Molecular evidence supports the functionality of a protein-trapped endogenous allele of Dally-like protein
Drew Delmore, Indrayani Waghmare

TL;DR
This study confirms that a modified version of the Dally-like protein in fruit flies is functional and essential for development.
Contribution
The study provides molecular evidence that the SVS-tagged Dlp protein is functional without producing a wild-type isoform.
Findings
SVS-dlp flies do not produce a wild-type Dlp isoform.
The SVS-tagged Dlp is functional and supports embryonic development.
Alternative splicing does not rescue the wild-type Dlp in SVS-dlp flies.
Abstract
The Drosophila glypican Dally-like protein (Dlp) is an evolutionarily-conserved cell-surface protein that modulates extracellular distribution of several secreted ligands for cell signaling. Several fly lines expressing tagged dlp have been used to study the role of Dlp in vivo including the PBac{602.P.SVS-1}dlp [CPTI000445] protein-trap line, which encodes StrepII-Venus-StrepII (SVS)-tagged Dlp from the endogenous locus. dlp is essential for embryonic development, and the SVS-dlp line is homozygous viable. Although this suggests that the SVS-tagged Dlp is functional, it is possible that that the SVS-dlp flies produce wild-type dlp isoform through alternative splicing, contributing to their survival. Here, we used a molecular analysis approach to show that the SVS-dlp flies do not produce wild-type isoform, confirming that the SVS-tagged Dlp is indeed functional.
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
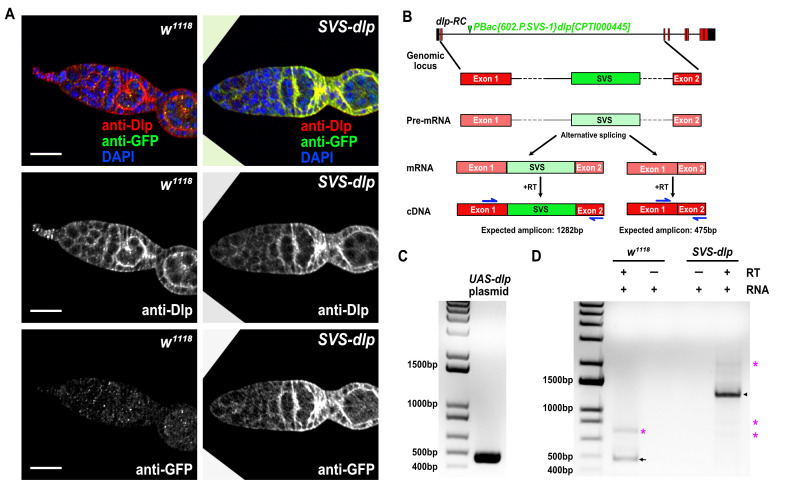 Figure 1
Figure 1|
|
|
|
|
|
Control |
|
Bloomington stock center |
BDSC_3605 |
|
|
|
Kyoto stock center |
115031 |
|
|
|
| |
|
Forward primer |
ATGCTACATCAGCAGCAACA |
Generated in this study | |
|
Reverse primer |
CTTTGAATTTTGTGGCCTTG |
Generated in this study | |
- —National Institutes of Health (United States)https://ror.org/01cwqze88
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsPlant Molecular Biology Research · Plant Taxonomy and Phylogenetics · Plant Reproductive Biology
Description
Cells in multicellular organisms communicate with each other to ensure proper tissue development and homeostasis. The intercellular communication is typically orchestrated by several secreted ligands such as Wnts, Hedgehog, and Decapentaplegic that are produced by source cells to activate signaling in target cells, which may be located at short or long range (Lawrence and Struhl, 1996; Neumann and Cohen, 1997) . The extracellular distribution of the secreted ligands from the source cells partly depends on a cell-surface glypican called Dally-like protein (Dlp), which performs an evolutionarily-conserved function of interacting with extracellular ligands ensuring proper ligand availability at both short range and long range distances (Baeg et al., 2001; Filmus et al., 2008; Yan and Lin, 2009; Waghmare and Page-McCaw, 2022). Loss of dlp in a tissue impairs signaling at both short and long range (Desbordes and Sanson, 2003; Baeg et al., 2004; Kreuger et al., 2004; Franch-Marro et al., 2005; Yan et al., 2009; Zhang et al., 2013; Chaudhary et al., 2019).
