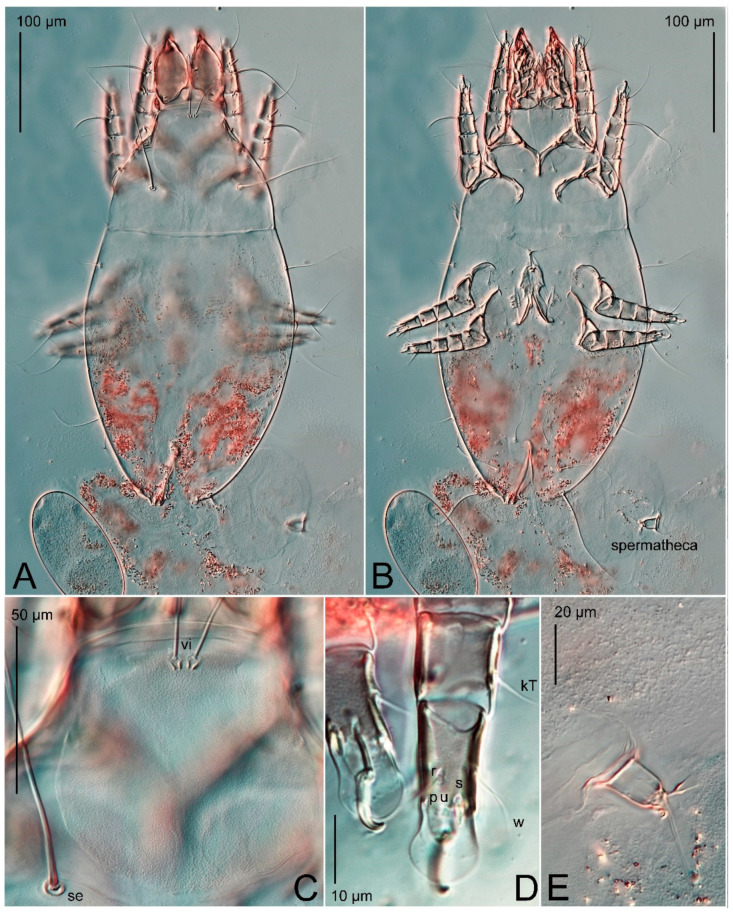 Figure 6
Figure 6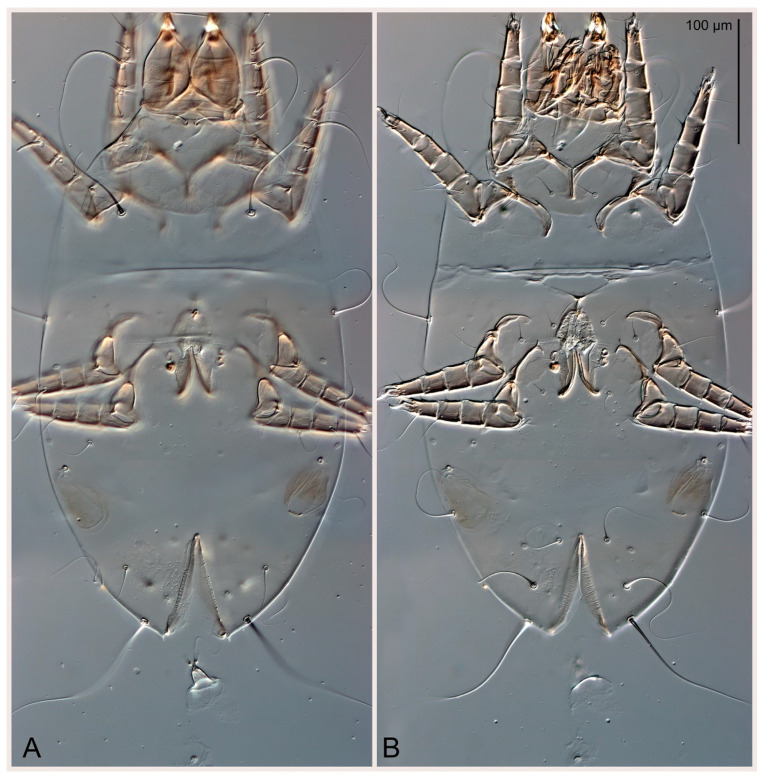 Figure 7
Figure 7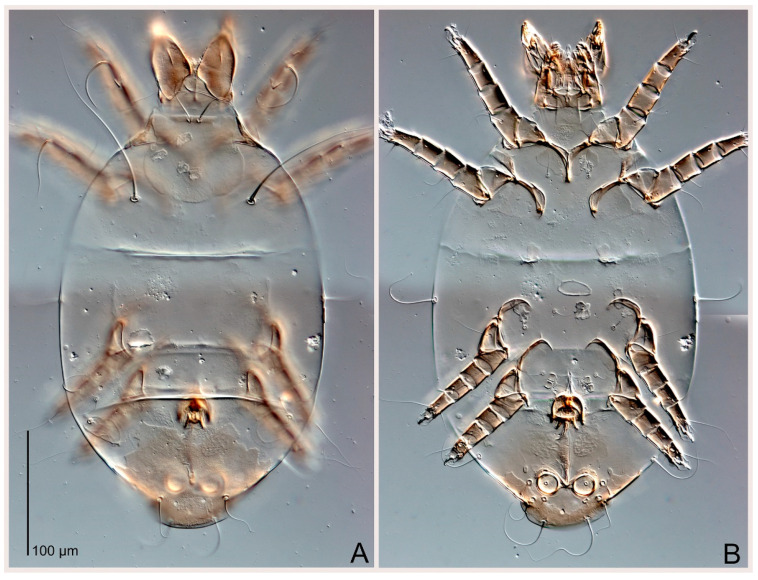 Figure 8
Figure 8 Figure 9
Figure 9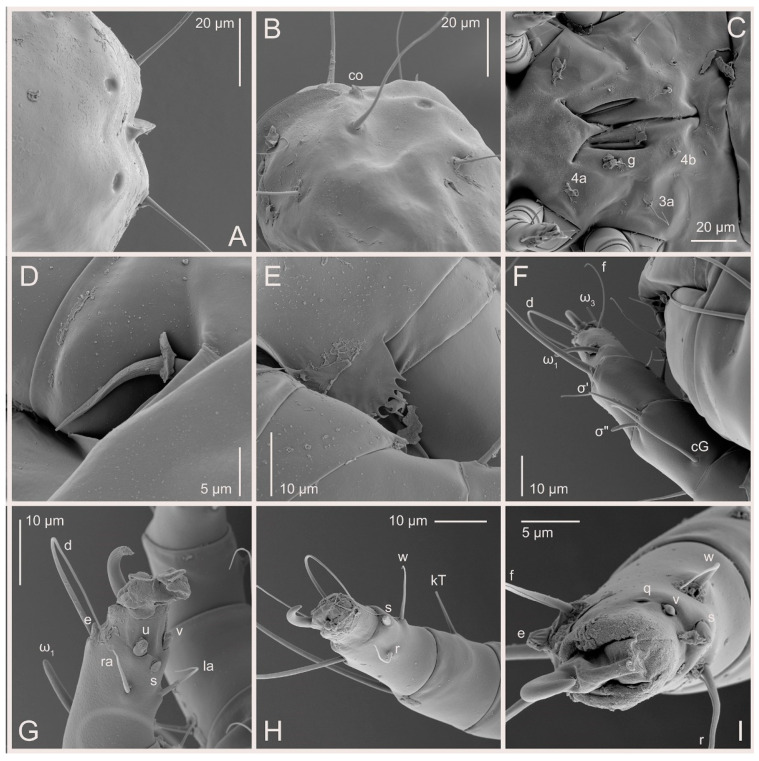 Figure 10
Figure 10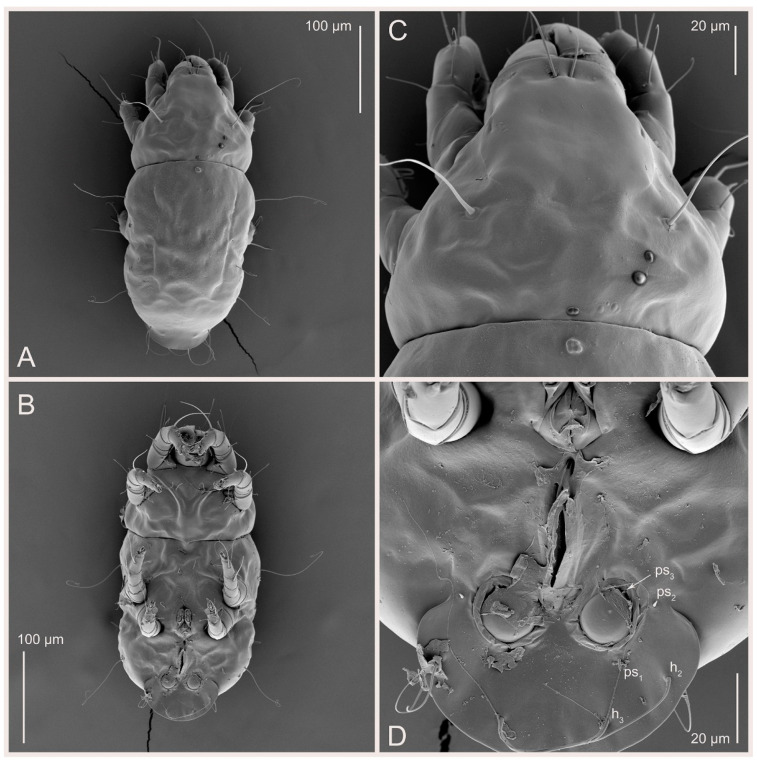 Figure 11
Figure 11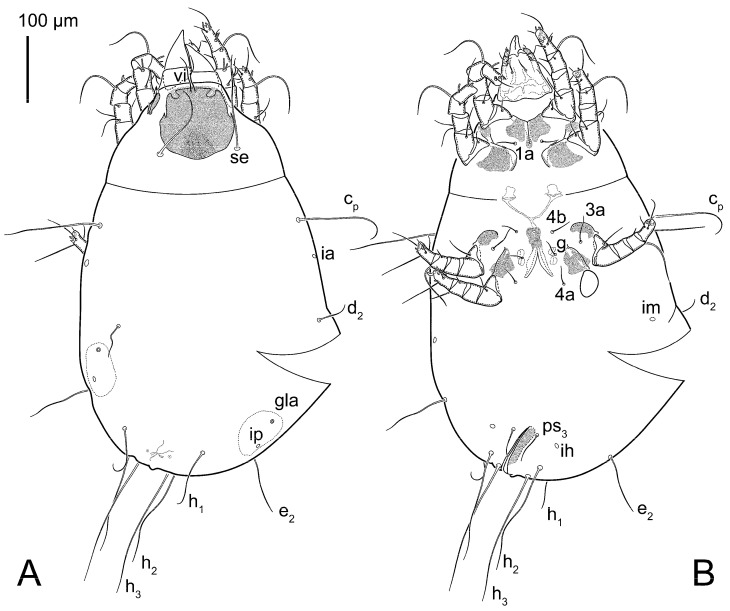 Figure 12
Figure 12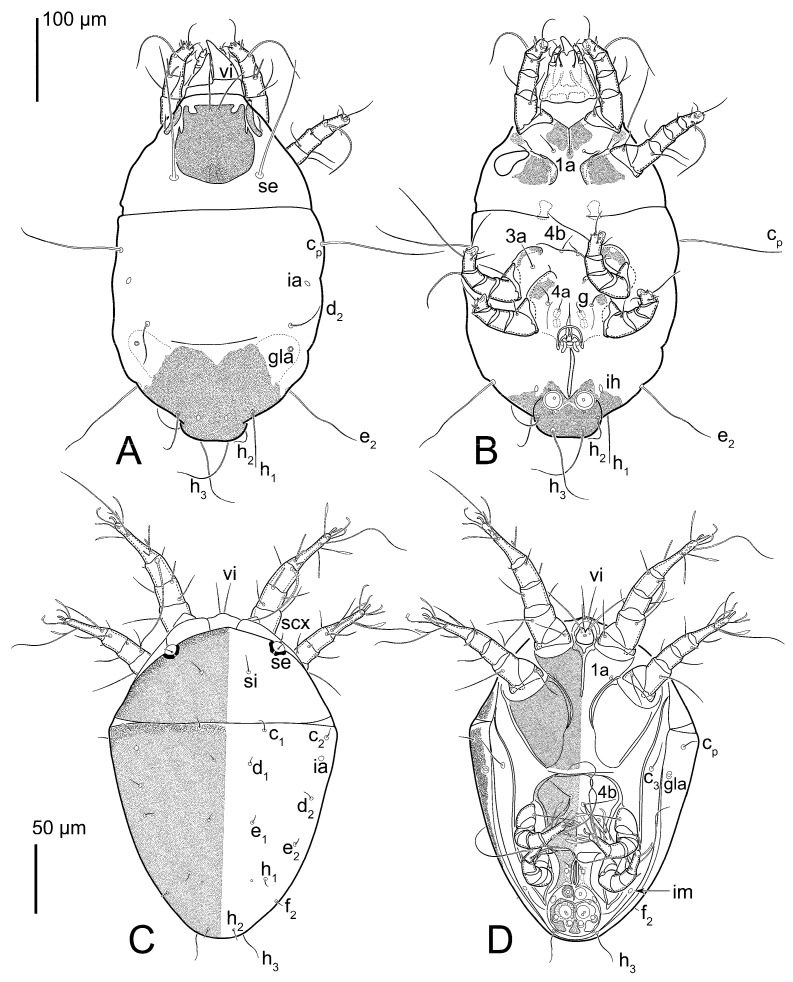 Figure 13
Figure 13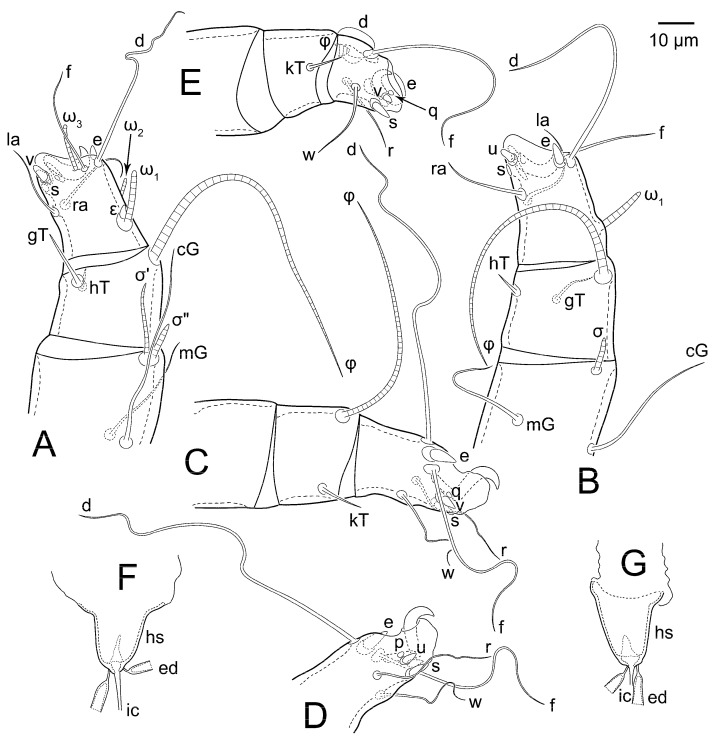 Figure 14
Figure 14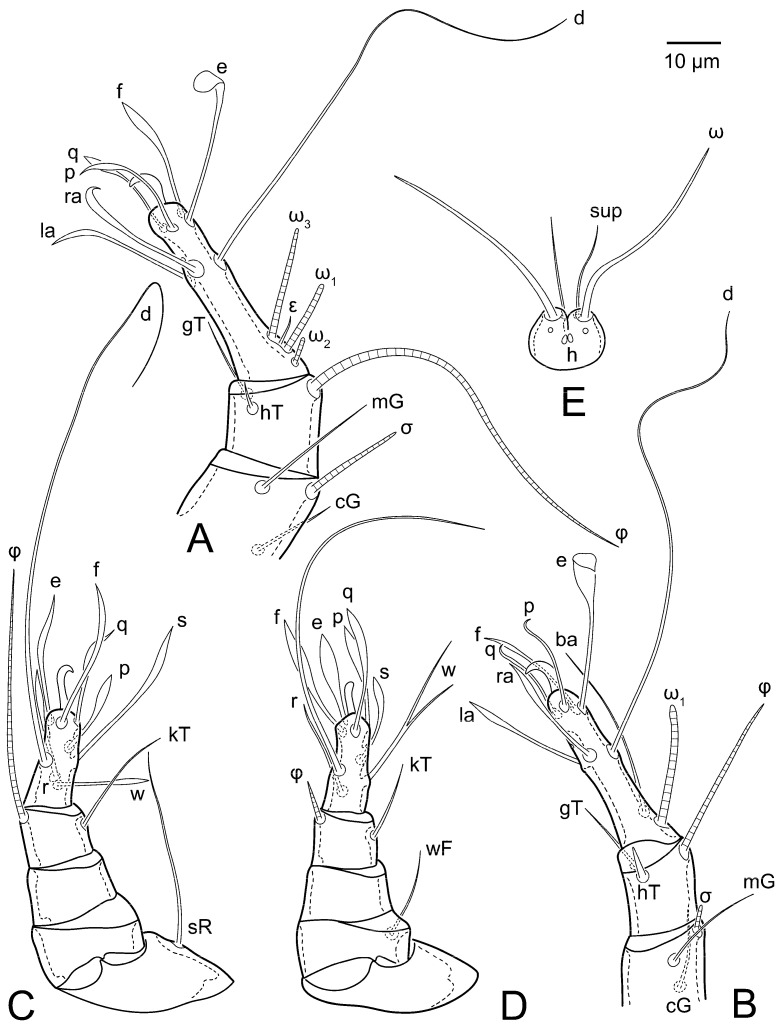 Figure 15
Figure 15 Figure 16
Figure 16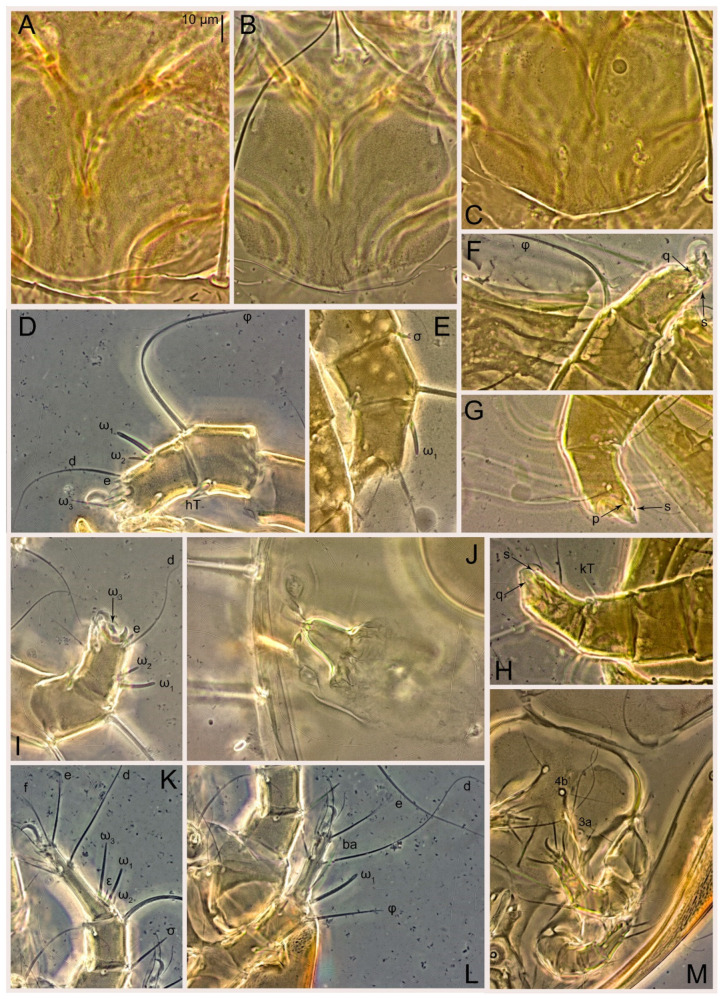 Figure 17
Figure 17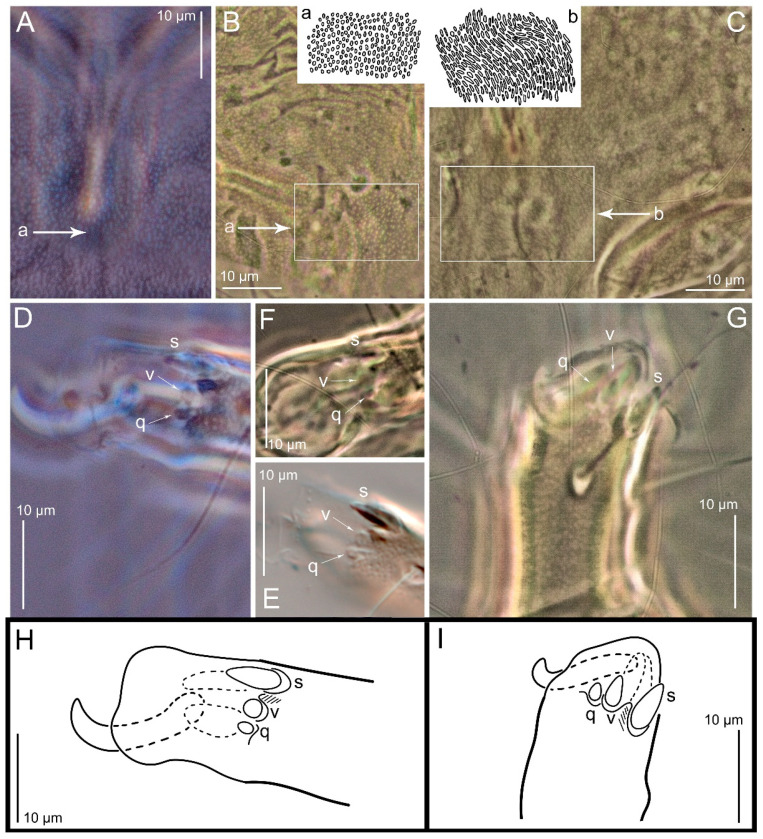 Figure 18
Figure 18- —Ministry of Science and Higher Education of the Russian Federation
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsStudy of Mite Species · Insect-Plant Interactions and Control · Forest Insect Ecology and Management
1. Introduction
Plant pests, particularly insect pests, pose a significant threat to global agricultural production. While chemical pesticides are effective in controlling these pests, they have adverse environmental and health effects, including the development of pesticide-resistant insect populations. Biological control, which utilizes beneficial insects and mites to prey on pest species, offers a more sustainable alternative. Predatory mites, such as those in the family Phytoseiidae, have shown promise in controlling pests like thrips, whiteflies, and red spider mites [1,2,3,4]. The mass