Exposure to copper metal enhances the tolerance of An. gambiae s.s. over multiple generations while reducing both fertility and fecundity in this primary malaria vector
Massioudou Koto Yérima Gounou Boukari, Genevieve Tchigossou, Innocent Djègbè, Ghislain T. Tepa-Yotto, Eric Tossou, Donald Hessou-Djossou, Camille Dossou, Louckman Monra Seidou, Aldo Emmanuel C. Glokpon, Danahé Adanzounon, Adam Gbankoto, Rousseau Djouaka, Isra Wahid

TL;DR
Exposure to copper reduces mosquito survival and reproduction over generations, but may help control malaria by lowering their fitness.
Contribution
This study shows copper exposure increases An. gambiae tolerance but reduces fertility and fecundity over multiple generations.
Findings
Copper exposure increases larval mortality initially but decreases over generations.
Copper exposure prolongs pupation time and reduces fecundity and fertility in adult mosquitoes.
Copper exposure lengthens the gonotrophic cycle and lowers egg hatching rates.
Abstract
Anopheles s.l. displays the potential to develop tolerance to heavy metals, particularly copper, this may occur at a significant biological cost, which can adversely affect its ecological fitness. This study investigated the larval metal exposure on larval development and reproduction of An. gambiae s.s., a laboratory susceptible strain, kisumu. Stage 2 larvae of Anopheles gambiae, Kisumu were exposed to C 1 = 484 μg L -1, C 2 = 300 μg L -1 and 0 μg L -1 (control) of copper chloride. Larval mortality, pupation time, pupation rate, gonotrophic cycle length, fecundity and fertility of larvae/adults were assessed over six generations. Results revealed that larval mortality rate was significantly higher in the C 1 groups of each group (p = 0.000), but this mortality rate decreased over generations. Pupation time was extended to 13 and 14 days respectively for C 2 and C 1 groups (p =…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
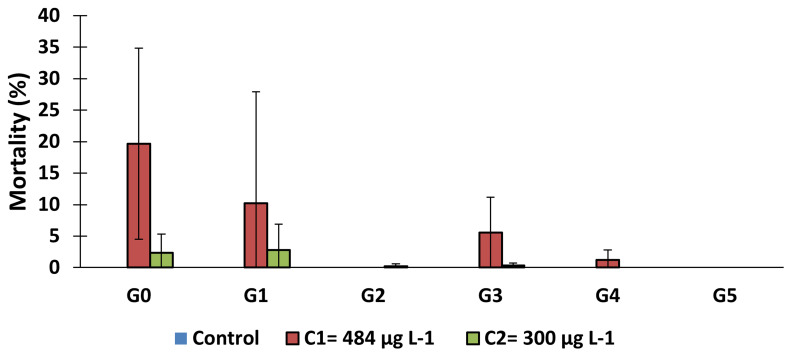 Figure 1
Figure 1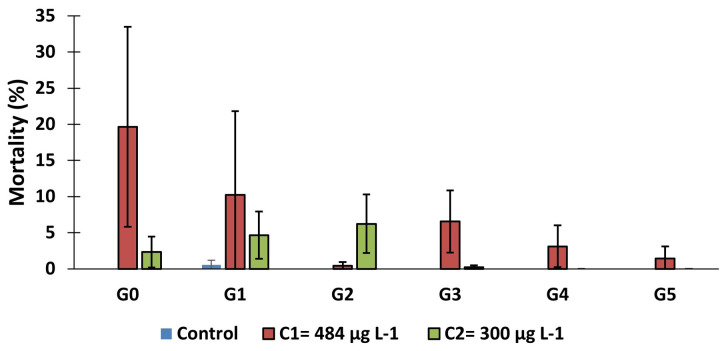 Figure 2
Figure 2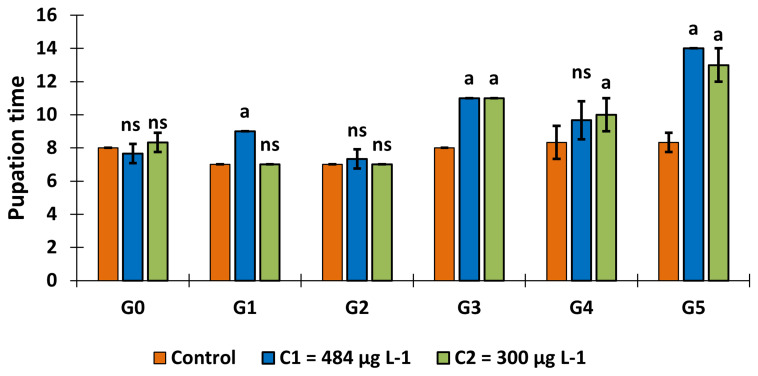 Figure 3
Figure 3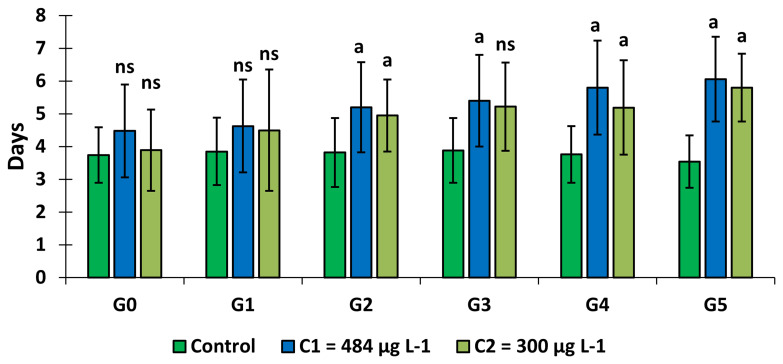 Figure 4
Figure 4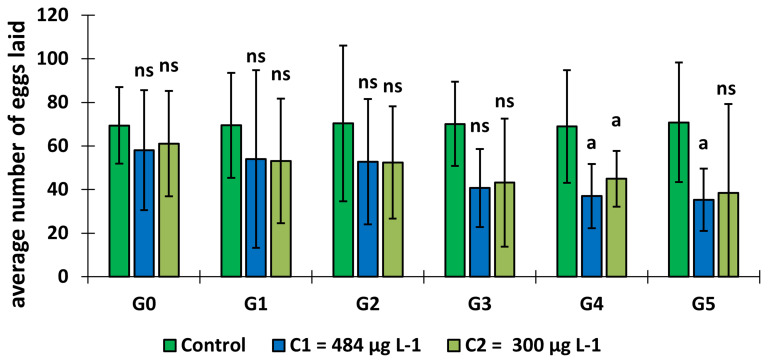 Figure 5
Figure 5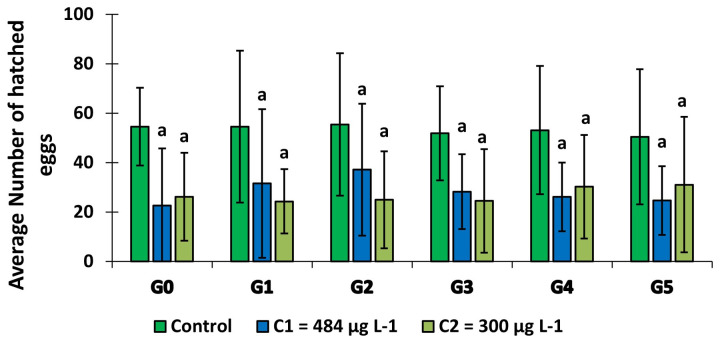 Figure 6
Figure 6| Generations | Control | C 1 = 484 μg L -1 | C 2 = 300 μg L -1 |
