Revealing the status of Orbicella: Main reef-builder of Morrocoy National Park and Cuare Wildlife Refuge, Venezuela, Southern Caribbean
Anaurora Yranzo-Duque, Ana Teresa Herrera-Reveles, Estrella Villamizar, Francoise Cabada-Blanco, Jeannette Pérez-Benítez, Hazael Boadas, José G. Rodríguez Quintal, Carlos Pereira, Samuel Narciso, Freddy A. Bustillos, Erik Caroselli, Erik Caroselli, Erik Caroselli

TL;DR
This study assesses the population status of two key reef-building corals in two Venezuelan marine protected areas, highlighting their importance and the threats they face.
Contribution
The study provides new population data for Orbicella corals in under-researched southern Caribbean regions, supporting conservation efforts.
Findings
Orbicella faveolata was more abundant and had higher live cover than Orbicella annularis in both MPAs.
The central sector of Morrocoy and Cuare reefs showed the highest abundance of reproductive colonies.
Diseases were identified as the main threat to both coral species.
Abstract
Reef-building corals are the main basis of coral reef ecosystems, and the Orbicella genus is currently the most important in the Caribbean region. Although Orbicella species have been extensively studied, gaps in some southern Caribbean areas still exists on their status, which is crucial for management and conservation plans. In this study, we aimed to describe the population