Several transgenic flies overexpressing GFP-tagged dlp have been previously generated to elucidate Dlp’s function in vivo (Baeg et al., 2001; Han et al., 2004; Kreuger et al., 2004; Marois et al., 2006; Gallet et al., 2008). However, only a few of tagged constructs have been tested for functionality and only in a tissue-specific context, and at least one GFP-dlp transgene expressed in flies has been previously reported to be non-functional (Baeg et al., 2001; Han et al., 2004; Waghmare et al., 2020) . The use of non-functional tagged protein constructs can obscure our understanding of its function. For example, previous studies that have used different tagged-Dlp constructs have arrived at contradictory findings regarding its role in transcytosis of Wingless ligand from apical to basal surfaces in the epithelial cells (Gallet et al., 2008; Yan et al., 2009) . Such contradictions in the published literature underscore the importance of assessing whether or not tagged proteins expressed *in vivo * are functional.
In 2014, a large-scale protein trap screen using a hybrid piggyBac vector isolated a fly line that produced StrepII-Venus-StrepII (SVS)-tagged Dlp from the endogenous locus (Lowe et al., 2014) . The StrepII tag can be used for affinity purification of the tagged protein, and Venus is a fluorophore derived from GFP that allows protein visualization using fluorescence microscopy ( Nagai et al., 2002; Maertens et al. 2015) . The
- piggyBac* insertion in the first intron contains an artificial exon that encodes SVS flanked by strong splice acceptor and donor sequences from the Myosin heavy chain ( Mhc ) gene. The SVS-dlp protein trap line is healthy and homozygous viable, and SVS-tagged Dlp can be detected in tissues isolated from SVS-dlp flies ( Fig. 1A ) (Lowe et al., 2014; Norman et al., 2016; Su et al., 2018; Waghmare et al., 2020) . Because *dlp * loss-of-function is embryonic lethal (Baeg et al., 2001; Han et al., 2004) , the homozygous protein trap line must produce a functional protein. While this is a reasonable assumption, it comes with a caveat. It is possible that the protein-trap produces wild-type *dlp * isoform through alternative splicing ( Fig. 1B ), contributing to the survival of the homozygous flies. This potential issue may be circumvented by using CRISPR-Cas9 to delete the endogenous locus followed by ФC31 integrase-mediated knock-in of tagged protein sequence. Indeed, a recent study has generated a fly line that expresses YFP-tagged *dlp * from endogenous locus using this strategy (Ell et al. 2024) . This line, however, is not homozygous viable, and therefore likely non-functional. These data further suggest that the position of the tag within a protein likely affects its function and underscore the importance of determining if the existing endogenously-tagged SVS-dlp line is functional. Standard genetic complementation assays cannot test this possibility. Therefore, we used Reverse Transcriptase- Polymerase Chain Reaction (RT-PCR) to test if the SVS-dlp flies produce the wild-type isoform. RT- PCR generates amplicons using cDNA as a template generated from the mRNA isoforms present in the sample using reverse transcriptase. Thus, the cDNA acts as a proxy for the mRNA isoforms present in the sample. We designed forward and reverse primers against sequences in exons 1 and 2 respectively that generate the expected 475bp amplicon when *UAS-dlp * plasmid is used as template ( Fig. 1C ). These primers are also able to amplify the 475bp amplicon from wild-type dlp isoform and the 1282bp amplicon from the *SVS-dlp * isoform present in the samples isolated from fly tissues ( Fig. 1D ). Our results show that the primers can detect the wild-type *dlp * isoform in
w ^1118^ * flies but this isoform is not detected in SVS-Dlp flies. Further, the same primers detect the *SVS-dlp * isoform in SVS-Dlp flies. Because SVS-Dlp flies do not produce detectable wild-type *dlp * isoform to contribute to their survival, we conclude that the *SVS-dlp * isoform is functional.