|---|---|---|---|
|
| 71 ± 16.70 b | 48.67±7.75 *b | 64.55±12.13 b |
|
| 94.11±34.27 a | 96.33±15.71 a | 93.44±33.54 a |
|
| 90.11±27.29 a | 98.33±23.01 a | 90.11±34.78 a |
|
| 96.56±16.02 a | 91.22±10.27 a | 98.67±13.53 a |
|
| 97.22±9.17 a | 67.56±6.84 *b | 80.22±9.73 b |
|
| 96.22±12.46 a | 23.56±3.02 *c | 80.22±9.05 b |
| Generations | Control | C 1 = 484 μg L -1 | C 2 = 300 μg L -1 |
|---|---|---|---|
|
| 69.70±16.59 b | 67.42±12.42 b | 46.82±11.52 b |
|
| 97.05±37.44 a | 98.04±16.47 a | 84.54±38.59 a |
|
| 93.83±32.19 a | 98.08±23.72 a | 88.04±41.12 a |
|
| 93.67±17.51 a | 93.17±11.79 a | 96.17±14.17 a |
|
| 84.91±10.19 a | 69.08±10.75 *b | 89.47±12.78 a |
|
| 95.73±13.17 a | 61.79±9.36 *b | 66.75±10.54 *a |
- —Wellcome Trust
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsMalaria Research and Control · Insect Pest Control Strategies · Insect Resistance and Genetics
Introduction
The prevalence of mosquito-borne diseases is highly dependent on the ability of larvae to survive in breeding sites and on the ability of adult females to lay eggs in aquatic ecosystems ^ 1, 2 ^. The pollution of aquatic ecosystems due to the widespread and overuse of pesticides in agriculture is a cause for concern in vector control ^ 3 ^. Pesticide residues are therefore easily found in aquatic ecosystems through leaching, erosion and spray drift ^ 4 ^. Human activity has led to a large-scale increase in various environmental pollutants ^ 5, 6 ^. Except agriculture, industry, mining, household and municipal waste are the main sources of environmental pollution containing heavy metal ^ 7, 8 ^. Demographic growth in Africa, especially in urban areas, is increasing the load of these pollutants ^ 9 ^.
Copper is a metal naturally present in the environment through volcanic eruptions, plant decomposition, forest fires, marine aerosols and soil dust ^ 10 ^. This metal can easily find its way into the environment through discharges of industrial water, incineration of household waste, etc. and especially in the manufacture of agricultural fertilizers and pesticide (fungicides or algicides). According to Kabata-Pendias ^ 10 ^, copper is much more concentrated in light silty or clayey soils. In these small ecosystems, larvae of An. gambiae s.l., the main vector of malaria in Africa, are easily found. The presence of this vector in these polluted ecosystems represents a major change because it was known to shelter clear, sunny, unpolluted water ^ 11, 12 ^. According to these authors, the urbanization of cities in sub-Saharan Africa has enabled these vectors to adapt to xenobiotics.