status of Orbicella faveolata and Orbicella annularis in two coastal Marine Protected Areas (MPA) of Venezuela: Morrocoy National Park and Cuare Wildlife Refuge. Between 2018-2020, 16 reefs in five sectors were surveyed using the Atlantic and Gulf Rapid Reef Assessment Protocol. Among the two Orbicella species, O. faveolata was dominant in both MPA´s with higher densities and live cover, varying at both reefs and sectors. The central sector of Morrocoy and reefs from Cuare are the most relevant for Orbicella…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
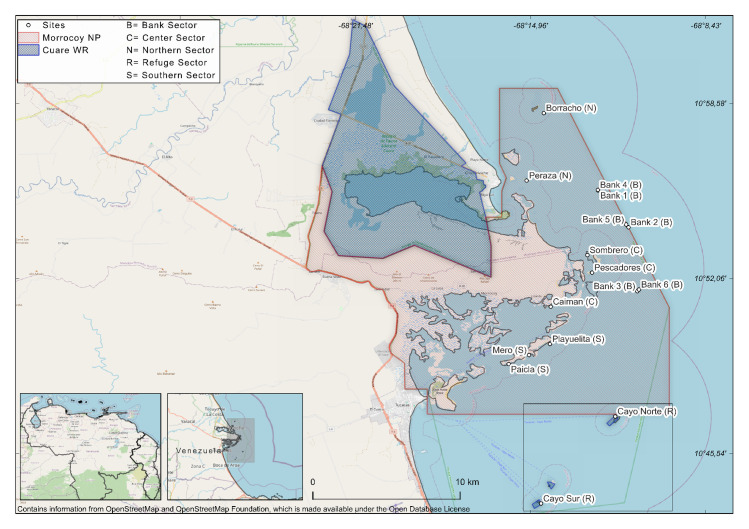 Fig 1
Fig 1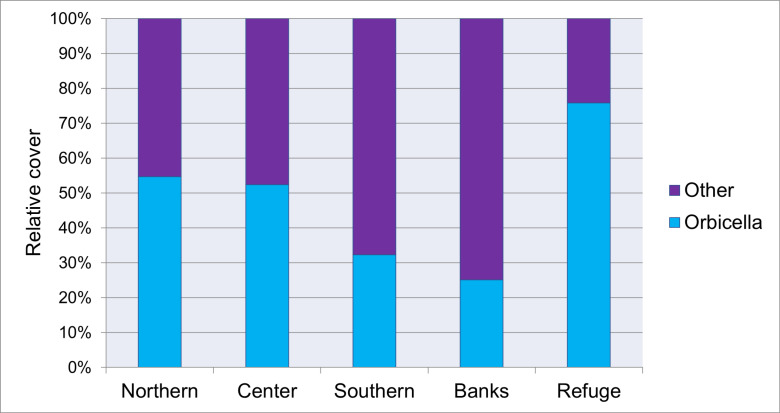 Fig 2
Fig 2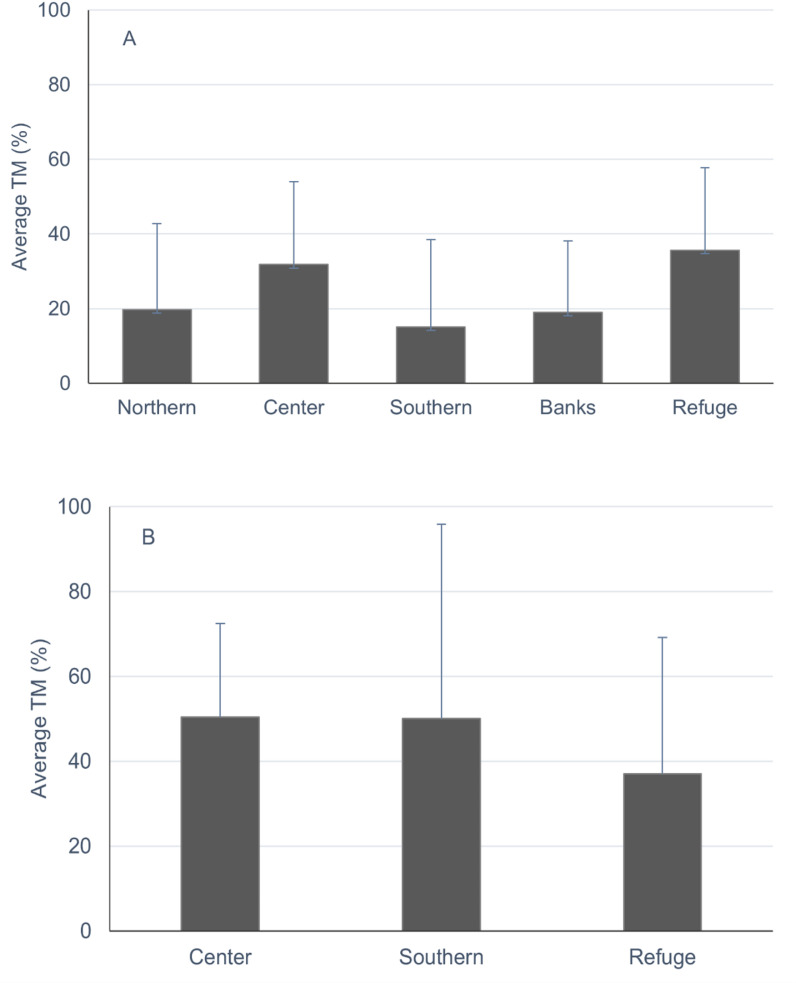 Fig 3
Fig 3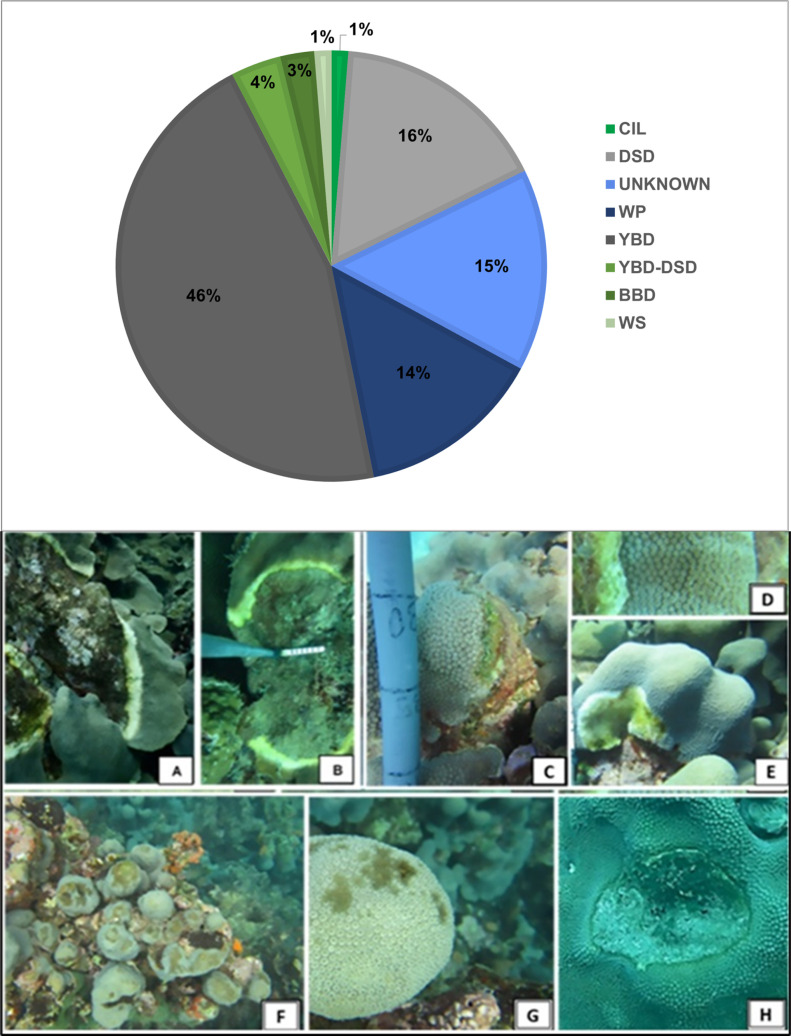 Fig 4
Fig 4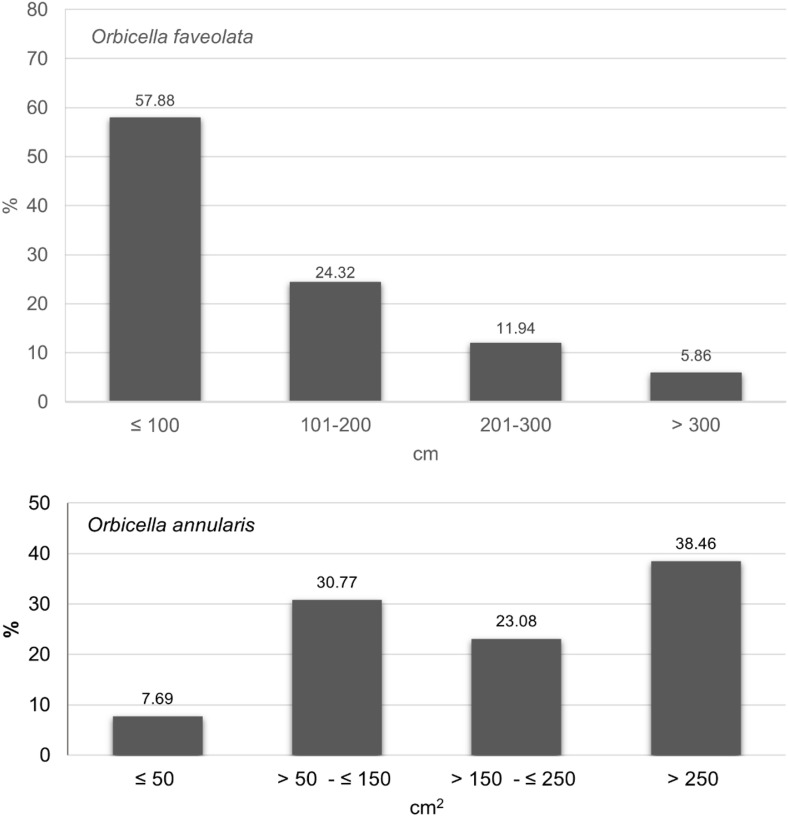 Fig 5
Fig 5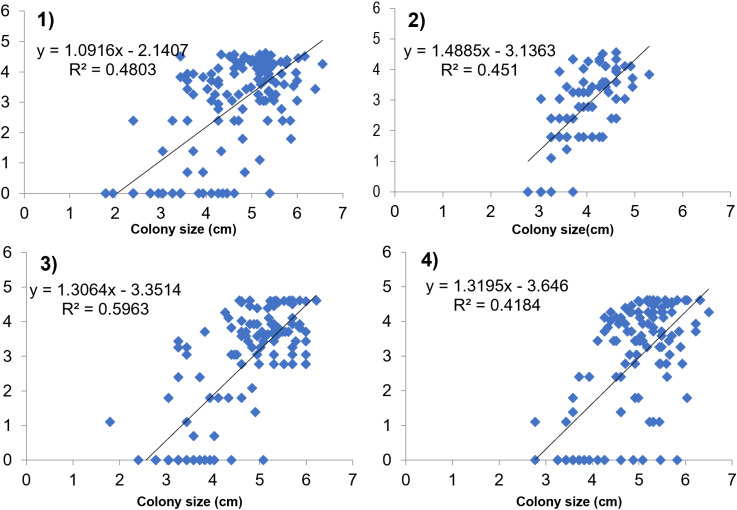 Fig 6
Fig 6- —http://dx.doi.org/10.13039/100009470Zoological Society of London
- —Clear Reef Social Fund
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsCoral and Marine Ecosystems Studies · Marine and coastal plant biology · Marine and fisheries research
Introduction
The relevance of coral reefs globally is extensively documented [1,2] as is that of the coral species responsible for building most of the structural reef framework [3,4]. As such, reef-building corals are key for the ecosystem function of coral reefs and therefore, for maintaining the many benefits they provide to human populations [5]. After the decline of Acropora species in the early 1980s due to white band disease episodes, along with other stressors [6], Orbicella was considered the most important genus of shallow reef building corals in the Caribbean region [7,8]. Orbicella species crucially contribute to reef carbonate production rate and the structural complexity provided by their colonies, enhances habitat heterogeneity [9,10]. They are distributed throughout over 30 countries in the Caribbean, including southern Florida, Bahamas, Bermuda and Gulf of Mexico. Nevertheless, their populations, especially Orbicella annularis and Orbicella faveolata, have experienced a drastic decline caused by bleaching events, disease episodes, and other negative impacts caused by anthropic activities [11,12] evidenced by a reduction in live coral cover and colony abundance [5,13]. Currently, O. annularis and O. faveolata are classified as Endangered according to the last International Union for Conservation of Nature (IUCN) Red List of Threatened species assessment [14]. They are also included in CITES (Appendix II), the SPAW Protocol (Annex III), and in 2014, the National Marine Fisheries Services from the United States (NMFS) included them under the Endangered Species Act (ESA) highlighting the need for conservation action across the species’ range.
