A chromosomal reference genome sequence for the malaria mosquito, Anopheles maculipalpis, Giles, 1902
Nil Rahola, Diego Ayala, Lemonde B. A. Bouafou, Boris K. Makanga, Harriet F. Johnson, Haynes Heaton, Martin G. Wagah, Joanna C. Collins, Ksenia Krasheninnikova, Sarah E. Pelan, Damon-Lee B. Pointon, Ying Sims, James W. Torrance, Alan Tracey, Marcela Uliano-Silva

TL;DR
This paper provides a detailed chromosomal genome sequence for the malaria mosquito Anopheles maculipalpis, including a complete mitochondrial genome.
Contribution
The novel contribution is the first chromosomal-level genome assembly for Anopheles maculipalpis, including the X chromosome and mitochondrial genome.
Findings
The genome assembly spans 224 megabases and is scaffolded into three chromosomal pseudomolecules.
The complete mitochondrial genome is 15.4 kilobases in length.
Abstract
We present a genome assembly from an individual female Anopheles maculipalpis (the malaria mosquito; Arthropoda; Insecta; Diptera; Culicidae). The genome sequence is 224 megabases in span. Most of the assembly is scaffolded into three chromosomal pseudomolecules with the X sex chromosome assembled. The complete mitochondrial genome was also assembled and is 15.4 kilobases in length.
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
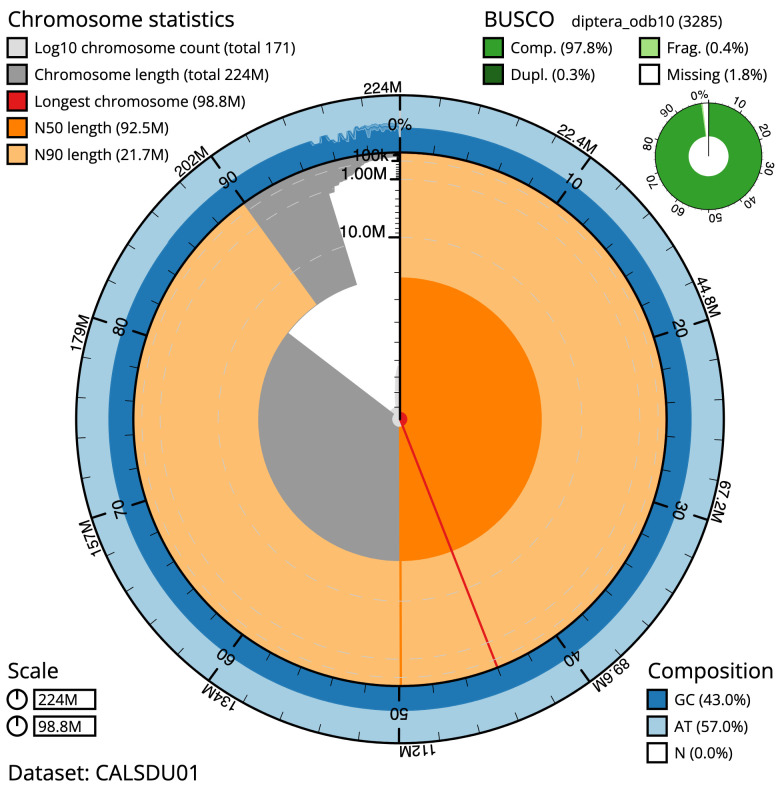 Figure 1
Figure 1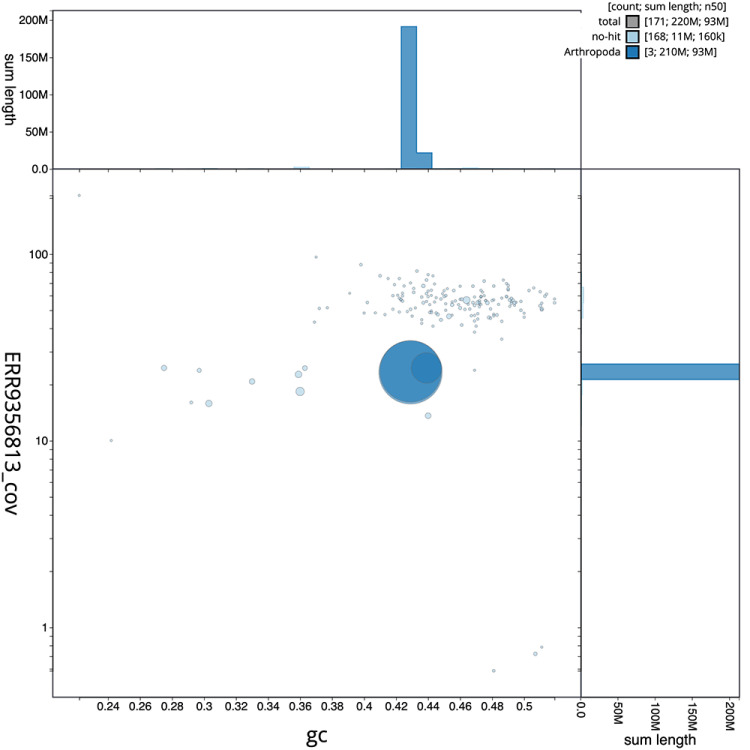 Figure 2
Figure 2 Figure 3
Figure 3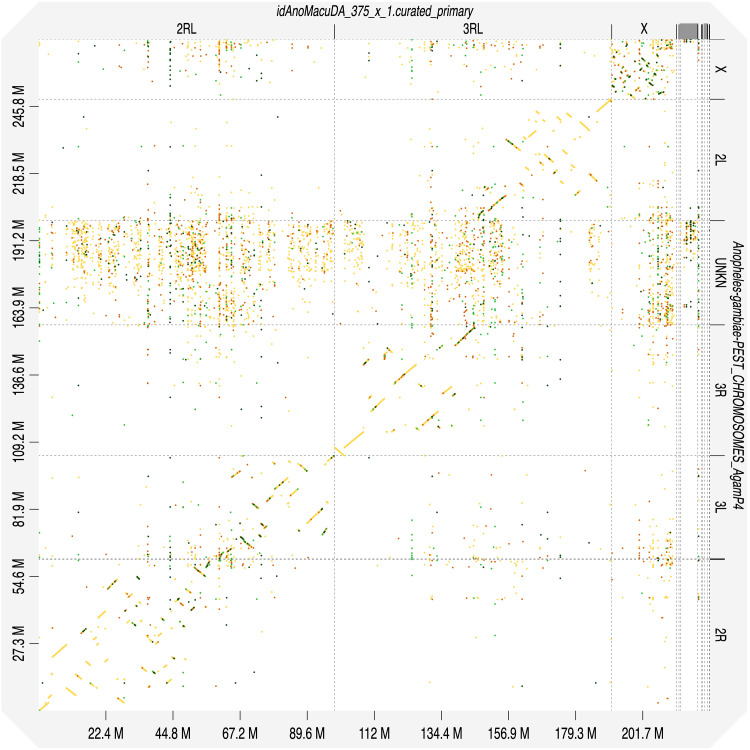 Figure 4
Figure 4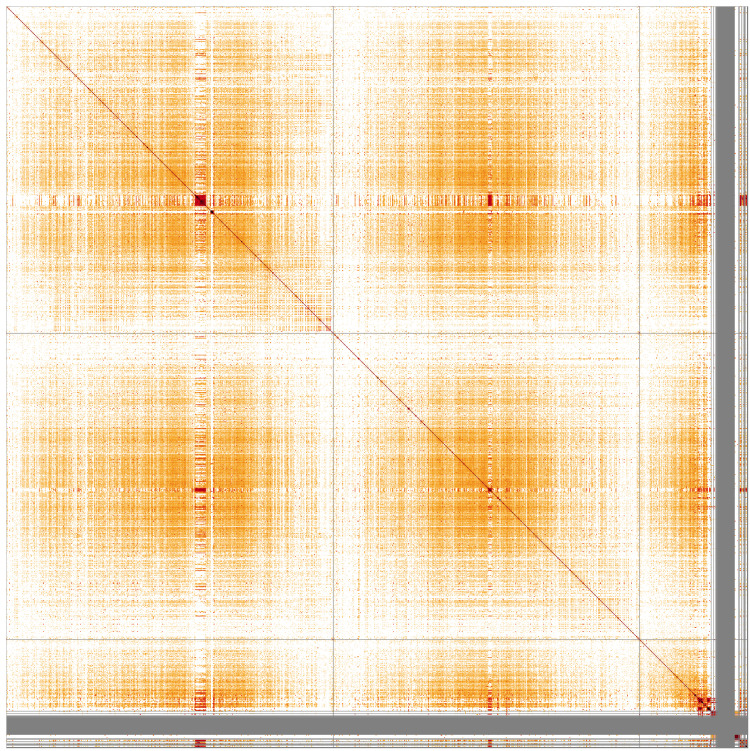 Figure 5
Figure 5|
| |
|---|---|
| Assembly identifier | idAnoMacuDA_375_x |
| Species |
|
| Specimen | idAnoMacuDA-375_x |
| NCBI taxonomy ID | 1496333 |
| BioProject | PRJEB53251 |
