Morpho-phylogenetic evidence reveals Pseudolomaanthathailandica gen. et sp. nov. and Submultiguttulisporamultiseptata gen. et sp. nov. in Chaetosphaeriaceae

Abstract
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
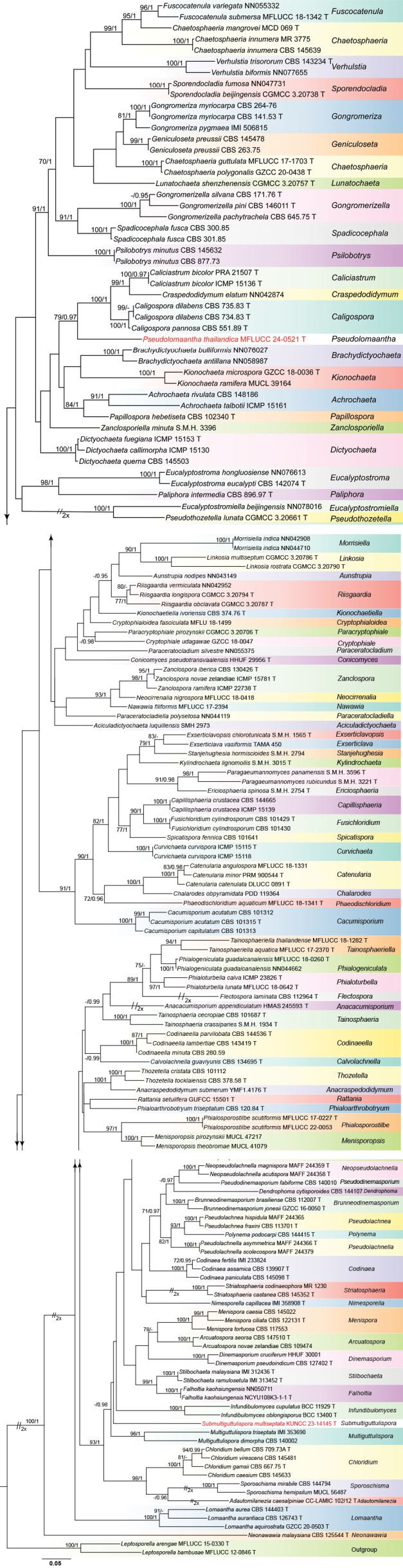 Figure 1
Figure 1 Figure 2
Figure 2 Figure 3
Figure 3| Taxon | Strain | Status |
|
|
|---|---|---|---|---|
|
| CBS 148186 |
|
| |
|
| ICMP 15161 |
|
| |
|
| SMH 2973 |
|
| |
|
| CC-LAMIC 102/12 | T |
|
|
|
| HMAS 245593 | T |
|
|
|
| YMF1.4176 | T |
|
|
|
| CBS 109474 |
|
| |
|
| CBS 147510 | T |
|
|
|
| NN043149 |
|
| |
|
| NN058987 |
|
| |
|
| NN076027 |
|
| |
|
| CBS 112007 | T |
|
|
|
| GZCC 16–0050 | T |
|
|
|
| CBS 101312 |
|
| |
|
| CBS 101315 | T |
|
|
|
| CBS 101313 |
|
| |
|
| ICMP 15136 | T |
|
|
|
| PRA-21507 | T |
|
|
|
| CBS 734.83 | T |
|
|
|
| CBS 735.83 | T |
|
|
|
| CBS 551.89 | T |
|
|
|
| CBS 134695 | T |
|
|
|
| CBS 144665 |
|
| |
|
| ICMP 15139 |
|
| |
|
|
|
| ||
|
| DLUCC 0891 | T |
|
|
|
| PRM 900544 | T |
|
|
|
| T |
|
| |
|
| M.R. 3775 |
|
| |
|
| CBS 145639 |
|
| |
|
| MCD 069 | T |
|
|
|
| GZCC 20–0438 | T |
|
|
|
| PDD 119364 |
|
| |
|
| CBS 709.73A | T |
|
|
|
| CBS 145633 |
|
| |
|
| CBS 667.75 | T |
|
|
|
| CBS 145481 |
|
| |
|
| CBS 139907 | T |
|
|
|
| IMI 233824 |
|
| |
|
| CBS 145098 | T |
|
|
|
| CBS 143419 | T |
|
|
|
| CBS 280.59 |
|
| |
|
| CBS 144536 | T |
|
|
|
| HHUF 29956 | T |
|
|
|
| NN042874 |
|
| |
|
| GZCC 18–0047 |
|
| |
|
|
|
| ||
|
| ICMP 15115 | T |
|
|
|
| ICMP 15118 |
|
| |
|
| CBS 144107 |
|
| |
|
| ICMP 15130 |
|
| |
|
| ICMP 15153 | T |
|
|
|
| CBS 145503 |
|
| |
|
| HHUF 30001 |
|
| |
|
| CBS 127402 | T |
|
|
|
| S.M.H. 2754 | T |
|
|
|
| CBS 142074 | T |
|
|
|
| NN076613 |
|
| |
|
| NN078016 |
|
| |
|
| TAMA 450 |
|
| |
|
| S.M.H. 1565 | T |
|
|
|
| NCYU108K3-1-1 | T |
|
|
|
| NN050711 |
|
| |
|
| CBS 112964 | T |
|
|
|
| T |
|
| |
|
| NN055332 |
|
| |
|
| CBS 101429 | T |
|
|
|
| CBS 101430 |
|
| |
|
| CBS 263.75 |
|
| |
|
| CBS 145478 |
|
| |
|
| CBS 264.76 |
|
| |
|
| CBS 141.53 | T |
|
|
|
| IMI 506815 |
|
| |
|
| CBS 645.75 | T |
|
|
|
| CBS 146011 | T |
|
|
|
| CBS 171.76 | T |
|
|
|
| BCC 11929 | T |
|
|
|
| BCC 13400 | T |
|
|
|
| GZCC 18–0036 | T |
|
|
|
| MUCL 39164 |
|
| |
|
| CBS 374.76 | T |
|
|
|
| S.M.H. 3015 | T |
|
|
|
| T |
|
| |
|
| T |
|
| |
|
| CGMCC 3.20786 | T |
|
|
|
| CGMCC 3.20790 | T |
|
|
|
| GZCC 20–0503 | T |
|
|
|
| CBS 126743 | T |
|
|
|
| CBS 144403 | T |
|
|
|
| CGMCC 3.20757 | T |
|
|
|
| CBS 145022 |
|
| |
|
| CBS 122131 | T |
|
|
|
| CBS 117553 |
|
| |
|
| MUCL 47217 |
|
| |
|
| MUCL 41079 |
|
| |
|
| NN042908 |
|
| |
|
| NN044710 |
|
| |
|
| CBS 140002 |
|
| |
|
| IMI 353690 |
|
| |
|
|
|
| ||
|
| CPC 16757 | T |
|
|
|
| MAFF 244358 | T |
|
|
|
| MAFF 244359 | T |
|
|
|
|
|
| ||
|
| IMI 358908 | T |
|
|
|
| CBS 896.97 | I |
|
|
|
| CBS 102340 | T |
|
|
|
| NN044119 |
|
| |
|
| NN055375 |
|
| |
|