rearing of these beneficial insects and mites is important for their widespread application in agricultural settings. While early rearing systems relied on natural food sources, recent technological advances have focused on using factitious prey mites, such as stored product mites, to provide a consistent and controlled food supply for predators [1,2,3,4]. Successful mass rearing of beneficial insects and mites is essential for the continued development and implementation of biological control strategies, reducing reliance on chemical pesticides, and promoting more sustainable agricultural practices.
Thyreophagus entomophagus (Laboulbène and Robin, 1862) (Acaridae) has gained popularity as a factitious prey mite due to its favorable characteristics, such as ease of consumption by predators and being less allergenic and less harmful as a stored food pest compared to other species like Tyrophagus putrescentiae (Schrank, 1781) [1,2,3,4]. Currently, Th. entomophagus is widely used in the mass production of predatory mites globally [5,6,7].
Historically, the specific name Thyreophagus entomophagus was proposed as Acarus entomophagus by Laboulbène in 1852 [8]; however, this name was a nomen nudum and not compliant with ICZN rules. An ICZN-compliant description was made in 1862 under the name Tyroglyphus entomophagus by Laboulbène and Robin for mites damaging entomological collections in Southern France [9]. In 1874, Rondani established a new genus, Thyreophagus, to include this species, leading to the currently used binomen Thyreophagus entomophagus. Since the type material of Th. entomophagus has been lost [10], it was redescribed from adults collected in the Birmingham region of the United Kingdom, without proposing a neotype by A. Fain in 1982 [10]. This species concept continues to be widely used today, however, deutonymphs remained unknown. Phoretic deutonymphs of Th. entomophagus were later described from a culture started from specimens collected in a sparrow nest in Dahlem, Berlin, Germany [11], although adults from this German population have never been described. Remarkably, industrially reared populations of Th. entomophagus never produce heteromorphic deutonymphs. This suggests that either cryptic species are involved (one can produce deutonymphs while the other cannot), or some populations of Th. entomophagus may have permanently lost the ability to produce heteromorphic deutonymphs. Furthermore (personal observations), GenBank has two mitochondrial COX1 sequences identified as Th. entomophagus from China (NC_066986) and Russia (OR640974) that differ by a 21.16% COX1 K2P distance [7], indicating that these samples represent two different species. These findings suggest that the taxonomic status of the economically important species Th. entomophagus is uncertain in terms of both morphology and genetics. There is an urgent need to standardize the usage of the name to ensure taxonomic stability and scientific repeatability of research that uses the name Thyreophagus entomophagus.