Over the last decade, studies have shown the negative impact of pesticides on the immature stage (larvae and pupae) and on the reproductive capacity of adult mosquitoes. These studies showed that atrazine and glyphosate influenced pupal emergence rate, sex ratio and mosquito size in Aedes aegypti and Aedes albopictus ^ 3 ^ as glyphosate and cypermethrin reduced the emergence rate of An arabiensis and Culex quinquefasciatus ^ 13 ^. Exposure of Aedes aegypti larvae to Roundup also reduced adults survival and their hatching time ^ 14 ^. Resistance of An. gambiae to deltamethrin following exposure of the larval stage to pesticides has recently been demonstrated ^ 15 ^.
Others studies have shown that non-pesticidal chemical residues modulate the capacity of detoxification enzymes ^ 16– 18 ^ as well as phenotypic resistance to insecticides ^ 19 ^. This regulation requires biological costs on the different developmental stages of the malaria vector and consequently on malaria transmission ^ 13, 16, 20 ^. Indeed, Copper plays an important role in insect immunity and in the regulation of proteins involved in melanization ^ 21, 22 ^. In Ae. aegypti and An. gambiae s.l., eggshell formation involves prophenoloxidas and lac-cases ^ 23, 24 ^ which are all copper-dependent metalloproteins ^ 25 ^.
Recently , studies revealed that water, soil and even market garden produces are contaminated with lead, cadmium, chromium, zinc and copper in Benin ^ 26, 27 ^. In the absence of an effective management policy for solid and liquid, domestic and industrial waste, this heavy metal pollution of the various compartments of the biosphere cannot be reduced. Exposure of An. gambiae larvae to copper confers on its adult stages resistance to pyrethroids ^ 20 ^, insecticides mainly used in public health. However, vector transmission capacity depends on the insect's ability to survive and reproduce. In Benin, very little work has been done on An. gambiae s.s. life history vis a vis to metal pollution. This study aimed to understand the effects of copper on life traits of An. gambiae s.s., the major malaria vector in Benin.