Implementing local conservation measures for endangered species requires a good understanding of the specie’s status locally. In Venezuela, a southern Caribbean country, information on Orbicella species is limited to some areas. The most recent publication dates back to 2015, which is part of the Red Book of Venezuelan Wildlife, a National level Red List of threatened species assessment, where O. annularis was classified as Vulnerable and O. faveolata as Least Concern [15]. Here we describe the population status of O. faveolata and O. annularis during 2018-2020, in the coastal Marine Protected Areas (MPA) Morrocoy National Park (MNP) and Cuare Wildlife Refuge, the only true fully carbonate coastal reefs in the country, which are considered the most developed of the continental area of Venezuela [16]. Extensive mangrove forests, seagrass meadows, and reef patches harbor high biodiversity in both protected areas [17,18]. Surrounding communities depend directly on the MPAs ecosystems, as their main economic activities are tourism and artisanal fishing. However, restricted activities are established in the Zoning Management Plans, which includes a higher access restriction in Cuare than in Morrocoy (only some tourism activities allowed), as the Wildlife Refuge have a stricter legal basis than National Parks in the country.
In 1996, a massive die-off resulted in a 90% mortality of the benthic fauna in MNP, including a drastic drop in live coral cover and diversity [19]. The die-off is believed to have been caused by a climatic and oceanic anomaly that caused a sudden oxygen depletion or a chemical pollution [20]. Additionally, chronic anthropogenic pressures have affected MNP for decades [21], including unregulated tourism, high sedimentation, sewage discharges, and pollution from nearby oil refinery and petrochemical facilities [22,23]. Unlike Morrocoy, coral reefs in Cuare were unaffected by the massive die-off, possibly due local circulation patterns [24].
This study is the first to focus on the ecology of Orbicella in this area of the southern Caribbean region. We specifically assessed spatial differences in abundance and other species attributes at reef scales as well as across sectors to better inform area-based conservation measures.
Materials and methods
Study area and data collection
Morrocoy National Park and Cuare Wildlife Refuge are coastal Marine Protected Areas in the central-west of Venezuela (Fig 1) and the last is included at the list of the Convention on Wetlands of International Importance, known as the Ramsar Convention (RAMSAR site).
Study sites in Morrocoy National Park and Cuare Wildlife Refuge, Falcón State, Venezuela, located in the Southern Caribbean.The assessed reefs (n = 16) were classified into five sectors, indicated by the letters in parentheses next to each reef name. In the southern area of the Cuare Wildlife Refuge, only the coral cays within its boundaries are included, which are highlighted within a square frame. All sites were fringing reefs, except for the banks, which were areas dominated by branched octocorals and some sponges, along with isolated corals. These banks formed mosaics with sandy bottoms, patches of coral rubble, and rocky slabs. The base map was created using OpenStreetMap (https://www.openstreetmap.org). Transects/sector: Banks (n = 18), Center (n = 36), Northern (n = 25), Refuge (n = 24); Southern (n = 37). Transect/survey: July 2018 (n = 38), November 2018 (n = 36), July 2019 (n = 36), January 2020 (n = 30).
The status of Orbicella was assessed considering the following variables: density, live coral cover, size structure, and mortality in 16 reefs across five sectors (Fig 1) which were selected based on a previous distribution assessment and considered to encompass the best spatial representation of the area. A colony was defined as one entity of coral skeleton with living tissue even with separate patches of living tissue by partial mortality, but morphologically still one entity [25].
In Morrocoy National Park we assessed eight fringing reefs: two at the northern, three at the center and three at the southern of the National Park. Additionally, six banks reefs were included (Fig 1). In Cuare Wildlife Refuge, two of the three fringing reefs of this site were included.
We use a modified version of the AGRRA Protocol 5.5 (Atlantic and Gulf Rapid Reef Assessment) [26] from which a general benthos characterization was obtained. We did a continue record of substratum cover under the transect line, instead of recording at 10 cm intervals points under the line, as the current protocol. On each reef, 10 m long x 1 m width belt transects were generally placed between 8 and 11 m depth [see 26 for further details). Four surveys were done: July and November 2018, July 2019, and late January 2020, totaling 141 transects (see details in Fig 1). The study was approved under permit 35114 of the Ministry of Popular Power for Ecosocialism.
Data analysis
Live coral cover, mortality and diseases.
The data was analyzed using Primer 6 V 6.1.16 & PERMANOVA + V1.0.6. Permutational analysis of variance (PERMANOVA) was used to analyzed i) live cover of both Orbicella species and ii) tissue mortality categories (recent, old, and total). The PERMANOVA mixed-effect linear model was done using two fixed factors (sampling period and sector) and a random factor (reef) nested within sector based on Euclidian distance.
A contingency analysis (Pearson’s chi-square) was used to assess differences in Orbicella diseases between sectors and survey periods. Diseases were classified according to literature gross lesion descriptions [27,28]. Disease prevalence was estimated as the number of diseased colonies/total number of colonies x 100.