In this study, we carefully examine the German deutonymph-producing population of Thyreophagus entomophagus to determine if there are morphological differences compared to a commercial population that never forms heteromorphic deutonymphs and compare it with specimens from Birmingham region (United Kingdom) described in Fain [10]. Additionally, we provide a detailed morphological description of the commercial population, supported by COX1 sequences and notes on the phylogeny of Thyreophagus. Since the German populations were morphologically distinct from specimens in the commercial culture and the UK population, we propose standardizing the use of the name Thyreophagus entomophagus by designating a neotype based on the commercial population, where both the COX1 sequence and morphology are known.
2. Materials and Methods
Commercial cultures were obtained from a European biocontrol company on 20 November 2021 and maintained in the lab. The cultures were reared on a mixture of yeast and bran in specialized rearing units. The purity of the cultures was verified through morphological identification of a large number of mites (n = 50). Mites were collected using a camel brush, preserved in 96% ethanol, cleared in 80% lactic acid for 1–2 days, and mounted in Hoyer’s medium, followed by a 7-day drying period at 60 °C [7]. Voucher specimens were deposited at the Tyumen State University Acarological Collection and the Zoological Institute, Russian Academy of Sciences, Saint Petersburg, Russia; the neotype was deposited the Royal Belgian Institute of Natural Sciences, Brussels, Belgium. We also examined specimens collected from a sparrow nest in Dahlem, a suburban district of Berlin, by W. Knülle [11], as well as specimens from the Birmingham region (United Kingdom) described in [10] (Figure 1).
DNA extraction, sequencing, and culture collection information was described previously [7]. For each cultured species, genomic DNA was extracted from 150 females obtained from a pure culture (see above), using a QIAamp DNA Micro kit (Qiagen, Venlo, The Netherlands) with modifications as described here [12]. Illumina sequencing libraries were generated and sequenced commercially on an Illumina NovaSeq 6000 sequencing system. Short Illumina reads were assembled in SPAdes v.3.15.5 [13] as follows: metaspades.py -t 24 -m 240 -1 {name}_R2_001.fastq -o {name}. Full-length cytochrome c oxidase subunit 1 (COX1) sequences were found using a local NCBI BLAST search [[14](#B14-animals-15-00357)]. Th sequences previously generated by us were used (GenBank accession IDs: [OR640973-OR640976](OR640973:OR640976)). We also used other GenBank species as detailed in the text and figures. We used Mesquite version 3.81 [[15](#B15-animals-15-00357)] to create sequence alignments, translate nucleotide matrices into protein sequences, and store Nexus matrices for both protein and nucleotide data. Genetic distances were calculated in PAUP v.4a168 [[16](#B16-animals-15-00357)] as follows: begin paup; dset distance = p; savedist format = tabtext undefined = asterisk file = 1_p_distances.tab; dset distance = k2p; savedist format = tabtext undefined = asterisk file = 2_k2p_distances.tab; end. Phylogenetic trees were inferred in IQ-TREE v. 2.3.4 [[17](#B17-animals-15-00357)] using amino acid data (command: iqtree2 -s ipf --seqtype AA -T AUTO -m MFP -alrt 1000 -bb 1000 -safe --prefix ipf -spp partitions.txt -nt 8 -m MFP+MERGE -rcluster 10 -alrt 1000 -bb 1000 -safe --prefix ipf is the input file name (a nexus matrix) and ipf.
Images were captured from multiple focal planes and assembled using Helicon Focus Pro 7.6.4 (algorithm B, occasionally A), with subsequent manual editing (retouching) of any misaligned regions. Partially overlapping images were merged into a full panorama in Adobe Photoshop 22.2.0. Line drawings were created in Photoshop 22.2.0 using microphotographs as the background. The background images were taken using a Euromex Color HD-Ultra camera attached to a Bioptic C-400 microscope (Bioptic, Moscow, Russian Federation) equipped with bright field and differential interference contrast (DIC) optics. Publication-quality microphotographs were captured using an Axio Imager A2 compound microscope (Carl Zeiss, Oberkochen, Germany) equipped with DIC and phase contrast optics, along with an Axiocam 506 color digital camera (Carl Zeiss, Oberkochen, Germany). For scanning electron microscopy (SEM) imaging, alcohol-preserved mites were dried in a JFD 320 freeze dryer (JEOL, Tokyo, Japan), coated with gold, and scanned using a TESCAN Mira3 LMU SEM microscope. Specimens used for SEM were not preserved.
In the descriptions, idiosomal chaetotaxy follows the system outlined in reference [18]; the terminology of coxal setae follows reference [19]. For appendages, chaetotaxy and solenidiotaxy follow Grandjean’s system for palps [20] and legs [21]. Designations of tarsal dorsoapical setae on legs III–IV follow reference [22]. All measurements are given in micrometers (µm).
3. Results
3.1. GenBank Data Analysis
Maximum likelihood phylogenetic trees were inferred using protein sequences with the mtART+I+G4 model (Figure 2A) and nucleotide sequences with a codon model (Figure 2B). The genus Thyreophagus was recovered as monophyletic. The internal relationships within Thyreophagus were resolved as two sister groups: (Th. tauricus, Th. corticalis) and (Th. “entomophagus” from China [NC 066986], Th. calusorum) (Figure 2). However, the placement of Thyreophagus entomophagus remained unresolved. In the amino acid phylogeny, T. entomophagus was recovered as sister to the Th. “entomophagus” (China) + Th. calusorum clade, whereas in the nucleotide phylogeny, it was sister to the Th. tauricus + Th. corticalis clade. In both phylogenetic trees, the remaining taxa morphologically assigned to the family Acaridae (Tyrophagus, Sancassania, and Aleuroglyphus) formed a monophyletic group, corresponding to the subfamilies Tyrophaginae and Acotyledonini (Figure 2). However, traditional morphological classifications place Sancassania (Acotyledonini) within the subfamily Rhizoglyphinae, which is typically defined by the presence of smooth dorsal setae. Interestingly, Fagacarus (Rhizoglyphinae) possesses strongly pectinate setae [23,24], while Sancassania exhibits sparse pectinations on some of its setae. These observations suggest that pectinated dorsal setae, a character subject to evolutionary lability, is unreliable for delimiting major groups within Astigmata. In contrast, the presence of the tarsal seta aa—although considered plesiomorphic—appears to be a more robust character for defining lineages within Astigmata. In addition, our trees indicate that the GenBank sequence originally identified as Rhizoglyphus robini (NC 038058) actually belongs to the genus Sancassania (=Caloglyphus) (Figure 2).
3.2. Morphological Description
3.2.1. Genus Thyreophagus Rondani, 1874
Thyreophagus Rondani, 1874: 67 (=Moneziella Berlese, 1897; Monetiella Berlese, 1897;
Monieziella Berlese, 1897; Fumouzea Zachvatkin, 1953; Michaelopus Fain and Johnston, 1974).
Type species: Thyreophagus entomophagus (Laboulbène and Robin, 1862), by monotypy.
3.2.2. Thyreophagus entomophagus (Laboulbène and Robin, 1862)
Acarus entomophagus Laboulbène, 1852: LIV (nomen nudum).
Tyroglyphus entomophagus Laboulbène and Robin, 1862: 321, Pl. 10 (female, homeomorphic male; types lost [10]).
Material. Cultures were obtained from a European biocontrol company and maintained on a mixture of yeast and bran in specialized rearing units by VAK. Cultures were started on 20 November 2021; specimens were harvested on # COX1 barcoding sequence GenBank Id: OR640974 (culture PBK 20-0101-199.SM38, from which the neotype was designated).
Type material. Neotype (female) same data, deposited at the Royal Belgian Institute of Natural Sciences, Brussels, Belgium.