| CGMCC 3.20706 | T |
|
|
|
| S.M.H. 3596 | T |
|
|
|
| S.M.H. 3221 | T |
|
|
|
| T |
|
| |
|
| CBS 120.84 | T |
|
|
|
| T |
|
| |
|
| NN044662 |
|
| |
|
| T |
|
| |
|
|
|
| ||
|
| ICMP 23826 | T |
|
|
|
| T |
|
| |
|
| CBS 144415 | T |
|
|
|
| CBS 140010 |
|
| |
|
| CBS 113701 | T |
|
|
|
| MAFF 244365 |
|
| |
|
| MAFF 244366 | T |
|
|
|
| MAFF 244379 |
|
| |
|
|
|
|
|
|
|
| CGMCC 3.20661 | T |
|
|
|
| CBS 877.73 |
|
| |
|
| CBS 145632 |
|
| |
|
| GUFCC 15501 | T |
|
|
|
| CGMCC 3.20794 | T |
|
|
|
| CGMCC 3.20787 | T |
|
|
|
| NN042952 |
|
| |
|
| CBS 301.85 |
|
| |
|
| CBS 300.85 |
|
| |
|
| CBS 101641 |
|
| |
|
| CGMCC 3.20738 | T |
|
|
|
| NN047731 |
|
| |
|
| MUCL 56487 |
|
| |
|
| CBS 144794 |
|
| |
|
| S.M.H.2794 |
|
| |
|
| IMI 312436 | T |
|
|
|
| IMI 313452 | T |
|
|
|
| CBS 145352 | T |
|
|
|
| M.R. 1230 |
|
| |
|
|
|
|
|
|
|
| CBS 101687 | T |
|
|
|
| S.M.H. 1934 | T |
|
|
|
| T |
|
| |
|
| T |
|
| |
|
| CBS 101112 |
|
| |
|
| CBS 378.58 | T |
|
|
|
| NN077655 |
|
| |
|
| CBS 143234 | T |
|
|
|
| ICMP 15781 | T |
|
|
|
| ICMP 22738 | T |
|
|
|
| CBS 130426 | T |
|
|
|
| S.M.H. 3396 |
|
|
| 1 |
| |
| – | Not sporidesmium-like genera |
|
| 2 | Conidiophores absent (reduced to conidiogenous cells), or solitary or in a small group |
|
| – | Conidiophores in synnemata |
|
| 3 | Conidiogenous cells polytretic; conidia hyaline, subfusiform to subacerose |
|
| – | Conidiogenous cells monoblastic; conidia black, helicoid |
|
| 4 | Conidia with appendage at the apex |
|
| – | Conidia without appendage |
|
| 5 | Conidia euseptate |
|
| – | Conidia distoseptate |
|
| 6 | Conidiophores absent or well-developed, conidia cylindrical, obclavate or narrowly fusiform, often with filamentous, hyaline apical appendages and typically bearing distinct pores in the distosepta |
|
| – | Conidiophores well-developed, conidia pyriform to obclavate, with a gold and glistening appendage around the apex, and distinct pores are not observed |
|
| 7 | Synanamorph of |
|
| – | Not synanamorph of |
|
| 8 | Conidiophores absent; conidia euseptate |
|
| – | Conidiophores absent, conidia distoseptate |
|
| 9 | Conidia obclavate, obclavate-rostrate, subcylindrical |
|
| – | Conidia cylindrical, clavate, or obclavate |
|
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsPlant Pathogens and Fungal Diseases · Mycorrhizal Fungi and Plant Interactions · Plant Diversity and Evolution
Introduction
Chaetosphaeriales was established by Huhndorf et al. (2004) to accommodate the family Chaetosphaeriaceae based on morphological characteristics and phylogenetic analysis of LSU sequence data. Currently, four families, viz., Chaetosphaeriaceae, Helminthosphaeriaceae, Leptosporellaceae, and Linocarpaceae are recognized within this order (Hyde et al. 2020; Wijayawardene et al. 2022). The estimated stem age of Chaetosphaeriales is approximately 158 MYA, based on divergence time analysis (Hyde et al. 2020).