Colonies were classified as alive (≤80% of partial mortality), almost dead (between > 80-<100% of partial mortality), and dead (100% mortality-standing dead), following the criteria of [12].
Colonies size structure.
Only colonies with a diameter of > 4 cm were considered, as juveniles were ≤ 4 cm in size [29]. Size frequency distribution was built for both species. For O. annularis, the categories of [5] were used: size class I = ≤ 50 cm^2^; size class II = > 50 cm^2^- ≤ 150 cm^2^; size class III = > 150 cm^2^ - ≤ 250 cm^2^; size class IV = > 250 cm^2^. For O. faveolata the size of the colonies was plotted considering the reproductive size reported: > 100 cm maximum diameter [30]. The reproductive size reported for O. annularis is > 200 cm^2^ area according to [31]. A Permutational analysis of variance (PERMANOVA) was performed to test differences in the density of reproductive and non-reproductive colonies between sectors. In addition, an analysis of lineal regression was used to estimate the relationship between total mortality (%) and size of O. faveolata colonies, and an analysis of variance was used to assess the lineal regression model fits, after testing assumptions using Shapiro-Wilk normality test, Breusch-Pagan test and Durbin-Watson test.
Results
Among the recorded coral genera, Orbicella had the highest relative percent cover within all sectors (Fig 2), with the exception of the banks, where Millepora spp. had a greater relative cover (34.81%). Live coral cover was generally dominated by O. faveolata (between 22.67% at the Banks and 66.63% at the Refuge), and significant spatial differences were found between sectors (F = 10.277, df = 4, p = 0.0036, CV = 35.73%) but not through time (S1 Table). The Banks and Southern sectors had the lowest cover of O. faveolata (t ≥ 3.00, df = 4, p < 0.05) while the highest percentage cover was recorded in the Refuge sector. There were no significant differences between the central and northern sectors (t ≤ 2.93, df = 4, p = 0.10). We found O. annularis in seven of the 16 reefs surveyed, including both reefs from Cuare, and in Morrocoy, two reefs of the southern, two of the center and one of the northern sectors. For this species, live cover was higher in Cuare than in the other sectors surveyed (t ≥ 3.00; df = 4, p < 0.05, average cover = 5.5%). It is important to highlight that in a shallow area of Sombrero reef (center sector) there is an extensive area of O. annularis colonies, unique along all the reefs surveyed. Previous reports related to Orbicella live cover at the study site are summarized in S2 Table. During the study period, a total of 30 colonies for O. annularis and 647 for O. faveolata were recorded*.*
Relative cover (%) of Orbicella: O. annularis and O. faveolata.Study site: Morrocoy National Park (Northern, Center, Southern, Bank sectors) and Cuare Wildlife Refuge (Refuge) in 2018-2020, Venezuela, southern Caribbean.
Concerning colony health, 85.88% of O. faveolata and 73.68% of O. annularis colonies fell within the alive category ( ≤ 80% of partial mortality). Average total mortality varied between 15.11% ( + 23.42) in the Southern and 35.69% ( + 22.04) in the Refuge for O. faveolata and 37.08% (+32.01) to 50.42% (+22.00) in the Refuge and Center for O. annularis (Fig 3). Tissue mortality categories showed differences between sectors (S3 Table). Old mortality stage was higher relative to recent mortality, and reefs from the Southern sector had lower percentages of recent mortality, compared to reefs in the central sector (t = 3.68, df = 4, p = 0.02) and Refuge (t = 4.64, df = 4, p = 0.01). Average values of recent mortality were between 0.45% (Banks) to 12.97% (Center) and for all sectors, recent mortality was higher in the last survey (late January 2020) than in the first one in July 2018 (t = 3.53, df = 3, p = 0.0083). For O. annularis there were no spatial differences in mortality categories and the average value of recent mortality for all surveys was 1.81%.
Average total mortality (TM) of O. faveolata (A) and O. annularis alive colonies (B).Study site: Morrocoy National Park (Northern, Center, Southern and Banks) and Cuare Wildlife Refuge (Venezuela). Bars are standard deviation (SD). Alive colonies = colonies with ≤ 80% partial mortality. Period: 2018-2020.
Diseases were the main detrimental factor recorded, with Yellow Band Disease being the most frequently observed in O. faveolata (Fig 4). The density of diseased O. faveolata colonies varied among the reefs (X^2^ = 30.169, df = 4, p = 0.0015), and was lower in the Southern sector. The average prevalence of diseases varies between 2.78% in the North (Borracho reef) and 55% in the Center (Caiman reef). We recorded the highest values mostly in the Center and Refuge sectors, with an average of 25.66% ( ± 28.64) and 24.32% ( ± 22.49), respectively. In general, prevalence was highest in July with less diseased colonies found in January. However, density of the diseased colonies was not significantly different between surveys (X^2^ = 3.425, df = 2, p = 0.1804). We found three diseases in O. annularis: White Plague Disease, Yellow Band disease, and Dark Spot Disease (Fig 4E-G) without differences in density of diseased colonies between reefs and sectors (X^2^ = 4.672, df = 4, p = 0.587). We observed an additional health condition mostly in O. faveolata colonies, a dark line bordering the live tissue adjoining the dead area (Fig 4H). We identified other detrimental factors in fewer colonies, including bioerosion by Cliona sponges, grazing by damselfish (Pomacentridae), bleaching and mechanical damage (2.44% to 8.76% of O. faveolata colonies and 12.5% to 25% of O. annularis colonies).
