ADAM8 inactivation modulates cellular component assembly gene expression in the mouse intervertebral disc
Ken Chen, Zuozhen Tian, Motomi Enomoto-Iwamoto, Yejia Zhang

Abstract
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
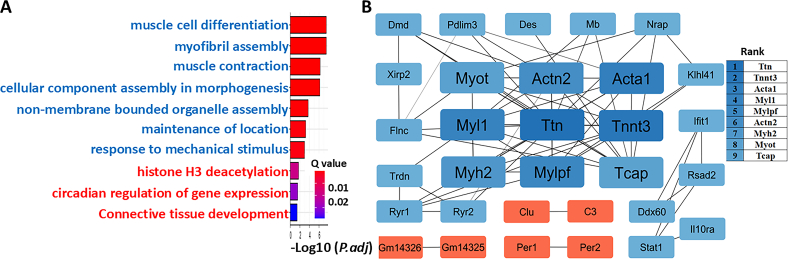 Figure 1
Figure 1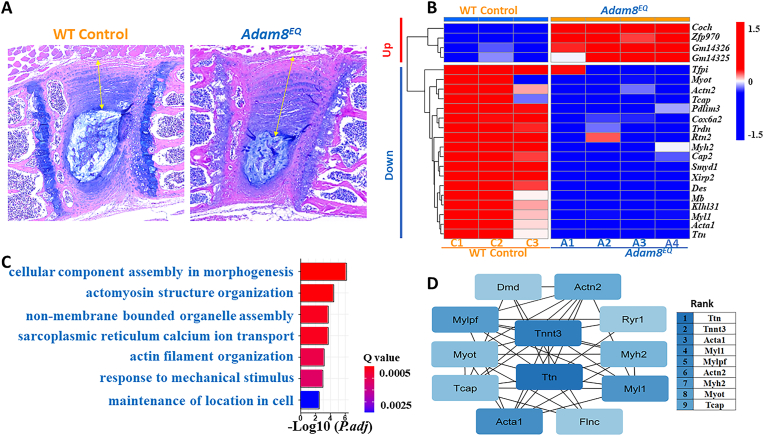 Figure 2
Figure 2Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsSpine and Intervertebral Disc Pathology · Orthopaedic implants and arthroplasty · Osteoarthritis Treatment and Mechanisms
Dear Editor
ADAM8 (ADAM metallopeptidase domain 8, aka MS2/CD156/CD156a) is a membrane-anchored proteinase. ADAM8 and its proteolytic product, fibronectin fragments, are elevated in degenerative human intervertebral discs (IVDs).1 Pathological fibronectin fragments can trigger further disc degeneration. ADAM8 proteolytic activities and fibronectin fragments may accelerate degenerative disc disease, a common clinical problem leading to tremendous socioeconomic burdens in the United States.
An ADAM8-inactivation mouse model has been generated by introducing a point mutation, replacing the Glutamic acid (E) at position 330 with a Glutamine (Q; abbreviated as Adam8^EQ^).2 The E to Q mutation prevents shedding of the prodomain, thereby preventing self-activation of the enzyme. ADAM8 inactivation resulted in higher proteoglycan content and thicker annulus fibrosus in intact discs.4 Despite the morphological differences between the Adam8^EQ^ and wild type (WT) mouse discs (Fig. 1A), no significant gene expression was detected in the limited panel of markers examined previously.3, 4 To examine the alteration of biological pathwayss in response to ADAM8 inactivation, an unbiased approach comparing the transcriptome of IVDs in the mutant and WT mice was used. Messenger RNA was isolated from the Adam8^EQ^ and wild type (WI) mouse IVDs, and bulk RNAseq was performed followed by transcriptomic data analysis. The RNASeq datasets generated and analyzed during this study have been deposited in the NCBI Gene Expression Omnibus (GEO) repository with the accession number [GSE255262](GSE255262).
IVDs in the Adam8^EQ^mutant mice displayed different gene expression profiles. Among the 17,489 genes identified by RNAseq, 94 genes were differentially expressed (p.adj<0.05) comparing Adam8^EQ^ and WT mouse IVDs. The genes were further sorted according to fold changes from high to low, resulting in 25 upregulated and 69 downregulated genes. Four upregulated and 38 downregulated genes with highest fold changes were selected for heatmap, which visualizes clearly different expression profiles between WT and mutant mice (Fig. 1B).Figure 1Differentially Expressed Genes Between Mutant Adam8^EQ^and Wild Type (WT) Control Mouse Intervertebral Discs (IVDs). A: lumbar IVDs stained with Alcian blue; Arrows: indicate thickness of the annulus fibrosus; B: heatmap; each column represents data from one Adam8^EQ^ or WT control mouse IVD.C1-3: WT controls; A1-4: Adam8 mutant; red: upregulated genes; blue: downregulated genes. C: pathway analysis; P.adj: adjusted p-value; Q value: estimated false discovery rate. D: network analysis; rank: genes with larger to smaller number of network connections; Ttn: titin; Acta: actin alpha; Myh: myosin heavy chain; Myl: myosin light chain; Mylpf: Myl phosphorylatable, fast skeletal muscle; Actn: actinin; Myot: myotilin; Tcap: Telethonin.Figure 1
Cellular component assembly pathway was over represented among genes differentially expressed in Adam8^EQ^ and WT control mice*.* A protein list was generated based on fold-changes. The 94 genes with p.adj<0.05 were separated into upregulated genes (Adam8^EQ^/WT ratio>1), and downregulated genes (Adam8^EQ^/WT ratio<1). Gene Ontology (GO) analysis for biological processes was performed with the R software and various R packages. Cellular component assembly was the most overrepresented pathway among downregulated genes (Fig. 1C). When both up- and downregulated genes were analyzed together, muscle differentiation and myofibril assembly were the two most overrepresented biological pathways among downregulated genes. Histone (H)3 deacetylation was the most overrepresented pathway among the upregulated genes (Fig. S1A).