| BioSample ID | ERS10527369 |
| Isolate information | female, whole organism |
|
| |
| PacificBiosciences SEQUEL II | ERR9439503 |
| 10X Genomics Illumina | ERR9356813, ERR9356814,
|
| Hi-C Illumina | ERR9356811, ERR9356812 |
| PolyA RNA-Seq Illumina | ERR9356817 |
|
| |
| Assembly accession | GCA_943734695 |
| Accession of alternate
| GCA_943734695 |
| Span (Mb) | 224.075 |
| Number of contigs | 182 |
| Contig N50 length (Mb) | 22.618 |
| Number of scaffolds | 171 |
| Scaffold N50 length (Mb) | 92.549 |
| Longest scaffold (Mb) | 98.751 |
| BUSCO
| C:97.8%[S:97.5%,D:0.3%],
|
| INSDC
| Chromosome | Size (Mb) | Count | Gaps |
|---|---|---|---|---|
| 2RL | 98.751 | 1 | 3 | |
| 3RL | 92.549 | 1 | 3 | |
| X | 21.651 | 1 | 1 | |
| MT | 0.015 | 1 | 0 | |
| X Unlocalised | 7.073 | 155 | 3 | |
| Unplaced | 4.035 | 12 | 1 |
| Chromosome | Start | End | Chromosome
|
|---|---|---|---|
| 2RL | 1 | 57,070,844 | 2R |
| 2RL | 60,243,198 | 98,751,411 | 2L |
| 3RL | 1 | 46,889,622 | 3R |
| 3RL | 47,935,636 | 92,548,589 | 3L |
| X | 1 | 21,651,475 | X |
| Software tool | Version | Source |
|---|---|---|
| hifiasm | 0.14 |
|
| purge_dups | 1.2.3 |
|
| SALSA2 | 2.2-4c80ac1 |
|
| longranger align | 2.2.2 |
|
| freebayes | 1.3.1 |
|
| MitoHiFi | 2 |
|
| gEVAL | N/A |
|
| HiGlass | 1.11.6 |
|
| PretextView | 0.1.x |
|
| BlobToolKit | 3.4.0 |
|
| BUSCO | 5.3.2 |
|
| D-GENIES | 1.4 |
|
| StainedGlass | 0.5 |
|
| ULTRA | 1.0.0 beta |
|
- —Wellcome Trust
- —Wellcome Trust Grant
- —ANR
- —Bill & Melinda Gates Foundation Award
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsInsect symbiosis and bacterial influences · Invertebrate Immune Response Mechanisms · Insect Resistance and Genetics
Species taxonomy