Female (Figure 3A,B, Figure 4, Figure 5E,F, Figure 6, Figure 7, Figure 9D–I, Figure 10 and Figure 18A,D,E,H). Idiosoma slightly elongate, 400–430 × 210–225 (n = 20), 1.9 times longer than wide. Idiosomal cuticle smooth. Subcapitular setae (h) long, widened basally; palp tibial setae (a), lateral dorsal palp tibial setae (sup), dorsal palp tarsal seta (cm) filiform; supracoxal seta elcp present; terminal palp tarsal solenidion ω short; external part of terminal eupathidium ul” dome-shaped; terminal eupathidium ul’ small, rounded. Prodorsal shield 73–80 long, 75–84 wide, nearly as long as wide, with setae vi (situated at anterior part of shield, alveoli of vi noticeably separated—the distance between them almost equal to their width), rounded anterolateral incisions, and elongate midlateral incisions (insertion points of setae ve). Prodorsal shield smoothly punctate (with small cells), without distinct lineate pattern (cells may be slightly elongated or rounded in the posterior part) and with several (3–5) curved longitudinal lines in posterior part. Grandjean’s organ (GO) with seven membranous finger-shaped processes. Supracoxal setae (scx) smooth, sword-shaped, widened and flattened, tapering at tip. Idiosomal setae (vi, c_p_, d2, e2, h1, h3, ps3) smooth, filiform and short, setae se and h2 twice as long as other idiosomal setae, smooth and filiform; opisthosomal gland openings between setal bases e2 and d2. Four pairs of fundamental cupules (ia, im, ip and ih) present. With a pair of additional cuticular pores between setae h1. Ventral idiosoma with four pairs of coxal setae (1a, 3a, 4a and 4b) and one pair of genital setae (g). Shape of coxal sclerites as in Figure 3B, Figure 6B and Figure 7B. Genital region situated between coxal fields III and IV; genital valves form an inverted Y; epigynal and medial apodemes well developed. Diameter of genital papillae approximately 0.4–0.5 the length of coxal and genital setae. Anal opening terminal. Copulatory tube present, situated anterodorsally to anus, with distinct opening. Inseminatory canal of spermatheca long, slender tube-like, leading from copulatory opening to spermatheca, uniformly wide. Base of spermatheca wide, bell-shaped, with a distinct vestibule. Paired sclerites of efferent ducts elongate, their length approximately 1/2 the length of spermatheca base, with short stem.
Legs short, all segments free. Trochanters I–III each with long, filiform seta, pR I–II, sR III; trochanter IV without setae. Femoral setation 1-1-0-1; setae vF I–II and wF IV long, filiform. Genual setation 2-2-0-0; setae mG and cG I–II long, filiform; seta nG III absent. Tibial setation 2-2-1-1; setae hT I-II spiniform, shorter than gT; setae gT I–II and kT III–IV elongate, somewhat spiniform. Tarsal setation 8-8-8-8; pretarsi consist of hooked empodial claws attached to short paired condylophores. Tarsi I and II with setae ra, la, f and d filiform, e, u, v spiniform, p and q represented by very small remnants, s flattened, button-shaped or minute spiniform (button-shaped in neotype); setae wa absent. Tarsi III and IV with setae f, d, r, w filiform, e and s spiniform, u and/or v flattened, button-shaped, p and q represented by very small remnants. Solenidion ω_1_ on tarsus I cylindrical, slightly arched, with a slight narrowing before apical widening. Solenidion ω_1_ on tarsus II simple, cylindrical, with clavate apex, nearly straight. Solenidion ω_2_ on tarsus I shorter than ω_1_, cylindrical, with rounded apex, slightly widened distally, situated slightly anteriad ω_1_. Solenidion ω_3_ on tarsus I cylindrical, with rounded tip, subequal to ω_1_, longer than ω_2_. Famulus (ε) of tarsus I wide, spiniform, with broadly rounded apex, widest at middle. Solenidia ϕ of tibiae I–III elongate, tapering, well extending beyond apices of respective tarsi with ambulacra; solenidion ϕ IV shorter than tarsus IV (with ambulacra). Solenidia σ’ and σ” on genu I elongate, tapering, σ” longer than σ’; σ” slightly not reaching bases of ϕ I. Solenidion σ on genu II more than 6–7 times longer than its width, with rounded tip. Solenidion σ of genu III absent.
Male (n = 5) (Figure 3C,D, Figure 5A–D,G, Figure 8, Figure 9A–C,L–J and Figure 11). Idiosoma slightly elongate, 300–360 × 180–200, 1.7–1.8 times longer than wide. Idiosomal cuticle smooth. Gnathosoma as in female. Prodorsal shield 73–82 long, 73–80 wide, nearly as long as wide, with setae vi, incisions and ornamentation as in female. Grandjean’s organ (GO) and supracoxal seta (scx) as in female. All idiosomal setae smooth and filiform, setae se longer and wider that other setae; setae cp, e2 and h3 longer than vi, d2, h1, h2. Two pairs of fundamental cupules (ia and ih) present, im and ip not observed. With a pair of of additional cuticular pores between setae h1. Opisthonotal shield smoothly punctate; ventral part extends to anal suckers. Ventral idiosoma with four pairs of coxal setae (1a, 3a, 4a and 4b) and one pair of genital setae (g). Posterior region of idiosoma with a large rounded lobe extending posteriorly (45–48 × 80–86, 1.7–1.8 times longer than wide). Shape of coxal sclerites as in Figure 3D and Figure 8B. Genital region between coxisternal fields IV; genital capsule rounded; aedeagus short, not protruding beyond anterior edge of genital capsule. Diameter of genital papillae approximately 0.4–0.5 the length of coxal and genital setae. Anal suckers rounded in outline. Setae ps1–3 very short. Figure 8Thyreophagus entomophagus (Laboulbène and Robin, 1862), PBK20-0101-199.SM38, male, not neotype, DIC images: (A)—dorsal view; (B)—ventral view. Figure 9Thyreophagus entomophagus (Laboulbène and Robin, 1862), PBK20-0101-199.SM38, not neotype, DIC images: (A) male prodorsal shield; (B,C) male gnathosoma; (D) spermatheca; (E) female leg I, dorsal view; (F) female tarsus I, ventral view; (G) female leg I, dorsal view; (H) female tarsi III–IV, ventral view; (I) female tarsi III–IV, dorsal view; (J) male tarsi III–IV, ventral view; (K) male tarsus IV, dorsal view; (L) male tarsus I, ventral view. Figure 10Thyreophagus entomophagus (Laboulbène and Robin, 1862), PBK20-0101-199.SM38, female, not neotype, SEM images: (A) posterior part of idiosoma, dorsal view; (B) posterior part of idiosoma, lateral view; (C) genital area, ventral view; (D) supracoxal seta; (E) Grandjean’s organ; (F) leg I, dorsal view; (G) tarsus II, ventral view; (H,I) tarsi III, lateral and ventral views. Figure 11Thyreophagus entomophagus (Laboulbène and Robin, 1862), PBK20-0101-199.SM38, male, not neotype, SEM images: (A) dorsal view; (B) ventral view; (C) propodosoma, dorsal view; (D) ano-genital region, ventral view.
Legs I–III as in female, except solenidion ω_3_ on tarsus I very short, truncated. Trochanter and genu IV without setae, femur IV with wF IV long, filiform, tibia IV with kT IV elongate, somewhat spiniform. Tarsus IV with 7 or 8 setae (one v or u may be absent), of them, f, r, w filiform, d and e represented by suckers, s spiniform, u and v flattened, button-shaped, p and q represented by very small remnants. Solenidion ϕ on tibia IV short and wide.
Heteromorphic deutonymph. Absent.
The Birmingham population. The currently used species concept of Th. entomophagus was established by A. Fain in 1982 [10], who described specimens from the United Kingdom (Birmingham) without designating a neotype. We re-examined two females from the Birmingham sample and compared them with females from our culture of Thyreophagus entomophagus (PBK20-0101-199.SM38). These two were conspecific. Particularly, we note the identical shapes of u and v III, which are flattened and button-shaped (Figure 18F) and slightly elongated or rounded cells on the posterior part of the prodorsal shield (Figure 18B), without a lineate pattern; A. Fain [10] probably mistook the slightly elongated punctations for a lineate pattern, but they are markedly different from the lineate pattern seen in Th. holda sp. n. (Figure 16C). We also note that ventroterminal setae on tarsi III–IV, u and v III–IV were originally figured by Fain [10] as spiniform, but actually they are flattened and button-shaped structures. The shape of the spermatheca base in the specimens described by A. Fain [10] agrees well with that of all the specimens we studied. In some cases, its width may change, but this is due to deformation during specimen preparation. We were unable to find any noticeable widening of the inseminatory canal, as depicted in A. Fain [10]. Heteromorphic deutonymphs are unknown for the Birmingham sample.
Diagnosis. Adults of Th. entomophagus differ from all other species (except Th. hobe Klimov et al., 2023) by the presence of flattened, button-shaped setae u and v on tarsus III (vs. spiniform in other species). Thyreophagus entomophagus differs from Th. hobe by the presence of filiform w III (minute spiniform in Th. hobe) and the presence of wF IV (absent in Th. hobe). Also, Th. entomophagus differs from all other species by the bell-shaped base of spermatheca (vase-shaped, narrow, parallel-sided, broad-arc-shaped, or reduced in other species).
For Th. entomophagus, three subspecies have been described: Th. entomophagus nominalis Kadzhaya, 1973, Th. entomophagus ponticus Kadzhaya, 1973 [25] and Th. entomophagus italicus Vacante, 1989 [26]. Subsequently, Th. entomophagus ponticus and Th. entomophagus italicus were elevated to the rank of species [6].