Protein–Protein Interaction (PPI) Network Visualization. The gene list described above was analyzed based on STRING database (https://string-db.org/, Version: 11.5) with the CytoScape software, with minimal interaction score set at high confidence (0.70). Titin (Ttn) has the highest network connection among downregulated genes (Fig. 1D). Interestingly, period (per) 1 and 2 genes, encoding proteins known to drive the circadian rhythm, were among the 25 upregulated genes (Fig. S1B).
ADAM8 is known to cleave fibronectin in IVD tissues, generating pathological fibronectin fragments,1 which may trigger further disc degeneration. Previous examination of a mouse model revealed that IVD tissues with inactivate ADAM8 retained more proteoglycans and had thicker AF.4 In response to injury, the IVD tissues of mutant mice expressed higher levels of type II collagen gene, but also higher levels of inflammatory marker genes including Cxcl1 and Il6.5 In these studies, a candidate marker approach was used, where a panel of markers was selected based on past work by our group and others. No statistically significant gene expression changes were detected among the limited panel of genes studied, despite morphological changes in the intact IVDs of ADAM8-inactivation mice compared with WT controls.3, 4 To elucidate the mechanism of the morphological differences, an unbiased approach was used to compare the entire transcriptome of IVDs in ADAM8-inactivation and WT mice.
Among the 17,489 genes with detectable levels of expression in the mouse IVDs, profound changes in gene expression profiles have been found, with most genes never studied previously in the IVDs (Fig. 1). Cellular component assembly and response to mechanical stimulus pathways were downregulated. We have searched nucleus pulposus cell single cell RNA sequencing data, and found genes with the highest magnitude of changes expressed in various subpopulations of intact NP cells.1 Among the genes downregulated in Adam8^EQ^ mice compared with WT, Ttn (Titin), encodes a large protein, and has the largest protein–protein interaction (PPI). Tfpi (Tissue Factor Pathway Inhibitor) encodes a protein in the coagulation process. Zfp970 (Zinc Finger Protein 756) and Coch (Cochlin) are among the genes most upregulated in the Adam8^EQ^ mouse IVDs. Zfp970 encodes a member of the zinc finger protein family that modulates gene expression, while Coch encodes a protein that plays a role in the control of cell shape and motility. None of the above genes highly regulated by ADAM8-inactivation had been well described in IVDs before the current work. Future directions include real-time PCR and immunostaining to confirm the distribution of the genes and proteins. It is also important to be aware that some genes without detectable level of expression in intact IVDs may be induced in pathological states. Another limitation is that young adult male mice on the DBA were used in this study, and genetic background, sex and age are known to influence levels of gene expression.
In summary, an unbiased bulk RNAseq approach was used to compare the transcriptomes of ADAM8-inactivated and WT control mouse IVDs. Profound differences in the gene expression profiles were detected between the mutant and WT mouse IVDs, which could explain their morphological differences reported previously.4
Data availability statement
The RNASeq datasets generated and analyzed during this study have been deposited in the NCBI Gene Expression Omnibus (GEO) repository with the accession number GSE255262. Additional study data will be made available upon reasonable request to the corresponding author.
CRediT authorship contribution statement
Ken Chen: Conceptualization, Data curation, Investigation, Writing – original draft, Writing – review & editing. Zuozhen Tian: Conceptualization, Data curation, Writing – original draft, Writing – review & editing. Motomi Enomoto-Iwamoto: Investigation, Writing – original draft, Writing – review & editing. Yejia Zhang: Conceptualization, Writing – original draft, Writing – review & editing.
Conflict of interests
There is no financial or non-financial conflicts of interest to report by the authors.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Ruel N.Markova D.Z.Adams S.L.Fibronectin fragments and the cleaving enzyme ADAM-8 in the degenerative human intervertebral disc Spine 392014127412792501001310.1097/BRS.0000000000000397 PMC 4229950 · doi ↗ · pubmed ↗
- 2Zack M.D.Melton M.A.Stock J.L.Reduced incidence and severity of experimental autoimmune arthritis in mice expressing catalytically inactive A disintegrin and metalloproteinase 8 (ADAM 8)Clin Exp Immunol 15820092462561973713910.1111/j.1365-2249.2009.04009.x PMC 2768814 · doi ↗ · pubmed ↗
- 3Tian Z.Shofer F.S.Fan M.ADAM 8 inactivation retards intervertebral disc degeneration in mice Genes Dis 11320231010593829219410.1016/j.gendis.2023.06.028PMC 10825293 · doi ↗ · pubmed ↗
- 4Zhang Y.Tian Z.Gerard D.Elevated inflammatory gene expression in intervertebral disc tissues in mice with ADAM 8 inactivated Sci Rep 11202118043346910110.1038/s 41598-021-81495-y PMC 7815795 · doi ↗ · pubmed ↗
- 5Zhang C.Zhong L.Lau Y.K.Single cell RNA sequencing reveals emergent notochord-derived cell subpopulations in the postnatal nucleus pulposus FASEB J 382024 e 2336310.1096/fj.202301217 RPMC 1075756438085183 · doi ↗ · pubmed ↗