Animalia; Arthropoda; Insecta; Diptera; Culicidae; Anophelinae; Anopheles; Anopheles maculipalpis; Giles, 1902 (NCBI txid:1496333).
Background
The mosquito Anopheles maculipalpis (Giles, 1902) has a very large distribution throughout Africa and is also present in the islands of South-West Indian Ocean ^ 1– 4 ^. It is a savannah species found up to 1000 metres above sea level in Central Africa ^ 1, 5 ^. Its larvae develop in poorly oxygenated environments, typically among abundant aquatic vegetation. Their habitats are sunny with shallow, stagnant water, often muddy. They vary greatly in type: rice fields, stagnant ditches and canals, holes in the ground, hoofprints, rock hollows, shallow or drying ponds. Some larvae have been collected in rivers, which is exceptional. An. maculipalpis is often found in the larval stage in association with An. gambiae, An. squamosus, and An. mascarensis ^ 4 ^. The pre-imaginal development period of this species is relatively long, at least one month. An. maculipalpis is generally considered zoophilic, exophagic, and exophilic ^ 4, 6 ^. In some localities, however, this species is reported to exhibit moderate anthropophily. This species is considered to have no medical importance, although a strain of the West Nile virus has been isolated in Madagascar, and experimental transmission of Wuchereria bancrofti has been achieved in the laboratory in Mauritius (Mascarene Archipelago, Indian Ocean) ^ 7 ^. Few studies have generated genetic sequences such as cytochrome oxidase subunit II (COII) or internal transcribed spacer 2 (ITS2) ^ 8, 9 ^. However, no population genetic work has been published, because of its no proven medical importance as a malaria vector.