Methods
Metal
Copper was used in the form of copper chloride dihydrate, CuCl2, 2H2O; purity 99.0% (Park Scientific Limited, 24 Low Farm Place, Moulton Park, Northampton, NN3 6HY, UK, Catalogue No. C11761). From this product, a stock solution of 1g L ^-1^ was prepared in a 35 mL bottle. Thus, 30.30 mg of copper chloride was dissolved in 30 mL of ultrapure water.The bottle was wrapped in aluminum foil then stored at 4°C for subsequent uses.
Mosquito breeding
Susceptible strain of An. gambiae s.s., kisumu was reared in the insectary of the AgroEcoHealth platform of the International Institut of Tropical Agriculture (IITA-Benin). Mosquitoes (from eggs to adult stage) were reared under laboratory conditions (temperature 27°± 1°C, relative humidity 70–80% and a photoperiod of (12L: 12D). The larvae were fed with Tetramin baby (Tetra GmbH, Made in Germany, Herrenteich, Distributed in UK by Spectrum Brands, Manchester, Catalogue No. 879917) while the adults were fed with 10% sucrose (10g of local sugar diluted in 100 ml of distilled water) solution.
Effect of copper on mortality of larvae, pupae and adults of
An. gambiae s.s. kisumu
Anopheles gambiae s.s. larvae ( kisumu strain) were exposed to two different concentrations of copper chloride, the Lethal Dose (LD 25) (C 1 = 484 μg L ^-1^) and a lower concentration (C 2 = 300 μg L ^-1^) than this LD 25. Nine (9) bowls of 4L capacity were placed on the shelf. Distilled water was used as control group with no presence of copper. Each concentration was repeated three time and bowls were filled with a final volume of two (2) liters. 2 ^nd^ instar larvae (Fo) were transferred to each bowl according to modified methods of Miah ^ 28 ^ who used third instar larvae. Furthermore, these larvae were exposed to these concentrations of copper throughout their larval stage. They were fed daily with Tetramin baby (Tetra GmbH, Made in Germany, Herrenteich, Distributed in UK by Spectrum Brands, Manchester). They were kept in insectary at 27±1˚C under 12:12 light: dark regime with 70–80% relative humidity and mortalities were recorded every morning. Pupae were counted daily in small cup and transferred to their respective cages covered with mosquito nets. Adults (G0) were fed with 10% sugar solution. After emergence, the larval mortality rate, mean of pupae recorded according to concentration were calculated in the control group as well as in the test groups. The pupation time was also determined as the day on which the maximum number of pupae was obtained ^ 5 ^. Thus, the adults obtained from each group were separated into two batches.
The first batch of adult mosquitoes (G0) was used to obtain the next generation (G1). Thus, the females (G0) of each group (control, Concentration 1 and Concentration 2) were blood fed and allow to lay eggs in the petri dishes (Fisherbrand, USA) containing cotton soaked in distilled water and surmounted by a filter paper (Wattman 3 mm/1 cm diameter). The eggs (G1) were placed in small bowls (250 ml) containing distilled water. After egg hatching, the larvae (G1) were transferred to large bowls according to the hatching rate of each group. These larvae (G1) were maintained until the 2 ^nd^ instar. At this stage, they were exposed to copper according to the procedure previously described at the same concentrations (484 μg L ^-1^, 300 μg L ^-1^). The monitoring remained the same until the adults were obtained (G1) and the cycle was repeated till generation 5.