Thyreophagus ponticus Kadzhaya, 1973 has a well-developed opisthosomal projection in males, as in Th. entomophagus. However, in females of Th. ponticus, setae h3 are only slightly longer than ps_3_ (more than twice as long in Th. entomophagus), and the anal opening is approximately equal to the length of setae h3 (more than twice as long as the anal opening in Th. entomophagus). Thyreophagus ponticus is insufficiently described and is considered a species inquirenda [6].
Thyreophagus italicus Vacante, 1989 differs from Th. entomophagus by the shape of the base of the spermatheca, which is vase-shaped, narrow, with constriction (bell-shaped, wide, without constriction in Th. entomophagus), very short setae w III (long, filiform in Th. entomophagus), spiniform setae u and v III, the same length as s III (flattened, button-shaped, more than twice as short as s III in Th. entomophagus).
Thyreophagus entomophagus nominalis is an insufficiently described taxon, considered a species inquirenda [6]. It differs from Th. entomophagus by having subequal setae h1 and ps1 in females [25] (h1 are distinctly longer than ps1 in Th. entomophagus).
Neotype. The need for typification of Thyreophagus entomophagus through neotype designation is based on the following: (i) several morphologically similar species are known (Th. holda sp. n., Th. leclercqi), but the differences between these nominal species are not well understood and may represent either interspecific or intraspecific variations; sequence data are lacking for these species; (ii) specimens from China identified as Thyreophagus “entomophagus” [27] (without morphological description) show uncorrected genetic distances (p-distance) of 18.09% at nucleotide level and 6.36% at the amino acid level (NC_066986 vs. OR640974), indicating that two genetically distinct species are involved. This suggests that previous re-descriptions of Thyreophagus entomophagus may not suffice for accurate identification of this species and a neotype is needed. Our proposed neotype will provide stability in the usage of the name Thyreophagus entomophagus, facilitating confident identification based on both morphology and DNA sequence data. To standardize the current usage of the name Thyreophagus entomophagus, we propose designating a neotype (female, accession #) from a commercial culture PBK20-0101-199.SM38 used for the industrial rearing of predatory mites as described here using light microscopy, SEM, and COX1 sequence data.
3.2.3. Thyreophagus holda Klimov, Kolesnikov, Merckx, Duarte et Vangansbeke sp. n.
Thyreophagus entomophagus Fain et al., 2000: 154, Figure 1, Figure 2, Figure 3, Figure 4, Figure 5, Figure 6, Figure 7, Figure 8 and Figure 9 (heteromorphic deutonymph; misidentification).
Material. Adults and heteromorphic deutonymphs—culture 1, started from adult specimens collected in a sparrow nest in the suburban district of Berlin (Berlin Dahlem), W. Knülle [11].
Type material. Holotype (female, slide 29-90a, recognizable by unique shape on slide) and paratypes (six females, five males and four heteromorphic deutonymph) same data (slides 29-89, 29-89a, 29-90, 29-90a).
Depository. The holotype and 15 paratypes are deposited in the Royal Belgian Institute of Natural Sciences, Brussels, Belgium.
Etymology. Holda is a popular figure in German folklore.
Female (n = 7) (Figure 12, Figure 14A–D,F,G, Figure 16A,B, Figure 17A,B,D–G,J and Figure 18B,E,I). Idiosoma slightly elongate, 580–800 × 300–400, 1.7–2.0 times longer than wide. Idiosomal cuticle smooth. Subcapitular setae (h) long, widened basally; palp tibial setae (a), lateral dorsal palp tibial setae (sup), dorsal palp tarsal seta (cm) filiform; supracoxal seta elcp present; terminal palp tarsal solenidion ω short; external part of terminal eupathidium ul” dome-shaped; terminal eupathidium ul’ small, rounded. Prodorsal shield 100–110 long, 110–112 wide, 1.0–1.1 times longer than wide, with setae vi (situated at anterior part of shield, alveoli noticeably separated—distance between them almost equal to their width), rounded anterolateral incisions, and elongate midlateral incisions (insertion points of setae ve). Prodorsal shield smoothly punctate (small cells), with lineate cells in posterior part and with several (3–5) curved longitudinal lines in posterior part. Grandjean’s organ (GO) with seven membranous finger-shaped processes. Supracoxal setae (scx) smooth, sword-shaped, widened and flattened, tapering at tip. Idiosomal setae (vi, cp, d2, e2, h1, h3, ps3) smooth, filiform and short, setae se and h2 twice as long as other idiosomal setae, smooth and filiform; opisthosomal gland openings between setal bases e2 and d2. Four pairs of fundamental cupules (ia, im, ip and ih) present. With a pair of additional cuticular pores between setae h1. Ventral idiosoma with four pairs of coxal setae (1a, 3a, 4a and 4b) and one pair of genital setae (g). Shape of coxal sclerites as in Figure 12B. Genital region situated between coxal fields III and IV; genital valves form an inverted Y; epigynal and medial apodemes well-developed. Diameter of genital papillae approximately 0.4–0.5 the length of coxal and genital setae. Anal opening terminal. Copulatory tube present, situated anterodorsally to anus, with developed opening. Inseminatory canal of spermatheca long, slender tube-like, leading from copulatory opening to spermatheca, slightly widened at junction with base of spermatheca. Base of spermatheca wide, bell-shaped, with a distinct vestibule. Paired efferent ducts elongated, their length is approximately 1/2 the length of the base of spermatheca, with short stem.
Legs short, all segments free. Trochanters I–III each with long, filiform seta, pR I–II, sR III; trochanter IV without setae. Femoral setation 1-1-0-1; setae vF I–II and wF IV long, filiform. Genual setation 2-2-0-0; setae mG and cG I–II long, filiform; seta nG III absent. Tibial setation 2-2-1-1; setae hT I-II spiniform, shorter than gT; setae gT I–II and kT III–IV elongate, somewhat spiniform. Tarsal setation 8-8-8-8; pretarsi consist of hooked empodial claws attached to short paired condylophores. Tarsus I and II with setae ra, la, f and d filiform, e, u, v spiniform, p and q represented by very small remnants, s flattened, button-shaped or minute spiniform; setae wa absent. Tarsus III and IV with setae f, d, r, w filiform, e, s, u and v spiniform (u and v slightly less than s), p and q represented by very small remnants. Solenidion ω_1_ on tarsus I cylindrical, with clavate apex, in front of which there is a slight narrowing, curved; solenidion ω_1_ on tarsus II simple, cylindrical, with clavate apex, not bent. Solenidion ω_2_ on tarsus I shorter than ω_1_, cylindrical, with rounded apex, slightly widened distally, situated slightly distal to ω_1_. Solenidion ω_3_ on tarsus I cylindrical, with rounded tip, subequal to ω_1_, longer than ω_2_. Famulus (ε) of tarsus I wide, spiniform, with broadly rounded apex, widest at middle. Solenidia ϕ of tibiae I–III elongate, tapering, extending well beyond apices of respective tarsi with ambulacra; solenidion ϕ IV shorter, shorter than tarsus IV (with ambulacra). Solenidia σ’ and σ” on genu I elongate, tapering, σ” longer than σ’, σ” slightly not reaching bases of ϕ I. Solenidion σ on genu II more than 6–7 times longer than its width) with rounded tip. Solenidion σ of genu III absent.
Male (n = 5) (Figure 13A,B, Figure 14E, Figure 16C,D and Figure 17C,H,I). Idiosoma slightly elongate, 430–470 × 250–300, 1.6–1.7 times longer than wide. Idiosomal cuticle smooth. Gnathosoma as in female. Prodorsal shield 86–95 long, 90–98 wide, 1.0–1.1 times longer than wide, with setae vi, incisions and ornamented as in female. Grandjean’s organ (GO) and supracoxal seta (scx) as in female. All idiosomal setae smooth and filiform, setae se linger and widener that other, setae cp, e2 and h3 longer than vi, d2, h1, h2. Two pairs of fundamental cupules (ia and ih) present, im and ip not observed. Between setae h1, there is a pair of additional cuticular pores. Opisthonotal shield smoothly punctate; ventral part extends to anal suckers. Ventral idiosoma with four pairs of coxal setae (1a, 3a, 4a and 4b) and one pair of genital setae (g). Posterior region of idiosoma with a large rounded lobe extending the body backwards (45–50 × 78–85, 1.5–1.8 times longer than wide). Shape of coxal sclerites on Figure 13B. Genital region between coxisternal fields IV; arms of genital capsule rounded; aedeagus short, not protruding beyond anterior edge of genital capsule. Diameter of genital papillae approximately 0.4–0.5 the length of coxal and genital setae. Anal suckers rounded in outline. Setae ps1–3 very short.