Chaetosphaeriaceae was invalidly introduced by Locquin (1984) without a formal description but was later validly re-established by Réblová et al. (1999) to accommodate Chaetosphaeria and its relatives. Since its re-establishment, the family has shown remarkable diversity with a significant increase in the number of genera and species described in recent years (Lin et al. 2019; Yang et al. 2019; Réblová et al. 2020, 2021a, b, c, d, e, 2022, 2024; Wijayawardene et al. 2022; Wu and Diao 2022, 2023; Réblová and Nekvindová 2023; Yang et al. 2023; Hyde et al. 2024a). Wu and Diao (2022) conducted a comprehensive study of the anamorphic Chaetosphaeriaceae, which included 89 genera, establishing the family as one of the largest within Sordariomycetes (Wijayawardene et al. 2022). Their study also provided identification keys for most genera (Wu and Diao 2022). Following this, Réblová et al. (2022) and Réblová and Nekvindová (2023) carried out systematic reviews of chloridium-like morphotypes, resulting in the addition of seven new genera to Chaetosphaeriaceae. In subsequent years, five more new genera, viz., Gongromerizella, Neocirrenalia, Paragongromeriza, Pseudophialocephala and Pseudostriatosphaeria, were introduced based on morphology and phylogeny (Manawasinghe et al. 2022; Yang et al. 2023; Wu and Diao 2023; Tian et al. 2024; Zhang et al. 2024). Réblová et al. (2024) re-evaluated species in genera Exserticlava, Phaeostalagmus, Phialocephala, and several chalara- and stanjehughesia-like fungi, which led to the establishment of three new genera in Chaetosphaeriaceae. On the other hand, Ellisembia was removed from Chaetosphaeriaceae and reclassified under Sporidesmiaceae (Delgado et al. 2024; Hyde et al. 2024a). Hyde et al. (2024a) accepted 107 genera in Chaetosphaeriaceae.
The sexual morph of Chaetosphaeriaceae is characterized by perithecial, papillate, globose to subglobose, setose, dark brown to black ascomata; unitunicate, clavate to cylindrical asci with a J-, apical ring; and 0–3-septate, fusiform, cylindrical to ellipsoid, hyaline to brown ascospores, often with guttules, a sheath, or appendages (Réblová et al. 1999; Réblová and Gams 2000; Hyde et al. 2020; Wu and Diao 2022). The asexual morphs of Chaetosphaeriaceae include both hyphomycetes and coelomycetes. Coelomycetous morphs are characterized by stromatic, cupuliform or globose, unilocular, setose conidiomata; numerous, septate, ovoid to cylindrical setae; 4–6-septate, unbranched, pigmented conidiophores; integrated, holoblastic or enteroblastic, phialidic conidiogenous cells with conspicuous periclinal thickening at an attenuated apex; and aseptate, hyaline to brown conidia with tubular appendages at the ends (Hashimoto et al. 2015; Hyde et al. 2020; Li et al. 2020). Hyphomycetous morphs are further divided into phialidic and non-phialidic anamorphs. Phialidic anamorphs exhibit macronematous, mononematous, septate, pigmented conidiophores; mono- or polyphialidic conidiogenous cells that proliferate percurrently or sympodially, often with funnel-shaped collarettes; and aggregated, fusiform, allantoid, cylindrical or doliiform conidia, which are mostly hyaline but sometimes pigmented, and often possess filiform appendages (Réblová 2004; Fernández and Huhndorf 2005; Liu et al. 2016; Lin et al. 2019; Luo et al. 2019; Yang et al. 2019; Réblová et al. 2020, 2021a, b, d). Non-phialidic fungi in Chaetosphaeriaceae are predominantly characterized by sporidesmium-like asexual morphs (Ellis 1971, 1976; Wu and Zhuang 2005; Wu and Diao 2022; Yang et al. 2023; Delgado et al. 2024). These taxa are primarily saprobic, occurring on various plant substrates in both terrestrial and aquatic habitats, with some species also found in soil or as fungicolous taxa (Hughes and Kendrick 1968; Perera et al. 2016; Hyde et al. 2018; Réblová et al. 2020, 2021d; Wu and Diao 2022; Zhang et al. 2022; Calabon et al. 2023; Yang et al. 2024; Zhang et al. 2024).