Orbicella diseases and syndromes in Morrocoy National Park and Cuare Wildlife Refuge, in 2018-2020, Venezuela.Graph showing diseases and syndromes in O. faveolata: CIL: Ciliate, DSD: Dark Spot Disease; WPD: White Plague Disease; YBD: Yellow Band Disease; BBD: Black Band Disease; WS: White Syndrome. Photos above: Orbicella faveolata with Yellow Band Disease (A-B); ciliate infection (C) and White Plague Disease (D); Orbicella annularis with White Plague Disease (E), Photos below: O. annularis with Dark Spot Disease (F-G) and Thin Dark Line in O. faveolata (H).
Of the total alive O. faveolata colonies measured, 42.12% were of adult reproductive size (more than 100 cm maximum diameter, Fig 5 above). Density of O. faveolata reproductive colonies varied among sectors (F = 7.33, p = 0.0209, %CV = 46,37, S4 Table) with higher densities in the Refuge (t ≥ 4.23, df = 4, p ≤ 0.022), followed by the Center sector (t = 1.63, df = 4, p = 0.20), where a high proportion of adult colonies was also found. Meanwhile, the Banks had densities significantly lower than those in the Northern, Central, and Refuge sectors (t ≥ 2.95, df = 4, p ≤ 0.05) and a density similar to that of the Southern sector (t = 1.44, df = 4, p = 0.24). Most of O. annularis colonies, were in size class IV (max. diameter = >250 cm^2^, Fig 5 below). For this species, most colonies (53.84%) had a reproductive size ( > 200 cm^2^ area) and were mainly found in Cuare (85.71%). No Orbicella recruits were observed during the surveys.
Size classes in Orbicella faveolata and O. annularis.Above, N = 444 colonies of O. faveolata with measures; below, N = 13 colonies of O. annularis with measures. from Morrocoy National Park and Cuare Wildlife Refuge (Venezuela). Period: 2018-2020. O. faveolata size classes according to adult reproductive sizes.
Finally, the simple linear regression showed a positive relationship between colony sizes and mortality for O. faveolata at the four periods evaluated (R^2^ = 0.418 – 0.596, F > 0.61, p < 0.00002) (Fig 6, S5 Table). In contrast, O. annularis colony size and total mortality did not show a relationship (R^2^ = 0.006, F = 0.1563, p = 0.698).
Relationships between total mortality (%) and colony size (cm) of O. faveolata for the periods evaluated.1) July 2018 (n = 147), 2) November 2018 (n = 77), 3) July 2019 (n = 136) and 4) January 2020 (n = 120). Lines show the best-fit regression model and fitted functions of the form Total mortality (%) = ß0 + ß1 * colony size (cm). P values for all functions < 0.0001. R-Squared (R² = the coefficient of determination or the goodness of fit).
Discussion
We assessed the status of the main coral reef builders, O. annularis and O. faveolata, in Morrocoy National Park and Cuare Wildlife Refuge in Venezuela (southern Caribbean). The results indicated that O. faveolata is the dominant species for coral reefs in both MPAs. Among the sectors we evaluated, the Center in Morrocoy and the Refuge in Cuare were the most relevant for Orbicellas´s population due to the higher live coral cover and density of reproductive colonies for both species. The reefs where we found Orbicella colonies are in better health, are the same reefs which were reported healthier more than 20 years ago: Sombrero at the Center sector (which had a low mortality as a consequence of the massive die-off event in 1996) and Cayo Norte in Cuare, which was unaffected by the 1996 die-off [19,24,32]. This suggests a localized effect of stressors and threats to the species within the MPA with great potential for both area-based management interventions and source-mitigation of threats to help conserve the species.
Influence of land activities modulated by local circulation has been identified as one of the main threats to Morrocoy National Park’s coral reefs. The least influence of sediment discharges has been related to a better condition of reefs located at the center of Morrocoy National Park [17,24,33], unlike the northern (closest to Tocuyo River) and southern sectors (closest to Aroa river). Sombrero has the highest current speed reported for the National Park (35 cm/second) with a northwards direction [34], which avoids of the plume of sediments coming from the Tocuyo river. In case of Cuare, no direct causes have been identified, but the direction and intensity of the currents could be related [24].