Determination of the gonotrophic cycle
In order to assess the effect of copper on reproduction, three parameters were recorded: the gonotrophic cycle, fecundity and fertility. The larvae of An. gambiae, kisumu were exposed to 484 μg L ^-1^ (C 1) and 300 μg L ^-1^ (C 2) as described previously. After emergence, 20 females and 40 males day-old mosquitoes from each treatment (concentration) were transferred to new cages ^ 20 ^. Each treatment was repeated three times, as well as the control. On the 3 ^rd^ day after emergence, the females were fasted for 6 hours and then blood-fed successively for three days ^ 29 ^. 48 h after the last blood meal, the 20 females in each cage were then introduced individually and gently into small cups containing cotton soaked in water and surmounted by a filter paper (Wattman 3 mm/1 cm diameter). The cups were covered with mosquito net with cotton soaked in 10% sucrose was placed on the top of the net. Each cup was checked daily to identify females of An. gambiae s.s. that have laid eggs and mosquitoes were gently removed from the cup and transferred to a new eppendorf tubes containing cotton and silica gel and stored at -20°C for subsequent experiments. The time taken for the females to lay eggs was determined.
Fecundity and fertility
To determine the fecundity and fertility, the number of mosquitoes that laid eggs and the number of eggs laid (fecundity) in each cup was counted under a binocular loupe (Wild 445111 Heerbrugg, Switzerland) every 24h ^ 29 ^.
The eggs from each mosquito were placed in 250 ml cup containing distilled water. The number of larvae hatched was counted every 24 hours for 96 hours to determine fertility. The experiment was carried out over 6 generations.
Statistical analysis
All data collected were registered in Microsoft Excel 2016 software. The Kolmogorov-Smirnov test was used to assess the normality of the distribution. The chi-square test was used to compare larval mortality rates. General Linear Model (GLM) permitted to determine the difference between the different concentrations, between generations and parameters (Pupation time, fecundity, fertility, larvae, pupae) meanwhile, the Tukey HSD test was used to identify possible differences.
For all analyses, the difference was considered significant at the 5% threshold.
Results
The effect of larval exposure to copper on larval mortality
The mortality rate of An. gambiae s.s., larvae after 3 days' exposure to copper varied according to concentrations ( Figure 1). For the same generation, the mortality rate increased according to the concentration. Excepted for generation 2 and the last generation (G5), mortality rate in test group (C 1) increased significantly in all the generations compared to control group (generation 0: *X ^2^
- = 305.79, df = 2, p = 0.000; generation 1: *X ^2^
- = 121.39, df = 2, p = 0.000; generation 3: *X ^2^
- = 94.28, df = 2, p = 0.000; generation 4: *X ^2^
- = 22.09, df = 2, p = 0.0002). However, this rate decreased over the generations for C 1 and C 2 concentrations.
Mortality rate of larvae after 3 days of exposure to 484 μg L -1 and 300 μg L -1 copper.‘G’ means Generation. bars represent standard deviation.
The mortality rate of An. gambiae s.s., larvae after 7 days' exposure to copper was calculated ( Figure 2). This mortality rate decreased from 19.67% in generation 0 to 1.44% in generation 5 for C 1 while it was 2.33% and 0 in C 2. In the first generation (G0), mortality rate in C 1 group was significantly higher compared to control group ( *X ^2^
- = 305.79, df = 2, p = 0.000). Similar results were observed in generation 1 ( *X ^2^
- = 78.16, df = 2, 0.000), generation 2 ( *X ^2^
- = 99.82, df = 2, 0.000), generation 3 ( *X ^2^
- = 109.39, df = 2, p = 0.000), generation 4 ( *X ^2^
- = 56.58, df = 2, p = 0.000) and generation 5 ( *X ^2^
- = 16.05, df = 2, p = 0.0003) compared to control.
Mortality rate of larvae after 7 days of exposure to 484 μg L -1 and 300 μg L -1 copper.‘G’ means generation. bars represent standard deviation.
The effect of larval exposure to copper on pupation time
In the control group, the average pupation time was 7.78±0.65 days over the 6 generations (GLM: F = 16.55, df = 10, p = 0.000) ( Figure 3). In generation 1, pupation time increased in C 1 = 484 μg L ^-1^ group (9.00 ±0.00, p = 0.005, Tukey test). This mean pupation time increased significantly in generation 3, with a fair value of 11.00±0.00 days in C 1 and C 2 (Control and both concentrations: p = 0.00017, Tukey test) and 10.00±1.00 days in C 2 in generation 4 (p = 0.037, Tukey test). Finally, pupation time in generation 5 reached 14.00±0.00 days in C 1 and 13.00±1.00 days in C 2 (p = 0.00016, Tukey test).