In this study, we aim to introduce two new genera, Pseudolomaantha and Submultiguttulispora, to accommodate two new species, P.thailandica and S.multiseptata, respectively. Evidence from morphology and phylogenetic analysis of a combined LSU and ITS sequence dataset supports the establishment of these two new genera (Pseudolomaantha and Submultiguttulispora) within Chaetosphaeriaceae, Chaetosphaeriales, Sordariomycetes.
Material and methods
Collections, isolation and conservation
Samples of dead bamboo stems and wood were collected from Thailand and China. The collection information of the samples was noted (Rathnayaka et al. 2024), and the samples were taken to the laboratory in zip-lock plastic bags and subsequently examined using the methods described in Senanayake et al. (2020). Morphological observations of the fungal colonies on natural substrates were conducted using a stereomicroscope (Leica EZ4 Microsystems (Schweiz) AG, Singapore). A detailed examination of fungal structures was carried out using a Nikon ECLIPSE Ni compound microscope (Nikon, Japan) and photographed with a Nikon DS-Ri2 digital camera attached to the microscope. Measurements of fungal structures were made using Tarosoft^®^ Image Frame Work, and images used in figures were processed and assembled with Adobe Illustrator CS6 (Adobe Systems, San Jose, CA, USA).
Single-spore isolations were performed on water agar (WA), and germinated spores were transferred to potato dextrose agar (PDA) to obtain pure cultures (Chomnunti et al. 2014). Dried specimens were deposited in the Herbarium of Mae Fah Luang University (MFLU), Chiang Rai, Thailand, and the Herbarium of Cryptogams, Kunming Institute of Botany, Academia Sinica (HKAS), Kunming, China, and the Herbarium of Guizhou Academy of Agricultural Sciences (GZAAS), Guiyang, China. Pure cultures were deposited in the Mae Fah Luang University Culture Collection (MFLUCC) and the Kunming Institute of Botany Culture Collection (KUNCC). Faces of Fungi and Index Fungorum numbers were registered following the guidelines of Jayasiri et al. (2015) and Index Fungorum (http://www.indexfungorum.org/Names/Names.asp; accessed on 15 November 2024)
DNA extraction, PCR amplification and sequencing
Pure cultures were incubated at 25 °C–28 °C for one month. Fresh fungal mycelia were scraped from the surface of the colonies and transferred to a 1.5 mL microcentrifuge tube using a sterilized scalpel for genomic DNA extraction. Genomic DNA was extracted using the Biospin Fungus Genomic DNA Extraction Kit (Biospin Fungus Genomic DNA Extraction Kit, BioFlux®, Shanghai, China) following the manufacturer’s instructions. The large subunit of ribosomal DNA (LSU) and the internal transcribed spacer (ITS) gene regions were amplified using primers LR0R and LR5 (Vilgalys and Hester 1990) and ITS5 and ITS4 (White et al. 1990), respectively. Polymerase chain reaction (PCR) was performed in a 50 µL reaction mixture containing 2 µL of DNA template, 2 µL of each forward and reverse primer (10 µM), 25 µL of 2 × Taq PCR Master Mix with blue dye (Sangon Biotech, China), and 19 µL of distilled–deionized water. Amplification conditions for the LSU and ITS regions followed the protocol described by Zhang et al. (2022). The quality of PCR products was assessed using 1% agarose gel electrophoresis stained with ethidium bromide. Purification and sequencing of PCR products were performed by Beijing Qingke Biotechnology Co., Ltd.
Phylogenetic analyses
Original sequences were verified using BioEdit v. 7.1.3.0 (Hall 1999), and were assembled using SeqMan v. 7.0.0 (DNASTAR, Madison, WI, USA). The newly generated sequences were subjected to BLAST searches in GenBank to determine closely related taxa. Taxa used in the phylogenetic analysis for Chaetosphaeriaceae were selected and obtained from previous studies and GenBank (Wu and Diao 2022; Zhang et al. 2022; Réblová and Nekvindová 2023; Réblová et al. 2024). Sequence alignments for each locus were aligned using the online multiple alignment program MAFFT v.7 (http://mafft.cbrc.jp/alignment/server/, accessed September 2024; Katoh et al. 2019). The alignments were visually checked and manually improved where necessary using BioEdit v. 7.1.3.0 (Hall 1999). LSU and ITS sequences were combined using SequenceMatrix 1.7.8 (Vaidya et al. 2011). Sequences generated in this study were deposited in GenBank (Table 1).