The genome of the African malaria mosquito, Anopheles maculipalpis, was sequenced as part of the Anopheles Reference Genomes Project (PRJEB51690). Here we present a chromosomally complete genome sequence for Anopheles maculipalpis, based on a single wild-caught female.
Genome sequence report
The genome was sequenced from a single female Anopheles maculipalpis collected in Lopé, Gabon (-0.142, 11.610) in April 2019. A total of 52-fold coverage in Pacific Biosciences single-molecule HiFi long reads (N50 10.236 kb) and 79-fold coverage in 10X Genomics read clouds were generated. Primary assembly contigs were scaffolded with chromosome conformation Hi-C data from an unrelated female individual.
The final assembly has a total length of 224 Mb in 171 sequence scaffolds with a scaffold N50 of 92.549 Mb ( Table 1). The snail plot in Figure 1 provides a summary of the assembly statistics, while the distribution of assembly scaffolds on GC proportion and coverage is shown in Figure 2. 98.19% of the assembly sequence was assigned to three chromosomal-level scaffolds, representing two autosomes and the X sex chromosome ( Figure 3; Table 2). Chromosomes were numbered and oriented using synteny to the AgamP3 assembly ^ 10 ^ (accession GCF_000005575.2) ( Figure 4) based on cytogenetics data ^ 11 ^ ( Figure 4). The assembly has a BUSCO 5.3.2 ^ 12 ^ completeness of 97.8% using the diptera_odb10 reference set. While not fully phased, the assembly deposited is of one haplotype and also includes the circular mitochondrial genome. Contigs corresponding to the second haplotype have also been deposited.
Table 1.: Genome data for An. maculipalpis, idAnoMacuDA_375_x.
Snail plot summary of assembly statistics for Anopheles maculipalpis assembly.The main plot is divided into 1,000 size-ordered bins around the circumference with each bin representing 0.1% of the 224,074,805 bp assembly. The distribution of sequence lengths is shown in dark grey with the plot radius scaled to the longest sequence present in the assembly (98,751,411 bp, shown in red). Orange and pale-orange arcs show the N50 and N90 sequence lengths (92,548,589 and 21,651,475 bp), respectively. The pale grey spiral shows the cumulative sequence count on a log scale with white scale lines showing successive orders of magnitude. The blue and pale-blue area around the outside of the plot shows the distribution of GC, AT and N percentages in the same bins as the inner plot. A summary of complete, fragmented, duplicated and missing BUSCO genes in the diptera_odb10 set is shown in the top right. An interactive version of this figure is available at https://blobtoolkit.genomehubs.org/view/idAnoMacuDA_375_x/dataset/CALSDU01/snail.
Blob plot of base coverage in a subset of idAnoMacuDA-375_x 10x linked reads against GC proportion for An. maculipalpis assembly idAnoMacuDA_375_x.Chromosomes are coloured by phylum. Circles are sized in proportion to chromosome length. Histograms show the distribution of chromosome length sum along each axis. An interactive version of this figure is available at https://blobtoolkit.genomehubs.org/view/idAnoMacuDA_375_x/dataset/CALSDU01/blob.
Genome assembly of An. maculipalpis, idAnoMacuDA_375_x: Hi-C contact map.Visualised in HiGlass. Chromosomes are ordered as 2RL, 3RL, X, then remaining scaffolds. The interactive Hi-C map can be viewed at https://genome-note-higlass.tol.sanger.ac.uk/l/?d=LR0ErKCBRiyLbpoq-qAnnw.
Table 2.: Chromosomal pseudomolecules in the genome assembly of An. maculipalpis, idAnoMacuDA_375_x.
*Alignment dotplot between genome assemblies of An. maculipalpis idAnoMacuDA_375_x and An. gambiae, AgamP4 (PEST).Chromosome arms correspondence (maculipalpus-gambiae): 2R-2R, 2L-3L, 3R-3R, 3L-2L in agreement with
11 .*
Chromosome arms, candidate centromere and the rDNA regions were delineated based on the presence of characteristic tandem repeat arrays ( Figure 5; Table 3). Putative centromeres of autosomes comprised long stretches of irregular tandem repeats with varying unit lengths and arrangements, but with significant sequence homology between chromosomes 2 and 3. These locations were in agreement with the Hi-C contact map ( Figure 3) and synteny to An. gambiae ( Figure 4). Terminal part of chromosome X, starting at 17,790,992, also featured variable tandem repeats, which could be grouped into at least six different types based on sequence similarity. None of those could be reliably identified as centromeric. Arrays of rRNA genes were located in X-linked unlocalised scaffolds.