Variation in pupation time over generations following exposure of An. gambiae s.s. larvae to copper at C 1 = 484 μg L -1 and C 2 = 300 μg L -1.‘G’ means Generation. ‘ns’ means “no significant difference with the control of the same generation”. ‘a’ means “significant difference with the control of the same generation” and ‘bars’ represent standard deviation.
It should also be noted that this pupation time increased over the generations in the larvae exposed to copper (( F = 98.31, df = 5, p = 0.00), with a significant difference observed from generation 3 to the last generation.
Effect of larval exposure to copper on pupation rates
The pupation rate was significantly lower in females only in (C 1) group (GLM: F = 17.68; df = 10, p = 0.000), particulary in the generations 0, 4 and 5 (C 1) compared to control group ( Table 1).
Effect of exposure of
An. gambiae s.s., to copper on emergence rates
Exposure of An. gambiae s.s. larvae to copper (C 1) significantly reduced pupal emergence rates (GLM: F = 14.66, df = 10, p = 0.000) in generations 4 and 5 while exposure to copper (C 2) reduced emergence rate in generation 5 only. The results for emergence rates were summarized in Table 2.
Effect of copper on the gonotrophic cycle of
An. gambiae s.s.
The number of days taken by adult females to lay eggs after the last blood feeding was determined. For the two concentrations C 1 = 484 μg L ^-1^ and C 2 = 300 μg L ^-1^, mosquitoes took longer (4.48 days to 6.06 days in C 1 and 3.89 days to 5.80 in C 2 respectively from G0 to G5) to lay eggs than the control group (3.55 days to 3.88 days) ( Figure 4). This time increased slightly over generations in the test groups (GLM: F = 2.84, df = 9, p = 0.003). It was significantly higher in C 1 and C 2 (Tukey test, p = 0.009 and p = 0.002, respectively) in generation 2 and C 1 (p = 0.003; Tukey test) in generation 3. The gonotrophic cycle was also higher in C 2 (p = 0. 008; Tukey test), C 1 (p= 0.016; Tukey test) in generation 4 and finally in C 1 (p = 0.000) and C 2 (p = 0.000; Tukey test) in the last generation (G5) compared to control.
Changes in adult fecundity after exposure of the larval stage to copper.‘G’ means Generation. ‘ns’ means “no significant difference with the control of the same generation”. ‘a’ means “significant difference with the control of the same generation” and ‘bars’ represent standard deviation.
Between treatments (C 1 and C 2), no significant difference (p = 0.10, Tukey test) was observed within the same generation. Larval exposure to 300 μg L ^-1^ of copper significantly increased the gonotrophic cycle over the generations ( F = 6.52, df = 4, p = 0.003), from generation 0 to generation 5. A significant increase was observed only between generation 0 and generation 5 (GLM: F = 6.59, df = 4, p = 0.004, Tukey test) on larvae exposed to 484 μg L ^-1^.
Effect of copper on the fecundity of adult females
An. gambiae ss.
Mosquito larval exposure to copper had an effect on the fecundity of adults An. gambiae s.s. ( Figure 5). The average number of eggs laid per female decreased over the generations in the test groups and remained lower than in the control group (GLM: F = 1.87, df = 9, p = 0.05). Indeed, it was significantly lower in C 1 (p = 0.00, Tukey test), C 2 (p = 0.014, Tukey test) at generation 4 and C 1 at generation 5 (p = 0.00, Tukey test).
Changes in adult fecundity after exposure of the larval stage to copper.‘G’ means Generation. ‘ns’ means “no significant difference with the control of the same generation”. ‘a’ means “significant difference with the control of the same generation” and ‘bars’ represent standard deviation.
Effect of larval exposure to copper on the adult fertility
The average number of eggs hatched in the groups which larvae were exposed to copper was much lower than in the control group ( Figure 6). In larvae exposed to 300 μg L ^-1^, this egg fertility was roughly equal over the first 4 generations (G0, G1, G2 and G3) before rising slightly in the last two generations. In contrast, in C _1 _= 484 μg L ^-1^ group, fertility increased from G0 to G2 and decreased from G2 to G5. For each generation, the fertility rate was significantly lower (GLM: F = 84,673, df = 2, p = 0.000) in the groups exposed to copper compared to the control group. There was no significant difference between generations for the same concentration (p = 0,539).