The fasta files were converted to formats required for the AliView program (Larsson 2014), PHYLIP for maximum likelihood analysis (ML), and NEXUS for Bayesian inference (BI). Phylogenetic analyses were performed through the CIPRES science Gateway CIPRES science Gateway V. 3.3 (https://www.phylo.org/portal2/home.action; Miller et al. 2010). Maximum likelihood analysis was performed using RAxML-HPC v.8 tool with rapid bootstrap analysis, followed by 1000 bootstrap replicates (Miller et al. 2010; Stamatakis 2014). The final tree was selected from the suboptimal trees of each run by comparing likelihood scores under the GTRGAMMA substitution model. Bayesian analysis was performed in MrBayes 3.2.7a (Ronquist et al. 2012). The best-fit substitution model GRT + I + G was decided for all two genes by MrModeltest 2.3 under the Akaike Information Criterion (AIC) (Nylander 2004). The Markov Chain Monte Carlo (MCMC) sampling approach was used to calculate posterior probabilities (PP) (Rannala and Yang 1996; Huelsenbeck 2001; Zhaxybayeva and Gogarten 2002). Four simultaneous Markov chains were run for 1 million generations, with trees sampled every 100 generations, resulting in 10,000 trees. The first 2,000 trees, representing the burn-in phase of the analyses, were discarded, and the remaining trees were used for calculating posterior probabilities (PPs) in the majority rule consensus tree (Larget and Simon 1999).
Phylogenetic trees were visualized using FigTree v.1.4.4 (Rambaut 2014), and the layouts were reorganized using the methods described by Xie et al. (2023) and finalized with Adobe Illustrator CS6 software (Adobe Systems, USA). Sequences generated from our collections were deposited in GenBank and are listed in Table 1. Decisions regarding the discovery of new species or records were made following the guidelines of Maharachchikumbura et al. (2021).
Phylogenetic analysis results
The partial LSU-ITS nucleotide sequences were used to determine the phylogenetic position of the new taxa within the family Chaetosphaeriaceae. The concatenated sequence matrix comprises 157 ingroup taxa of Chaetosphaeriaceae and two outgroup taxa, Leptosporellaarengae (MFLUCC 15–0330) and L.bambusae (MFLUCC 12–0846). After alignment, the dataset contained 1,450 characters (LSU: 861 bp, ITS: 589 bp), including 853 distinct alignment patterns, with 11.93% comprising undetermined characters or gaps. Base frequencies were as follows: A = 0.224314, C = 0.274605, G = 0.307808, and T = 0.193272. Substitution rates were AC = 1.327038, AG = 1.998330, AT = 1.575283, CG = 0.648947, CT = 6.385392, and GT = 1.000000, with a tree length of 12.245369. The distribution shape parameter (α) was calculated as 0.317788. The ML and BI trees displayed similar topologies with no significant differences. The best-scoring RAxML tree is shown in Fig. 1, with a final likelihood value of -31034.684968.
The phylogenetic tree generated from ML analysis is based on a concatenated LSU-ITS dataset for the Chaetosphaeriaceae family. Bootstrap support values for ML equal to or greater than 75% and Bayesian posterior probabilities (PPs) equal to or greater than 0.95 were indicated above or below the nodes as ML/PP. Leptosporellaarengae (MFLUCC 15–0330) and L.bambusae (MFLUCC 12–0846) were selected as the outgroup taxa. The newly obtained sequences are indicated in red.
Our two isolates were identified as Pseudolomaanthathailandica gen. et sp. nov. and Submultiguttulisporamultiseptata gen. et sp. nov. in Chaetosphaeriaceae. Pseudolomaantha shares a sister relationship with a clade comprising Caliciastrum, Caligospora, and Craspedodidymum, while Submultiguttulispora forms a separate clade within Chaetosphaeriaceae that is close to Multiguttulispora. Both genera represent distinct, independent lineages and do not belong to any existing genera within Chaetosphaeriaceae.
Taxonomy
Pseudolomaantha
Taxon classificationFungiChaetosphaerialesChaetosphaeriaceae
J.Y. Zhang, Y.Z. Lu & K.D. Hyde gen. nov.
FE3D2B07-D7F0-54F0-A30E-7C0B557B6184
Index Fungorum: IF903140
Facesoffungi Number: FoF16983
Etymology.
The name refers to the new genus’s similarity to the genus “Lomaantha”.
Type species.
Pseudolomaanthathailandica J.Y. Zhang, Y.Z. Lu & K.D. Hyde
Description.
Saprobic on dead stems of bamboo in terrestrial habitats. Sexual morph Undetermined. Asexual morph Colonies on natural substrate, effuse, scattered, hairy, dark brown, glistening. Mycelium partly immersed, composed of brown hyphae. Conidiophores macronematous, mononematous, cylindrical, straight or slightly flexuous, septate, dark brown to pale brown. Conidiogenous cells integrated, terminal, holoblastic, monoblastic, cylindrical, brown or pale brown at the apex. Conidia acrogenous, solitary, rostrate, tapering to the round apex, truncate at base, straight or slightly curved, septate, with distoseptate, pale brown to dark brown; with a gold and glistening sheath near the apex.
Pseudolomaantha
thailandica
Taxon classificationFungiChaetosphaerialesChaetosphaeriaceae
J.Y. Zhang, Y.Z. Lu & K.D. Hyde sp. nov.
56EE1EE2-14B5-52C6-93B4-2A6E624529EA
Index Fungorum: IF903138
Facesoffungi Number: FoF16984
Etymology.
The name refers to the country “Thailand” from where the holotype was collected.
Holotype.
MFLU 24-0394.
Description.