Sequence similarity heatmap for genome assembly of An. maculipalpis, idAnoMacuDA_375_x.Produced with StainedGlass, visualised in HiGlass. CChromosomes are ordered as 2RL, 3RL, X, then remaining scaffolds. Darker colours represent higher sequence similarity, notably at putative centromeres and terminal part of X.
Table 3.: Chromosome arms in the genome assembly of An. maculipalpis, idAnoMacuDA_375_x.
Gene annotation was performed with NCBI Eukaryotic Genome Annotation Pipeline and is available in the RefSeq ^ 13 ^ under the accession GCF_943734695.1. A total of 12,929 genes were predicted, including 11,533 protein-coding genes and 1,092 non-coding RNAs. The genome assembly and gene annotations are hosted on VectorBase, www.vectorbase.org ^ 14 ^ under the identifier AmacGA1.
Methods
Sample acquisition and nucleic acid extraction
Anopheles maculipalpis individuals were caught using a human landing catch. A single female idAnoMacuDA-375_x was used for Pacific BioSciences and 10x genomics, an unrelated female idAnoMacuDA-405_x was used for Arima Hi-C.
For high molecular weight (HMW) DNA extraction one whole insect (idAnoMacuDA-375_x) was disrupted by manual grinding with a blue plastic pestle in Qiagen MagAttract lysis buffer and then extracted using the Qiagen MagAttract HMW DNA extraction kit with two minor modifications ^ 15 ^. The quality of the DNA was evaluated using an Agilent FemtoPulse to ensure that most DNA molecules were larger than 30 kb, and preferably > 100 kb. In general, single mosquito extractions ranged in total estimated DNA yield from 200 ng to 800 ng, with an average yield of 500 ng. Low molecular weight DNA was removed using an 0.8X AMpure XP purification. A small aliquot (less than ~5% of the total volume) of HMW DNA was set aside for 10X Linked Read sequencing and the rest of the DNA was sheared to an average fragment size of 12–20 Kb using a Diagenode Megaruptor 3 at speeds ranging from 27 to 30.Sheared DNA was purified using AMPure PB beads with a 1.8X ratio of beads to sample to remove the shorter fragments and concentrate the DNA sample. The concentration and quality of the sheared and purified DNA was assessed using a Nanodrop spectrophotometer and Qubit Fluorometer with the Qubit dsDNA High Sensitivity Assay kit. Fragment size distribution was evaluated by running the sheared and cleaned sample on the FemtoPulse system once more. The median DNA fragment size for Anopheles mosquitoes was 15 kb and the median yield of sheared DNA was 200 ng, with samples typically losing about 50% of the original estimated DNA quantity through the process of shearing and purification.
For Hi-C data generation, a separate unrelated mosquito specimen (idAnoMacuDA-405_x) was used as input material for the Arima V2 Kit according to the manufacturer’s instructions for animal tissue. This approach of using a sibling was taken in order to enable all material from a single specimen to contribute to the PacBio data generation given we were not always able to meet the minimum suggested guidance of starting with > 300 ng of HMW DNA from a specimen. Samples proceeded to the Illumina library prep stage even if they were suboptimal (too little tissue) going into the Arima reaction.
To assist with annotation, which will be made available through VectorBase ^ 14 ^ in due course, RNA was extracted from a separate whole unrelated female mosquito specimen (idAnoMacuDA-401_x) using TRIzol, according to the manufacturer’s instructions. RNA was then eluted in 50 μl RNAse-free water, and its concentration was assessed using a Nanodrop spectrophotometer and Qubit Fluorometer using the Qubit RNA Broad-Range (BR) Assay kit. Analysis of the integrity of the RNA was done using Agilent RNA 6000 Pico Kit and Eukaryotic Total RNA assay. Samples were not always ideally preserved for RNA, so qualities varied but all were sequenced anyway.