Legs I-III as female, except solenidion ω_3_ on tarsus I very short, truncated. Trochanter and genu IV without setae, femur IV with wF IV long, filiform, tibia IV with kT IV elongate, somewhat spiniform. Tarsus IV with 8 setae, of them, f, r, w filiform, d and e represented by suckers, s, e, u and v spiniform (u and v slightly less than s), p and q represented by very small remnants. Solenidion ϕ on tibia IV short and wide.
Heteromorphic deutonymph (n = 4) (Figure 13C,D, Figure 15, Figure 16E,F and Figure 17K–M). Body ovoid, 1.3–1.4 times longer than wide, widest in sejugal region; idiosomal length 240–250 width 170–190. Gnathosoma short, subcapitulum and palps fused, bearing palpal solenidia (ω) apically and filiform apicodorsal setae (sup); setae h absent (their positions marked by somewhat refractile spots), setae cm absent.
Dorsum. Idiosoma smoothly punctate; distinct linear pattern present on anterior and lateral sides of prodorsal sclerite and hysterosomal shield. Apex of propodosoma anterior to anterior border of prodorsal sclerite, with apical internal vertical setae (vi) (bases separated) and a pair of band-like sclerites coalescing anteriorly. A pair of lateral, widely separated ocelli (distance 68–70) present on prodorsum; lenses and pigmented spots present; maximum diameter of lenses 9–10. External vertical setae (ve) absent; external scapular setae se situated below lenses; internal scapular setae (si) distinctly posterior external scapulars (se). Supracoxal setae of legs I (scx) filiform, with extended base, situated below si and anterolaterad of ocelli. Sejugal furrow well developed. Prodorsal sclerite 70–75, hysterosomal shield 158–173, ratio hysterosomal shield/prodorsal sclerite length = 2.2–2.3. Hysterosoma with 11 pairs of simple, filiform setae on hysterosomal shield (c1, c2, cp, d1, d2, e1, e2, f2, h1, h2, h3), setae h3 distinctly longer than other setae. Opisthonotal gland openings (gla) situated ventrally on hysterosomal shield, slightly posterior setae c3, much closer to ventral seta c3 than to dorsolateral seta cp (in one specimen approximately equidistant from setae c3 and cp). Of four fundamental pairs of cupules, three pairs were observed: ia posteriomediad setae c2, im ventral, laterad trochanters IV and ih ventral, laterad posterior sides of attachment organ.
Venter. Coxal fields sclerotized, smoothly punctate. Anterior apodemes of coxal fields I fused forming sternum; sternum not reaching posterior border of sternal shield by distance exceeding its length. Posterior border of sternal shield not sclerotized. Anterior apodemes of coxal fields II curved medially. Posterior apodemes of coxal fields II weakly developed, thin, curved medially. Sternal and ventral shield contiguous. Anterior apodemes of coxal fields III free, connected by thin transverse sclerotization. Posterior medial apodeme present in area of coxal fields IV, well-separated from anterior apodemes IV and genital opening. Posterior apodemes IV absent. Subhumeral setae (c3) filiform, situated on ventral surface between legs II and III, near region separating sternal and ventral shields. Coxal setae 1a and 3a reduced, represented by minute structures. Setae 4b, g filiform; 4a in form of small, rounded conoids. Genital region in posterior portion of coxisternal fields IV; genital opening elongate, with two pairs of genital papillae within genital atrium; genital papillae two-segmented, with rounded apices. Coxal setae (4b) situated at free ends of anterior coxal apodemes IV; genital setae (g) laterad genital opening. Attachment organ posterior to coxal fields IV. Anterior suckers (ad3) round, median suckers (ad1+2) distinctly larger, with paired vestigial alveoli (not situated on common sclerite); pair of small refractile spots anterolaterad median suckers (ps3); lateral conoidal setae of attachment organ (ps2) situated distinctly posterior to line joining centers of median suckers, slightly anteriad conoidal setae (ps1); anterior and posterior lateral and posterior median cuticular conoids well developed; anus situated between anterior suckers (ad3).
Legs. Legs elongate, all segments free. Trochanters I–III each with long, filiform seta, pR I–II, sR III. Femoral setation 1-1-0-1; setae vF I–II and wF IV long, filiform. Genual setation 2-2-0-0; setae mG and cG I–II filiform, seta nG III absent. Tibial setation 2-2-1-1; setae hT I somewhat spiniform; setae gT I-II filiform; setae hT II spiniform, setae gT longer than hT; setae kT III-IV filiform, without a prong. Tarsal setation 8-9-8-8. All pretarsi consisting of hooked empodial claws attached to short paired condylophores. Tarsus I with setae ra, la, p, q, e, f foliate; seta d filiform, its base at level of ra and la; seta s represented by alveolus; setae wa, aa and ba I absent; tarsus II similar to tarsus I except seta ba present, filiform, situated close to ω_1_. Tarsus III with setae w, r, s, p, q, e, f and d smooth; all setae, except for d III, more or less foliate; seta d longer than leg. Tarsus IV similar to tarsus III, except seta r filiform and w filiform with a distinct prong. Solenidia ω_1_ on tarsi I–II cylindrical, with slightly clavate apices; ω_3_ on tarsus I longer than ω_1_, with rounded apex, situated slightly distal to ω_1_; ω_1_ and ω_3_ separated by bulbous famulus (ε); solenidion ω_2_ of tarsus I slightly expanding apically, situated somewhat more basal and posterior to ω_1_ + ε + ω_3_ group; solenidia φ of tibiae I–III elongate, tapering; φ I and III longer than tarsus I and III, respectively; solenidion φ II shorter than tarsus II; solenidion φ IV short; solenidion σ of genu I elongate, slightly tapering, nearly reaching tip of tibia I; solenidion σ of genu II much shorter, cylindrical, not reaching midlength of tibia II; solenidion σ of genu III absent.
Diagnosis. Adults of Th. holda are very similar to those of Th. entomophagus, but differ by the following: apical tarsal setae u and v III-IV normally developed, spiniform (vs. flattened and button-shaped in Th. entomophagus) (Figure 18G,I vs. Figure 18D–F,H); prodorsal shield with a lineate cells in its posterior part (vs. rounded or slightly elongated in Th. entomophagus) (Figure 18C vs. Figure 18A,B); larger body measurements (females 580–800 and males 430–470 vs. females 400–430 and males 300–360 in Th. entomophagus). The new species forms heteromorphic deutonymphs (vs. lacking in Th. entomophagus). Figure 12Thyreophagus holda sp. n., female, holotype: (A) dorsal view; (B) ventral view. Figure 13Thyreophagus holda sp. n., paratypes: (A) male, dorsal view; (B) male, ventral view; (C) heteromorphic deutonymph, dorsal view; (D) heteromorphic deutonymph, ventral view. Figure 14Thyreophagus holda sp. n., holotype (A–D,F) and paratypes (E,G): (A) female leg I, posterior view; (B) female leg II, anterior view; (C) female leg III, anterior view; (D) female tarsus III, posterior view; (E) male leg IV, anterior view; (F,G) spermathecae. Figure 15Thyreophagus holda sp. n., heteromorphic deutonymph, paratype: (A) leg I, antiaxial view; (B) leg II, anterior view; (C) leg III, anterior view; (D) leg IV, anterior view; (E) gnathosoma, ventral view. Figure 16Thyreophagus holda sp. n., phase contrast images: (A) female holotype; (B) female paratype; (C,D) male paratypes; (E,F) heteromorphic deutonymph paratypes. Figure 17Thyreophagus holda sp. n., holotype (A,F,J) and paratypes (B–E,I,H–M), phase contrast images: (A,B) female prodorsal shields; (C) male prodorsal shield; (D) female leg I, anterior view; (E) female leg II, posterior view; (F) female leg III, posterior view; (G) female leg III, anterior view; (H) male leg III, posterior view; (I) male leg I, anterior view; (J) spermatheca; (K) heteromorphic deutonymph leg I, anterior view; (L) heteromorphic deutonymph leg II, anterior view; (M) heteromorphic deutonymph legs III–IV. Figure 18Thyreophagus entomophagus (Laboulbène and Robin, 1862) (A,B,D–F,H), PBK20-0101-199.SM38, not neotype (A,D,E,H), Birmingham specimens (B,F) and Th. holda sp. n. (C,G,I), phase contrast images (A–D,F,G), DIC images (E) and lineal drawings (H,I) distinctive morphological characteristics: (A–C) punctated posterior part of female prodorsal shields; (D–F) female tarsi III with flattened, button-shaped setae v, anterior view; (G) female tarsi III with spiniform setae v, anterior view. Abbreviations: a—rounded and elongated cells; b—lineate cells; s, v, q—tarsal ventral setae.