Saprobic on dead stems of bamboo in a terrestrial habitat. Sexual morph Undetermined. Asexual morph Hyphomycetous. Colonies on natural substrate superficial, effuse, scattered, hairy, dark brown, with gold glistening on the apex of conidia. Mycelium partly immersed, partly superficial, composed of septate, mostly unbranched, smooth, brown hyphae. Conidiophores 176–275 × 6–9(–11) µm (x̄ = 219.6 × 7.5 µm, n = 20), macronematous, mononematous, solitary, cylindrical, straight or slightly flexuous, septate, black at the base, paler to light brown or brown towards the apex. Conidiogenous cells 12–22 × 5.5–7 µm (x̄ = 16.4 × 6.1 µm, n = 20), integrated, terminal, holoblastic, monoblastic, cylindrical, brown or pale brown at the apex. Conidia (92.5–)95–112.5 × 12.5–15.5 µm (x̄ = 105.8 × 13.8 µm, n = 25), acrogenous, solitary, dry, pyriform to obclavate, rostrate, tapering to the round apex, truncate at base, basal cell conical-truncate, straight or slightly curved, up to 12-septate, with distoseptate, not constricted or slightly constricted at septum, guttulate, brown, two upper cells subhyaline to hyaline, with gold and glistening appendages around the apex of the conidia.
Pseudolomaanthathailandica (MFLU 24–0394, holotype) a the host substrate b, c colonies on the host substrate d–g conidiophores with conidiogenous cells h–k conidia l, m conidial appendage n, o pure culture from front and reverse. Scale bars: 200 µm (b); 100 µm (c); 50 µm (d–k); 20 µm (l, m).
Culture characteristics.
Conidia germinating on WA within 15 h and germ tube produced from the ends of conidia. Colonies growing on PDA, reaching 22–26 mm in 20 days at 26 °C, circular, edge entire, umbonate with a knobby protuberance, white from above; zonate, yellowish orange in the center, grayish olive to yellowish towards to margin from below.
Material examined.
Thailand • Chiangmai Province, Mushroom Research Center (MRC), on dead stems of bamboo, 11 September 2020, H.W. Shen, Y205-1 (MFLU 24–0394, holotype), ex-type living culture, MFLUCC 24–0521.
Notes.
BLAST results for the ITS and LSU sequence data of Pseudolomaanthathailandica show 88.39% similarities with Caligosporadilabens (CBS 735.83) and 97.81% similarities with Craspedodidymumelatum (NN042874), respectively. Phylogenetic analysis shows that Pseudolomaanthathailandica forms a distinct lineage basal to Caliciastrum, Caligospora, and Craspedodidymum with statistical support (79% ML/0.97 PP, Fig. 1). Members of Caliciastrum, Caligospora, and Craspedodidymum are characterized by phialidic conidiogenous cells with open, vase-shaped collarettes, and brown or hyaline conidia. In contrast, our new species has a sporidesmium-like asexual morph with non-phialidic conidiogenous cells (Figueroa et al. 2018; Wu and Diao 2022; Réblová and Nekvindová 2023). Morphologically, Pseudolomaantha resembles Lomaantha in having macronematous, mononematous conidiophores, integrated holoblastic conidiogenous cells, and acrogenous, obclavate, rostrate, distoseptate, pale brown to brown conidia (Wu and Zhuang 2005; Wu and Diao 2022; Réblová and Nekvindová 2023). However, the two genera are phylogenetically distinct. Additionally, Lomaantha species have conidiogenous cells that are determinate or extend percurrently a few times, as well as conidia that lack or bear filiform, extended, simple or branched apical appendages and distinct septal pores (Wu and Zhuang 2005; Wu and Diao 2022; Réblová and Nekvindová 2023). In contrast, Pseudolomaantha has determinate conidiogenous cells, conidia with golden, glistening appendages at the conidial apex, and lack distinct pores in the distosepta. Based on the combination of morphological and phylogenetic evidence, Pseudolomaantha is introduced as a new genus to accommodate P.thailandica within Chaetosphaeriaceae.
Submultiguttulispora
Taxon classificationFungiChaetosphaerialesChaetosphaeriaceae
J.Y. Zhang, Y.Z. Lu & K.D. Hyde gen. nov.
5B3B2579-A7C9-5351-9F53-A2DE2E3C8964
Index Fungorum: IF903141
Facesoffungi Number: FoF16985
Etymology.
The name refers to the new genus’s close affinity with the genus “Multiguttulispora”.
Type species.
Submultiguttulisporamultiseptatum J.Y. Zhang, Y.Z. Lu & K.D. Hyde.
Description.
Saprobic on dead wood. Sexual morph Undetermined. Asexual morph Colonies on natural substrate, effuse, single, or gregarious, brown to black. Mycelium partly immersed, composed of brown hyphae. Conidiophores macronematous, mononematous, single or in small groups, septate, dark brown at the base becoming light brown towards the apex. Conidiogenous cells integrated, mono- to polyphialidic, terminal to lateral, with funnel-shaped collarettes, cylindrical to cylindrical-lageniform, brown to pale brown to subhyaline towards the apex. Conidia acropleurogenous, septate, pale brown to olive green to brown, with subhyaline cells at both ends of the conidia, fusiform, or ellipsoidal-fusiform, with a filiform appendage at each end.