Tissue mortality caused by detrimental factors among which diseases can be highlighted, is a concern for the future of Orbicella. Even when at the time of this study it was not observed the most recent coral disease reported: Stony Coral Tissue Loss disease (SCTLD) for which Orbicella species have been categorized as intermediately susceptible or highly susceptible [35], the recorded diseases are an important threat to their survival.
The dominance of Orbicella species before the mass mortality event in 1996 has been reported for some reefs along the Morrocoy National Park, but data was limited to live cover (see S2 Table for references) which can be a poor indicator of the abundance of a coral species, unlike colony density, especially for Orbicella [11]. Differences in the depth of surveys, as well as cover estimation methods (e.g., use of quadrats, chain transects) creates difficulties for obtaining a clear trend over time for Orbicella species. Nevertheless, taking these studies as a reference, there is a decrease in O. annularis live cover in most reefs, within MNP from 1999 through to 2018-2020, accounting for data in 2000, 2004 and 2005. For O. faveolata, there are no records of live coral cover before the massive die-off; however, considering the most recent record (years 2004-2005), the average live cover estimations for the present study are similar for some reefs. More recently in 2017–2018 [36] reported the dominance of O. faveolata in Caiman and Sombrero keys (center sector) as well as in the three Refuge keys (North, South and Middle).
General trends of Orbicella species in the Caribbean region are exhibiting a decline in live cover and abundance [8,11,37]. However, a status review [38] indicates a variable situation with mixed population trends of both reduction and stability due to the common presence of colonies in some areas (e.g., Florida, Puerto Rico, US Virgin Islands) and scarcity in others (e.g., Cuba). Orbicella species at the study site do not appear to have been replaced by weedy/opportunistic corals, at least in the depth interval evaluated. Only the most deteriorated sites, e.g., the Southern sector had a higher abundance of weedy corals than Orbicella (e.g., Porites) which has not been the common trend within other Caribbean reefs [39,40].
The proportion of adult colonies within an area can indicate their reproductive potential of a local population. However, the effective population size may be smaller than the total standing population because of clonality and low reproductive compatibility [41]. The positive relationship between size and mortality for O. faveolata colonies found here, can compromise the reproductive scope of this species at the study sites, as larger colonies tend to have higher partial mortality, which is common due to more time exposed to different disturbances [42–44] and the loss of living tissue affects the ability of corals to reproduce sexually [9].Smaller colonies in the Southern sector with less mortality, are far from healthier as they are most likely a result of colony fission by partial mortality [45]. Additionally, high levels of recent mortality, like those we observed at some sites is indicative of conditions in which growth and regeneration rates are not able to keep up with mortality, and suggests that a disturbance is ongoing [46,47]. This disturbance may have increased in intensity during the last survey, when´´red flags´´ values above 5% were recorded [48].
During the present study, no Orbicella recruits were observed. The only recruit recorded were of fast-growing species from the genera Agaricia, Porites and Pseudodiploria, which agrees with the findings of a previous study in the area [24] although other studies found recruits and juveniles of Orbicella [49,50]. Low recruitment of Orbicella genus hinders detection of recruits, making the time period of a study an important factor when assessing recruitment [5,11]. According to [29] the low recruitment of Orbicella might be related to high post-settlement mortality or low abundance of settlement cues from coralline algae as was found for Acropora [51] and not to failed reproduction or poor fertilization success [52,53]. This highlights the importance of long-term studies when assessing long-lived animals like Orbicella.
Mortality factors found for Orbicella species in the study site were in line with the literature from the Caribbean region, including Los Roques National Park in Venezuela, with Yellow Band Disease (YBD) and White Plague Disease (WPD) as the most prevalent diseases [8,54–57]. Orbicella’s health assessment has allowed us to obtain a broader vision of this topic as previous records of diseases from the study area were limited to Sombrero in Morrocoy and Cayo Norte in Cuare [58–61,63]. It is important to highlight that histopathology examination of diseased colonies should be done to have robust data on this topic in the study area [63,64]. With respect to the findings of the condition described as Thin Dark Line Syndrome (TDL), a temporal investigation should be performed as it could be an indicative of changes in the microbiome or in the adjacent bacterial biota of the coral (E Jordan-Dahlgren pers. comm., Nov,2022). Another hypothesis is it may be a pigmentation response to the presence of algal mats and sediments (R Rodríguez-Martínez,pers.comm., Nov,2022). Spatial differences obtained for the density of diseased colonies are probably related to density-dependent processes [65] as the reefs with fewer Orbicella diseased colonies had the lowest Orbicella abundance.