Average number of eggs hatched per female over the generations. G’ means Generation.‘ns’ means “no significant difference with the control of the same generation”. ‘a’ means “significant difference with the control of the same generation” and ‘bars’ represent standard deviation.
Discussion
This study examined how frequent copper exposure affects the growth and reproductive capacity of An. gambiae s.s. The findings indicated that copper exposure led to increased mortality, extended pupation times, and negatively impacted the pupation rates of the larvae. Previous studies showed that copper adversely affects mosquito mortality and normal pupation durations ^ 28, 30, 31 ^. Copper exposure has been linked to a decrease in the number of eggs produced by female mosquitoes. For example, when larvae were raised in environments with copper pollutants, the resulting adults had significantly lower fecundity, which directly impacts population dynamics and vector control efforts ^ 20 ^. In this study, the observed larval mortality following the copper exposure highlights its toxicity, which disrupts various organ functions and vital processes within the insects. Copper exposure can alter the intestinal microbiota of larvae and pupae, which is crucial for digestion, nutrient absorption, and overall growth ^ 30 ^. This disruption likely contributes to the increased larval mortality seen in the experimental groups, as it results in significant energy loss necessary for survival. Additionally, copper presence in the environment induces stress in larvae, modifying their physiological responses, as noted in Aedes aegypti ^ 31– 33 ^. This stress reduces lipid reserves during the transition to adulthood, diminishes body mass, and enhances starvation tolerance, effects that can extend to subsequent generations ^ 31 ^. Such adaptations to hunger in offspring may explain the reduction in larval mortality across generations observed in this study. Mosquito larvae appear to develop mechanisms to cope with heavy metal exposure like copper ^ 34 ^. This adaptation may facilitate the expansion of these vectors into polluted habitats, leading to heightened insecticide resistance ^ 5, 35 ^. Results from this study revealed that copper exposure significantly impacts the pupation process in mosquitoes, particularly affecting the timing and success of this critical developmental stage as reported by Miranda and colleagues, showing the detrimental effects of copper on both pupation and emergence rates ^ 36 ^. The shift from stage 4 to the pupal stage, as well as from the pupal stage to adulthood, demands considerable energy reserves accumulated during the larval phase, since pupae do not feed. Copper disrupts the intestinal microbiota in larvae, leading to intestinal dysfunction due to altered microbiota, which hampers nutrient absorption and diminishes energy reserves necessary for molting, metamorphosis, and adult development ^ 37, 38 ^ as reported by numerous studies ^ 5, 39, 40 ^. Moreover, insects are characterized by their high reproductive capacity ^ 41 ^, hence, female mosquitoes require iron, obtained from blood meals, to lay eggs, but the eggs must also be viable for successful hatching ^ 42 ^. Copper negatively impacts the reproductive cells of mosquitoes, inducing apoptosis and adversely affecting egg viability ^ 20, 43 ^. In both Aedes aegypti and An. gambiae s.l., eggshell formation relies on copper-dependent metalloproteins, such as prophenoloxidases (PPO) and laccases (lacs) ^ 23– 25 ^. Additionally, copper exposure in mosquito larvae heightens oxidative stress, which detrimentally affects adult fecundity ^ 31 ^. The adverse effects on fertility and fecundity may stem from malformations in the midgut during post-embryonic development, impairing blood digestion. Females need a protein-rich blood meal to initiate vitellogenesis and produce eggs ^ 44 ^. Consequently, inadequate nutrient absorption or difficulties in digesting blood meals can result in non-viable eggs, thereby reducing fecundity and hatching rates ^ 45, 46 ^. Also, the differential expression of specific proteins, such as mucins, may account for the tolerance of Anopheles species to copper ^ 47 ^. Metal exposure can disrupt protein synthesis in mosquitoes, leading to the blockage of certain proteins and the overproduction of others ^ 47, 48 ^. Down-regulated proteins can impair various functions, including metabolic, immune, reproductive, and cellular growth processes ^ 48 ^. Conversely, some upregulated proteins might help mitigate the intracellular concentration of harmful substances ^ 47 ^. Copper alters the midgut epithelium of pupae, potentially compromising midgut function and mosquito survival. The gonotrophic cycle, which measures the frequency of blood meals, is crucial for assessing the potential for parasite acquisition and transmission ^ 49 ^. Extending the gonotrophic cycle is significant for vector control, as mosquitoes are known for their prolific reproduction ^ 25 ^. Mosquitoes can take multiple blood meals, resulting in numerous egg-laying events. A prolonged gonotrophic cycle is favorable for vector control, as it decreases the reproductive rate of these vectors, leading to a lower human bite rate and reduced malaria transmission ^ 29 ^. Overall, copper exposure adversely affects the pupation process in mosquitoes by prolonging pupation time and decreasing adult emergence rates. These findings underscore the potential of copper as a tool in mosquito control strategies, particularly when considering its effects on larval development and subsequent life stages.