Submultiguttulispora
multiseptata
Taxon classificationFungiChaetosphaerialesChaetosphaeriaceae
J.Y. Zhang, K.D. Hyde & Y.Z. Lu sp. nov.
B30C6EE3-0BDC-50E5-9426-327E23573FCC
Index Fungorum: IF903139
Facesoffungi Number: FoF16986
Etymology.
The name refers to the multi-septate conidia of the new species.
Holotype.
HKAS 129868.
Description.
Saprobic on a dead wood log by a stream. Sexual morph undetermined. Asexual morph Hyphomycetous. Colonies on natural substrate superficial, effuse, single, or gregarious, arise in groups from knots of hyphal cells, brown to black. Mycelium partly superficial, partly immersed, composed of septate, pale brown to brown, smooth-walled hyphae. Conidiophores 285–385(–533) µm long × 5–7 µm wide at the base (x̄ = 341 × 6 µm, n = 15), macronematous, mononematous, single or clustered in groups, erect, straight or flexible, unbranched, septate, smooth, guttulate, dark brown or black at the base, becoming pale brown towards the apex. Conidiogenous cells 64.5–100 × 4.3–6.1 µm (x̄ = 80.2 × 5.2 µm, n = 15), mono- to polyphialidic, with discrete, terminal to lateral phialides, integrated, terminal, with lateral openings formed by successive sympodial elongation, cylindrical to cylindrical–lageniform, with funnel-shaped collarettes, smooth-walled, guttulate, brown at the base and becoming pale brown to subhyaline towards the apex. Conidia 33–40 × 7.5–9 µm (x̄ = 36.6 × 8.3 µm, n = 20), acropleurogenous, 5(–6)-septate, not constricted at the septum, pale brown to olive green to brown, with subhyaline cells at both ends, straight, sometimes slightly curved, occasionally guttulate, fusiform, or ellipsoidal-fusiform, with a filiform, short and hyaline appendage at each end.
Submultiguttulisporamultiseptata (HKAS 129868, holotype) a colonies on the host substratum b, c conidiophores d–g conidiogenous cells (arrows showing conidiogenous loci) h–p conidia q pure culture from front and reverse. Scale bars: 100 µm (b, c); 20 µm (d–p).
Culture characteristics.
Conidia germinating on WA within 15 h and germ tube produced from conidia. Colonies growing on PDA, reaching 35–40 mm diameter in 15 days at 26 °C, circular with slightly irregular edge, flat with a protuberance in the center, dry, velvety, zonate, tephrosiousto to grey from center to margin; dark brown or black from below.
Material examined.
China • Hainan Province, Wuzhishan City, Wuzhishan Tropical Rainforest Scenic Area, on a dead wood log by a stream, 15 August 2021, J.Y. Zhang, WZ44-1 (HKAS 129868, holotype; GZAAS 23–0763, isotype); ex-type living cultures, KUNCC 23–14145.
Notes.
Based on a BLASTn search in GenBank, the ITS and LSU sequences of our new collection show 91.92% and 95.35% similarity to Phialogeniculataguadalcanalensis (NN044662) and Multiguttulisporatriseptata (IMI 353690), respectively. The phylogenetic tree indicates that our new isolate forms a distinct lineage closely related to Multiguttulispora, without statistical support. This lack of support may be attributed to the absence of molecular sequences of many close phylogenetic relatives, which remain undiscovered (Hyde et al. 2024c). Submultiguttulispora shares similarities with Multiguttulispora in the absence of setae and the presence of macronematous conidiophores with polyphialidic conidiogenous cells that exhibit sympodial extension. Both genera produce septate conidia with a filiform, hyaline appendage at each end. However, Submultiguttulispora is distinguished from Multiguttulispora by its fusiform or ellipsoidal-fusiform, dematiaceous conidia, whereas the conidia of Multiguttulispora are cylindrical, oblong, and hyaline. Based on these morphological and phylogenetic differences, a new genus, Submultiguttulispora, is introduced to accommodate our new isolate, S.multiseptata.
Discussion
In this study, Pseudolomaanthathailandica gen. et sp. nov. and Submultiguttulisporamultiseptata gen. et sp. nov. were introduced based on morphological characteristics and phylogenetic analyses. These two species exhibit non-phialidic and phialidic asexual morphs, respectively. The introduction of these new taxa further highlights the richness and diversity of anamorphic chaetosphaeriaceous fungi (Réblová et al. 2021a, b, c, d, e; Wu and Diao 2022).