Methods
Total RNA was isolated from adult * w ^1118^ * (control) and *SVS-dlp * ( * PBac{602.P.SVS-1}dlp ^[CPTI000445]^ * ) protein-trap line using the RNeasy Mini Kit (Qiagen) per manufacturer’s protocol. 6-8 adult male and female flies were used. The RNA sample was treated with DNAse to remove any possible genomic DNA contamination during RNA isolation. SuperScript IV VILO kit (Invitrogen) was used to synthesize cDNA from RNA. PCR was performed using Q5 polymerase (NEB) using the manufacturer’s protocol. DNA gel electrophoresis was performed on PCR products, which were separated using ethidium bromide-containing 1% agarose gel. The UAS-dlp plasmid used in this study contains wild-type *dlp * sequence cloned into pUAST vector and was obtained from Xinhua Lin.
Immunohistochemistry was performed on age-matched seven-day old female flies using previously published protocol (Waghmare et al., 2020) . Briefly, ovaries were dissected in ice-cold PBS (Phosphate buffered saline), fixed in 4% paraformaldehyde for 20 minutes as room temperature, washed with 1X PBST (PBS plus 0.1% Triton X-100), and incubated for 1h in 5% NGS (normal goat serum in 1X PBS). After blocking, tissues were incubated overnight with mouse IgG1 anti-Dlp (13G8; 1:5) from Developmental Studies Hybridoma Bank (DSHB) and rabbit anti-GFP (Abcam ab6556; 1:200). Following 3 washes with 1xPBST, samples were were incubated with Cy3-conjugated goat anti-mouse IgG1 (115–165-205) and FITC-conjugated goat anti-rabbit (# 711-095-152) secondary antibodies for 1h in dark at room temperature (1:500, Jackson Immuno Research). DAPI was used to stain nuclei (1 μg/ml), and the samples were mounted in Vectashield (Vector Laboratories). Images were captured using Zeiss Apotome Imager M2 under identical settings, and figures were generated using Affinity designer and Affinity Photo.
Reagents
**: **
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Baeg GH Lin X Khare N Baumgartner S Perrimon N 200111 Heparan sulfate proteoglycans are critical for the organization of the extracellular distribution of Wingless.Development 12810950-1991879410.1242/dev.128.1.8711092814 · doi ↗ · pubmed ↗
- 2Baeg GH Selva EM Goodman RM Dasgupta R Perrimon N 2004121 The Wingless morphogen gradient is established by the cooperative action of Frizzled and Heparan Sulfate Proteoglycan receptors.Dev Biol 27610012-16068910010.1016/j.ydbio.2004.08.02315531366 · doi ↗ · pubmed ↗
- 3Chaudhary V Hingole S Frei J Port F Strutt D Boutros M 201989 Robust Wnt signaling is maintained by a Wg protein gradient and Fz 2 receptor activity in the developing Drosophila wing.Development 146150950-199110.1242/dev.17478931399474 PMC 6703709 · doi ↗ · pubmed ↗
- 4Desbordes SC Sanson B 2003115 The glypican Dally-like is required for Hedgehog signalling in the embryonic epidermis of Drosophila.Development 130250950-19916245625510.1242/dev.0087414602684 · doi ↗ · pubmed ↗
- 5Ell Clara-Maria Safyan Abu Chayengia Mrinal Kustermann Manuela M. M. Lorenz Jennifer Schächtle Melanie Pyrowolakis George 202474 A genome engineered tool set for Drosophila TGF-β/BMP signaling studies 10.1101/2024.07.02.601693 PMC 1160769339494616 · doi ↗ · pubmed ↗
- 6Filmus J Capurro M Rast J 2008522 Glypicans.Genome Biol 951465-690622422410.1186/gb-2008-9-5-22418505598 PMC 2441458 · doi ↗ · pubmed ↗
- 7Franch-Marro X Marchand O Piddini E Ricardo S Alexandre C Vincent JP 2005112 Glypicans shunt the Wingless signal between local signalling and further transport.Development 13240950-199165966610.1242/dev.0163915647318 · doi ↗ · pubmed ↗
- 8Gallet A Staccini-Lavenant L Thérond PP 200851 Cellular trafficking of the glypican Dally-like is required for full-strength Hedgehog signaling and wingless transcytosis.Dev Cell 1451534-580771272510.1016/j.devcel.2008.03.00118477454 · doi ↗ · pubmed ↗