Important local stress factors not addressed like overfishing, sediment and sewage discharges, pollution including oil spills, coastal development, and associated water quality degradation are important threats in the study area, which can affect Orbicella species at any stage of their life cycle [66–68]. Reducing local threats is essential, as stress factors can affect coral´s immune system [69,70]. For example [62] studied the effects of hydrocarbon pollution on healthy and diseased colonies of O. faveolata from Sombrero reef and found reduced enzymatic activity in colonies infected with Yellow Band Disease (the most frequent disease recorded in the present study) suggesting that diseased colonies may be more vulnerable to the effects of chemical pollution. In recent years, recurrent oil spills in the area (2020,2023,2024 originating from the oil refinery located near both MPA´s [71,72] could exacerbate the detrimental effects of diseases in Orbicellas’ within the study site. Other significant threats in the area include the invasive octocoral Unomia stolonifera [73] currently present in Cuare (Cayo Sur) as well as the increase in sea temperate as a result of climate change.
In conclusion, our results suggest that reefs from the Cuare Wildlife Refuge and the central sector of Morrocoy National Park should be the focus for conservation of the local Orbicella population to maintain and, if possible, recover the higher density of reproductive colonies and coral cover, relative to the rest of the MPA reefs. Proper management by environmental authorities, including regulating anthropic stressors, is critical for the future of these coral communities. Improving both compliance and enforcement of the current zoning with restricted access to Cuare reefs, is fundamental. Finally, responsible and sustainable tourism must be promoted and fostered in Morrocoy National Park, but especially in the central sector. Tourists and other national park users must be included in awareness raising campaigns to improve the effectiveness of the control and surveillance actions, through improved compliance.
Supporting information
S1 TableOrbicella annularis and Orbicella faveolata live cover in Morrocoy National Park and Cuare Wild life Refuge, Venezuela (period 2018-2020).PERMANOVA under a mixed-effect linear model with two fixed factors (sampling period and sector) and a random factor (reef) nested within sector based on Euclidean distance.(DOCX)
S2 TableSynthesis of previous live cover reports for Orbicella annularis (O.ann) and O. faveolata (O.fav) in Morrocoy National Park (northern, center and southern sectors) and Cuare Wildlife Refuge (Refuge sector) Venezuela.(DOCX)
S3 TableTissue mortality of O. faveolata colonies in Morrocoy National Park and Cuare Wildlife Refuge, Venezuela (2018-2020).PERMANOVA under a mixed-effect linear model using two fixed factors (sampling period and sector) and a random factor (reef) nested within sector based on Euclidian distance.(DOCX)
S4 TableDensity of Orbicella faveolata colonies in reproductive and non-reproductive sizes in Morrocoy National Park and Cuare Wildlife Refuge, Venezuela (2018-2020).PERMANOVA under a linear model with two fixed factors (sampling period and sector) and a random factor (reef) nested to the sector fixed factor.(DOCX)
S5 TableTotal mortality (%) and sizes of Orbicella faveolata colonies in Morrocoy National Park and Cuare Wildlife Refuge, Venezuela (2018-2020).Results for the simple linear regression in each period (1-July 2018, 2-November 2018, 3-July 2019 and 4-January 2020).(DOCX)
S1 FileSupporting information data(XLSX)
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Woodhead AJ, Hicks CC, Norström AV, Williams GJ, Graham NA. Coral reef ecosystem services in the Anthropocene. Funct Ecol. 2019; 33:1023–34. doi: 10.1111/1365-2435.13331 · doi ↗
- 2Santavy DL, Horstmann CL, Sharpe LM, Yee SH, Ringold P. What is it about coral reefs? Translation of ecosystem goods and services relevant to people and their well-being. Ecosphere. 2021;12(8):1–27. doi: 10.1002/ecs 2.3639 34938591 PMC 8686212 · doi ↗ · pubmed ↗
- 3Alvarez-Filip L, Dulvy NK, Gill JA, CôtéIM, Watkinson AR. Flattening of Caribbean coral reefs: region-wide declines in architectural complexity. Proc Biol Sci. 2009;276(1669):3019–25. doi: 10.1098/rspb.2009.0339 19515663 PMC 2817220 · doi ↗ · pubmed ↗
- 4Kuffner IB, Toth LT. A geological perspective on the degradation and conservation of western Atlantic coral reefs. Conserv Biol. 2016;30(4):706–15. doi: 10.1111/cobi.12725 27029403 · doi ↗ · pubmed ↗
- 5Edmunds PJ. A quarter-century demographic analysis of the Caribbean coral, Orbicella annularis, and projections of population size over the next century. Limnol Oceanogr. 2015; 60:840-55. doi: 10.1002/lno.10075 · doi ↗
- 6Aronson RB, Precht WF. White-band disease and the changing face of Caribbean coral reefs. Hydrobiologia. 2001;460:25-38. doi: 10.1023/A:1013103928980 · doi ↗
- 7Knowlton N, Weil E, Weigt LA, Guzmán HM. Sibling Species in Montastraea annularis, Coral Bleaching, and the Coral Climate Record. Science. 1992;255(5042):330–3. doi: 10.1126/science.255.5042.330 17779583 · doi ↗ · pubmed ↗
- 8Bruckner AW, Bruckner R. The recent decline of Montastraea annularis (complex) coral populations in western Curaçao: a cause for concern? Rev Biol Trop. 2006; 54:45–58.