Conclusion
Exposure of An. gambiae, kisumu susceptible strain to copper over multiple generations led to a decrease in larval mortality, indicating that the larvae adapted to this metal. However, significant reductions were observed in pupation time, pupation rates, and the number of adults in the later generations, suggesting that this adaptation came at a cost to certain essential functions of the insect. The extended gonotrophic cycle, along with decreased fecundity and fertility, highlights the significant impact of copper on the reproductive capabilities of An. gambiae s.s. While previous studies have noted that copper can promote resistance to insecticides, it is important to recognize its potential benefits in vector control due to its adverse effects on the species' reproductive functions, which may help limit the spread of malaria vectors.
Ethics and consent
Ethical approval and consent were not required
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Alto BW Lounibos LP Higgs S : Larval competition differentially affects arbovirus infection in Aedes mosquitoes. Ecology. 2005;86(12):3279–88. 10.1890/05-0209 19096729 PMC 2605070 · doi ↗ · pubmed ↗
- 2Reiskind MH Lounibos LP : Effects of intraspecific larval competition on adult longevity in the mosquitoes Aedes aegypti and Aedes albopictus. Med Vet Entomol. 2009;23(1):62–8. 10.1111/j.1365-2915.2008.00782.x 19239615 PMC 2651082 · doi ↗ · pubmed ↗
- 3Bara JJ Montgomery A Muturi EJ : Sublethal effects of atrazine and glyphosate on life history traits of Aedes aegypti and Aedes albopictus (Diptera: Culicidae). Parasitol Res. 2014;113(8):2879–86. 10.1007/s 00436-014-3949-y 24853538 · doi ↗ · pubmed ↗
- 4Schulz R : Field studies on exposure, effects, and risk mitigation of aquatic nonpoint-source insecticide pollution: a review. J Environ Qual. 2004;33(2):419–48. 10.2134/jeq 2004.4190 15074794 · doi ↗ · pubmed ↗
- 5Oliver SV Brooke BD : The effect of metal pollution on the life history and insecticide resistance phenotype of the major malaria vector Anopheles arabiensis (Diptera: Culicidae). P Lo S One. 2018;13(2): e 0192551. 10.1371/journal.pone.0192551 29408922 PMC 5800662 · doi ↗ · pubmed ↗
- 6Vareda JP Valente AJM Durães L : Assessment of heavy metal pollution from anthropogenic activities and remediation strategies: a review. J Environ Manage. 2019;246:101–18. 10.1016/j.jenvman.2019.05.126 31176176 · doi ↗ · pubmed ↗
- 7Eisler R Wiemeyer SN : Cyanide hazards to plants and animals from gold mining and related water issues.In: Ware GW, editor. Reviews of Environmental Contamination and Toxicology.New York, NY: Springer;2004;183:21–54. 10.1007/978-1-4419-9100-3_2 15369321 · doi ↗ · pubmed ↗
- 8Lekfeldt JDS Holm PE Kjærgaard C : Heavy metal leaching as affected by long-time organic waste fertilizer application. J Environ Qual. 2017;46(4):871–8. 10.2134/jeq 2016.11.0458 28783778 · doi ↗ · pubmed ↗