The characteristics of conidiophores, conidiogenous cells, and conidia are particularly important for delimiting asexual genera in Chaetosphaeriaceae, along with the presence or absence of appendages (Réblová et al. 1999, 2021b, c, d; Lin et al. 2019; Zheng et al. 2020; Wu and Diao 2022). A significant number of anamorphic chaetosphaeriaceous genera produce hyaline or subhyaline conidia in various shapes, often with filiform, hyaline setulae at the ends, as seen in genera like Arcuatospora, Codinaea, and Kinochaeta (Hughes and Kendrick 1968; Réblová et al. 2020, 2021b, c; Wu and Diao 2022; Hyde et al. 2024b). In contrast, many hyphomycetous genera with dematiaceous conidia lack setulae, such as Catenularia, Phaeodischloridium and Sporoschisma (Goh et al. 1997; Yang et al. 2016; Réblová et al. 2021e; Wu and Diao 2022). Submultiguttulisporamultiseptata gen. et sp. nov. resembles other anamorphic chaetosphaeriaceous genera in having phialidic conidiogenous cells and conidia with filiform, hyaline setulae at both ends (Réblová and Gams 2000; Liu et al. 2016; Lin et al. 2019; Wu and Diao 2022). However, it is distinct in its well-developed conidiophores, polyphialidic conidiogenous cells, and pale brown to olive green to brown, septate conidia with hyaline setulae at each end. The latest key to phialidic asexual genera in Chaetosphaeriaceae was provided by Wu and Diao (2022).
Wu and Diao (2022) recognized ten hyphomycetous genera with non-phialidic anamorphs in Chaetosphaeriaceae, viz., Aunstrupia, Ellisembia, Falholtia, Linkosia, Lomaantha, Morrisiella, Paliphora, Riisgaardia, Stanjehughesia, and Zanclospora. Subsequently, a new non-phialidic genus, Neocirrenalia, characterized by dark brown or black helicoid conidia, was added to this family (Meyers and Moore 1960; Somrithipol et al. 2002; Yang et al. 2023). Recently, Delgado et al. (2024) reclassified Ellisembia into Sporidesmiaceae (Sordariomycetes) based on analyses of a newly collected type species, E.coronata, and expanded and emended Lomaantha to include related ellisembia-like taxa within a monophyletic lineage in Chaetosphaeriaceae. Currently, Chaetosphaeriaceae comprises 10 non-phialidic hyphomycetous genera. Most of these genera are sporidesmium-like, with exceptions such as Neocirrenalia (a helicosporous genus) and Paliphora, which is characterized by setiform conidiophores, polytretic conidiogenous cells, and subfusiform to subacerose, hyaline conidia (Gusmão et al. 2008; Shenoy et al. 2010; Goh et al. 2014; Malosso et al. 2017; Wu and Diao 2022; Yang et al. 2023; Ma et al. 2024). In this study, we introduced a new genus, Pseudolomaantha, which also exhibits a sporidesmium-like asexual morph, characterized by well-developed, solitary or clustered conidiophores and pyriform to obclavate conidia with a glistening gold appendage around the apex, but is phylogenetically distinct. A key to hyphomycetous genera with non-phialidic anamorphs is provided herein.
Key to hyphomycetous genera with non-phialidic anamorphs
**: **
Supplementary Material
XML Treatment for Pseudolomaantha
XML Treatment for Pseudolomaantha thailandica
XML Treatment for Submultiguttulispora
XML Treatment for Submultiguttulispora multiseptata
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Calabon MS Hyde KD Jones EBG Bao DF Bhunjun CS Phukhamsakda C Shen HW Gentekaki E Al Sharie AH Barros J Chandrasiri KSU Hu DM Hurdeal VG Rossi W Valle LG Zhang H Figueroa M Raja HA Seena S Song HY Dong W El-Elimat T Leonardi M Li Y Li YJ Luo ZL Ritter CD Strongman DB Wei MJ Balasuriya A (2023) Freshwater Fungal Biology.Mycosphere 14(1): 195–413. 10.5943/mycosphere/14/1/4 · doi ↗
- 2Chomnunti P Hongsanan S Aguirre-Hudson B Tian Q Peršoh D Dhami MK Alias AS Xu J Liu X Stadler M Hyde KD (2014) The sooty moulds.Fungal Diversity 66(1): 1–36. 10.1007/s 13225-014-0278-5 · doi ↗
- 3Delgado G Koukol O Maciá-Vicente JG Colbert W Piepenbring M (2024) Redefining Ellisembia sensu stricto with a reassessment of related taxa in Sordariomycetes.Mycological Progress 23(1): 1–25. 10.1007/s 11557-024-01967-z · doi ↗
- 4Ellis MB (1971) Dematiaceous hyphomycetes. Commonwealth Mycological Institute. Kew, England, 1–608. 10.1079/9780851986180.0000 · doi ↗
- 5Ellis MB (1976) More dematiaceous hyphomycetes. CABI Publishing, Kew. 10.1079/9780851983653.0000 · doi ↗
- 6Fernández FA Huhndorf SM (2005) New species of Chaetosphaeria, Melanopsammella and Tainosphaeria gen. nov. from the Americas.Fungal Diversity 18: 15–57.
- 7Figueroa R Bran M Morales O Morataya E Minter DW Castañeda-Ruiz RF (2018) Craspedodidymumguatemalense sp. nov. from Guatemala.Mycotaxon 133(2): 315–322. 10.5248/133.315 · doi ↗
- 8Goh T Ho W Hyde K Umali T (1997) New records and species of Sporoschisma and Sporoschismopsis from submerged wood in the tropics.Mycological Research 101(11): 1295–1307. 10.1017/S 0953756297003973 · doi ↗