Heteromorphic deutonymphs of Th. holda sp. n. are similar to Th. leclercqi (Fain, 1982) [28] and Th. australis (Clark, 2009) [29] in terms of their ovoid bodies, which are 1.3–1.4 times longer than they are wide, compared to the elongate bodies (more than 1.7 times longer than they are wide) in other species. Th. holda differs from Th. leclercqi by their short setae hT I, which are more than 2 times shorter than gT I (less than 2 times shorter in Th. leclercqi). Th. holda differs from Th. australis by their setae kT III, which lack a prong (with a distinct prong in Th. australis), and the absence of setae wa I-II (present in Th. australis).
Based on Figure 1 in reference [11], Barbosa et al. [30] and Klimov et al. [5] suggested that one of the diagnostic character states of Th. holda sp. n. heteromorphic deutonymphs is the position of the opisthonotal gland openings, which are approximately equidistant from setae c3 and cp. However, our study showed that only one specimen of Th. holda sp. n. shows this character state, while the remaining specimens from the same culture have the usual arrangement of these openings (gla is much closer to ventral seta c3 than to dorsolateral seta cp). Based on these findings, we suggest that this character is variable and should not be considered diagnostic for Th. holda sp. n.
4. Discussion
The mite Thyreophagus entomophagus is an economically important species widely used as factitious prey in the industrial rearing of phytoseiid mites for biocontrol applications [1,5,6,7,31]. This is a cosmopolitan species [32] that has been reported in the literature from various habitats: entomological collections [9,10,33]; flour [34,35,36,37,38,39,40,41]; dry Spanish fly Lytta vesicatoria harvested for medicinal use [35]; vanilla pods and saffron for medicinal use [42]; medicinal plants, including ergot and cardamom; stimulants; spices; food products and fodders (rye and wheat bran) [33]; poultry meal [43]; on ergot of rye [42]; in bird nests [33,43]; bracket fungi Fomitopsis betulina [44]; and various materials in beehives of Apis mellifera [45,46] in association with scale insects [36,47,48], coccids [48], and trogid beetles [49].
However, the reliability of this name may be compromised due to several factors: the absence of original type specimens, difficulties in interpreting the classical species diagnosis by A. Fain [10], which is not entirely accurate with respect to actual specimens (e.g., the shape of ventroterminal setae u and v III–IV), the presence of closely related species such as Th. holda and Th. leclercqi, and the misidentified and divergent GenBank sequence that lacks accompanying morphological information. As a result, previous reports in the literature require verification. Here, to standardize the usage of the name Thyreophagus entomophagus, we propose designating a neotype for which both detailed morphological and sequence data are available.
Our research confirms that Th. entomophagus lacks the heteromorphic deutonymph stage (Figure 1), a trait consistently observed in laboratory populations cultured independently across multiple labs and biocontrol facilities (our observations). Earlier reports from Germany indicated the presence of heteromorphic deutonymphs [11], but our study did not corroborate this, suggesting that the German population represents a different species, Th. holda sp. n. The absence of heteromorphic deutonymphs is advantageous for mass production, as it simplifies the life cycle by eliminating an energetically costly developmental stage. These deutonymphs are also heavily sclerotized, making them less palatable to predators. However, Th. entomophagus remains a sexual species, which is a less desirable trait for mass production since asexual species reproduce more quickly, generating only females. The only known species that is both asexual and lacks the heteromorphic deutonymph stage is Th. plocepasseri [22].
Our preliminary phylogenetic analysis suggests that the two traits important for biocontrol—asexual reproduction and direct development (i.e., absence of the heteromorphic deutonymph stage)—are derived, having emerged after the most recent common ancestor of Thyreophagus (Figure 2). While speculative, asexual reproduction may provide an advantage in subcortical habitats like fallen branches, where predator activity is low and the “Red Queen” dynamics [50,51] are absent. The lack of heteromorphic deutonymphs might also confer an advantage in vertebrate nests, where animals can transport branches, facilitating long-distance dispersal. Short-distance dispersal, such as within a nest, can be accomplished by the mites themselves, particularly the more mobile immature stages. In contrast, inseminated females tend to burrow into tunnels where they produce large numbers of eggs as slow-moving, ovigerous individuals.
Our study not only standardizes the nomenclature of Thyreophagus entomophagus, a species widely utilized in the rearing of predatory mites, but also proposes identifying species that combine advantageous traits, such as asexual reproduction and a direct life cycle, for further bioprospecting efforts.
5. Conclusions
The mite Thyreophagus entomophagus is a cosmopolitan mite species with significant economic importance in biocontrol applications. Its diverse habitat range and frequent association with various food products and environments suggest its great adaptability. However, the taxonomic status of this species name is questionable due to inconsistencies in historical species identifications, the absence of original type specimens, and misidentified GenBank sequences. To address these issues and to standardize the nomenclature, we propose the designation of a neotype with both morphological and genetic data available.
Our research shows that Th. entomophagus lacks the heteromorphic deutonymph stage, an advantageous trait for mass production, streamlining its life cycle. Nonetheless, this is a sexual species, a feature less desirable for mass-rearing programs. Phylogenetic evidence suggests that asexuality and the absence of heteromorphic deutonymphs are derived traits, likely advantageous in specific ecological niches like subcortical habitats or vertebrate nests. These findings offer important insights into the evolutionary traits that improve the effectiveness of Th. entomophagus and related species in biocontrol settings. Our study specifically suggests the need for additional bioprospecting efforts to find new candidate species for biocontrol applications that exhibit both asexual reproduction and the absence of heteromorphic deutonymphs.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Fidgett M.J. Stinson C.S.A. A Method of Rearing Amblyseius Predatory Mites Using Thyreophagus entomophagus as Prey Patent CA 2658292 A 17February 2008 Available online: https://patentimages.storage.googleapis.com/e 7/4f/61/612fa 7977 fb 51a/CA 2658292 A 1.pdf(accessed on 10 July 2024)
- 2Steiner M.Y. Goodwin S. Wellham T.M. Barchia I.M. Spohr L.J. Biological studies of the Australian predatory mite Typhlodromips montdorensis (Schicha) (Acari: Phytoseiidae), a potential biocontrol agent for western flower thrips, Frankliniella occidentalis (Pergande) (Thysanoptera: Thripidae)Aust. J. Entomol.20034212413010.1046/j.1440-6055.2003.00343.x · doi ↗
- 3Nomikou M. Janssen A. Schraag R. Sabelis M.W. Phytoseiid predators suppress populations of Bemisia tabaci on cucumber plants with alternative food Exp. Appl. Acarol.200227576810.1023/A:102155942134412593512 · doi ↗ · pubmed ↗
- 4Bolckmans K.J.F. van Houten Y.M. Mite Composition, Use Thereof, Method for Rearing the Phytoseiid Predatory Mite Amblyseius Swirskii, Rearing System for Rearing Said Phytoseiid Mite and Methods for Biological Pest Control on a Crop WO Patent WO/2006/05755201 June 2006 Available online: https://patentscope.wipo.int/search/en/detail.jsf?doc Id=WO 2006057552(accessed on 10 July 2024)
- 5Klimov P.B. Kolesnikov V.B. Demard E.P. Stinson C.S.A. Merckx J. Duarte M.V.A. Pedroso L.G.A. Khaustov A.A. Myers-Hansen J.L. Wäkers F.L. Going asexual: A survey of mites of the genus Thyreophagus (Acari: Acaridae) revealing a large number of new parthenogenetic species in the Holarctic Region Life 202313216810.3390/life 1311216838004308 PMC 10672012 · doi ↗ · pubmed ↗
- 6Klimov P.B. Demard E.P. Stinson C.S.A. Duarte M.V.A. Wäckers F.L. Vangansbeke D. Thyreophagus calusorum sp. n. (Acari, Acaridae), a new parthenogenetic species from the USA, with a checklist of Thyreophagus species of the world Syst. Appl. Acarol.2022271920195610.11158/saa.27.10.7 · doi ↗
- 7Klimov P.B. Kolesnikov V.B. Khaustov V.A. Khaustov A.A. Thyreophagus tauricus sp. n., a New Subcortical Mite Species (Acari: Acaridae), with a COX 1 DNA Sequence Analysis of Several Economically Important Species of Thyreophagus Animals 202313354610.3390/ani 1322354638003163 PMC 10668731 · doi ↗ · pubmed ↗
- 8Laboulbène A. Description de l’Acams entomophagus Ann. Soc. Entomol. Fr. Bull.18522317338