Sequencing
We prepared libraries as per the PacBio procedure and checklist for SMRTbell Libraries using Express TPK 2.0 with low DNA input. Every library was barcoded to support multiplexing. Final library yields ranged from 20 ng to 100 ng, representing only about 25% of the input sheared DNA. Libraries from two specimens were typically multiplexed on a single 8M SMRT Cell. Sequencing complexes were made using Sequencing Primer v4 and DNA Polymerase v2.0. Sequencing was carried out on the Sequel II system with 24-hour run time and 2-hour pre-extension. A 10X Genomics Chromium read cloud sequencing library was also constructed according to the manufacturer’s instructions (this product is no longer available). Only 0.5 ng of DNA was used and only 25–50% of the gel emulsion was put forward for library prep due to the small genome size. For Hi-C data generation, following the Arima HiC V2 reaction, samples were processed through Library Preparation using a NEB Next Ultra II DNA Library Prep Kit and sequenced aiming for 100x depth. RNA libraries were created using the directional NEB Ultra II stranded kit. Sequencing was performed by the Scientific Operations core at the Wellcome Sanger Institute on Pacific Biosciences SEQUEL II (HiFi), Illumina NovaSeq 6000 (10X and Hi-C), or Illumina HiSeq 4000 (RNAseq).
Genome assembly
Assembly was carried out with Hifiasm ^ 16 ^; haplotypic duplications were identified and removed with purge_dups ^ 17 ^. One round of polishing was performed by aligning 10X Genomics read data to the assembly with Long Ranger ALIGN, calling variants with FreeBayes ^ 18 ^. The assembly was then scaffolded with Hi-C data ^ 19 ^ using SALSA2 ^ 20 ^. The assembly was checked for contamination as described previously ^ 21 ^. Manual curation was performed using gEVAL ^ 22 ^, HiGlass ^ 23 ^ and Pretext ^ 24 ^. The mitochondrial genome was assembled using MitoHiFi ^ 25 ^, which performs annotation using MitoFinder ^ 26 ^. The genome was analysed and BUSCO scores were generated within the BlobToolKit environment ^ 27 ^. Synteny analysis was performed with D-GENIES ^ 28 ^. Repetitive sequences were visualised with StainedGlass ^ 29 ^ and tandem repeats were annotated with ULTRA ^ 30 ^. Table 4 contains a list of all software tool versions used, where appropriate.
Ethics/compliance issues
The genetic resources accessed and utilised under this project were done so in accordance with the UK ABS legislation (Nagoya Protocol (Compliance) (Amendment) (EU Exit) Regulations 2018 (SI 2018/1393)) and the national ABS legislation within the country of origin, where applicable.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Kyalo D Amratia P Mundia CW : A geo-coded inventory of anophelines in the Afrotropical Region south of the Sahara: 1898–2016 [version 1; peer review: 3 approved]. Wellcome Open Res. 2017;2:57. 10.12688/wellcomeopenres.12187.1 28884158 PMC 5558104 · doi ↗ · pubmed ↗
- 2Tantely ML Le Goff G Boyer S : An updated checklist of mosquito species (Diptera: Culicidae) from Madagascar. Parasite. 2016;23: 20. 10.1051/parasite/2016018 27101839 PMC 4840257 · doi ↗ · pubmed ↗
- 3Gillies MT De Meillon B : The anophelinae of Africa south of the Sahara (Ethiopian zoogeographical region). South African Institute for Medical Research;1968. Reference Source
- 4Hervy JP Le Goff G Geoffroy B : Les anophèles de la région afro-tropicale: logiciel d’identification et d'enseignement.1998. Reference Source
- 5Irish SR Kyalo D Snow RW : Updated list of Anopheles species (Diptera: Culicidae) by country in the Afrotropical Region and associated Islands. Zootaxa. 2020;4747(3): zootaxa.4747.3.1. 10.11646/zootaxa.4747.3.1 32230095 PMC 7116328 · doi ↗ · pubmed ↗
- 6Saili K de Jager C Sangoro OP : Anopheles rufipes implicated in malaria transmission both indoors and outdoors alongside Anopheles funestus and Anopheles arabiensis in rural south-east Zambia. Malar J. 2023;22(1): 95. 10.1186/s 12936-023-04489-3 36927373 PMC 10018844 · doi ↗ · pubmed ↗
- 7Gebert S : Notes on filariasis and its transmission by Mauritian anophelines. Trans R Soc Trop Med Hyg. 1937;30(4):477–480. 10.1016/S 0035-9203(37)90136-6 · doi ↗
- 8Ayala D Akone-Ella O Rahola N : Natural Wolbachia infections are common in the major malaria vectors in Central Africa. Evol Appl. 2019;12(8):1583–1594. 10.1111/eva.12804 31462916 PMC 6708434 · doi ↗ · pubmed ↗