In silico analysis of molecular mimicry between human aquaporin 3, Aspergillus fumigatus aquaporin and aquaporins from allergic sources
Andrés Sánchez, Yaquelin Padilla, Adriana Lorduy, Jorge Sanchez, Marlon Munera, Claudia Baena, Carlos Bernal, Juan Urrego, Elizabeth Garcia, Celso Eduardo Olivier

TL;DR
This study explores how human and fungal aquaporins might trigger allergic reactions through molecular mimicry, potentially worsening atopic dermatitis.
Contribution
The paper identifies conserved antigenic regions and potential epitopes in aquaporins that suggest molecular mimicry between human and fungal sources.
Findings
Aquaporins from human and allergenic sources share conserved antigenic regions with up to 71.4% local identity.
Human AQP3 and A. fumigatus aquaporins share similar linear and conformational epitopes, suggesting molecular mimicry.
Aquaporins were grouped into five clades, with mammalian aquaporins showing the highest identity (95%).
Abstract
Atopic dermatitis (AD) is a chronic inflammatory skin condition that has a significant impact on quality of life. The immune response and allergy symptoms in AD are triggered by the recognition of specific allergens by IgE antibodies. Cross-reactivity can lead to auto-IgE responses, potentially worsening AD symptoms. Our research aimed to enhance our understanding of allergenic sources, including A. fumigatus, and their role in AD. We focused on molecular mimicry between human AQP3 and A. fumigatus aquaporin. In our in-silico analysis, we compared the amino acid sequences of human aquaporin 3 (AQP3) and A. fumigatus aquaporin with 25 aquaporins from various allergenic sources, sourced from the UniProt and NCBI databases. Phylogenetic relationship analysis and homology-based modeling were conducted. We identified conserved antigenic regions located within the 3D structures. The global…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
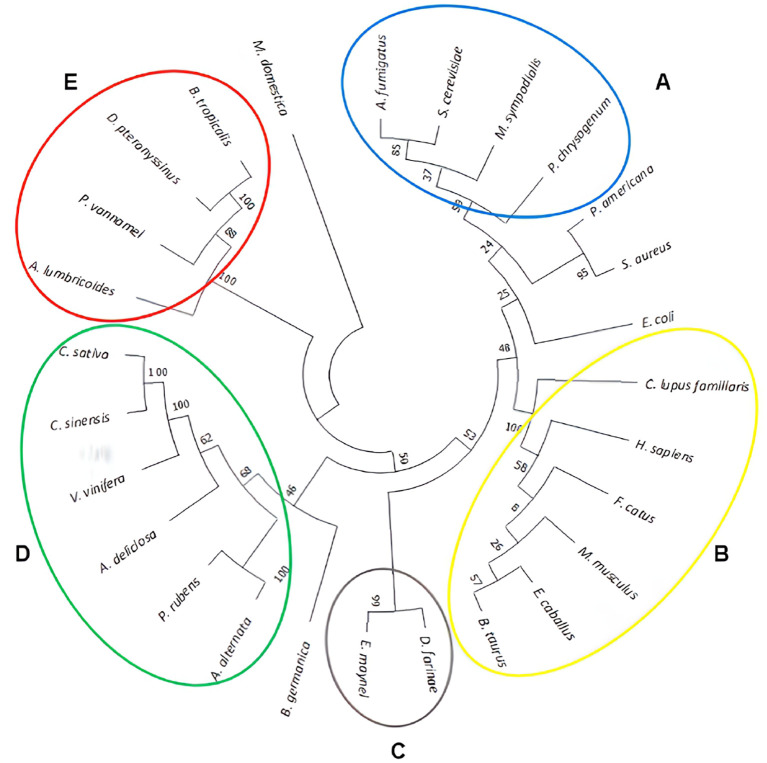 Figure 1
Figure 1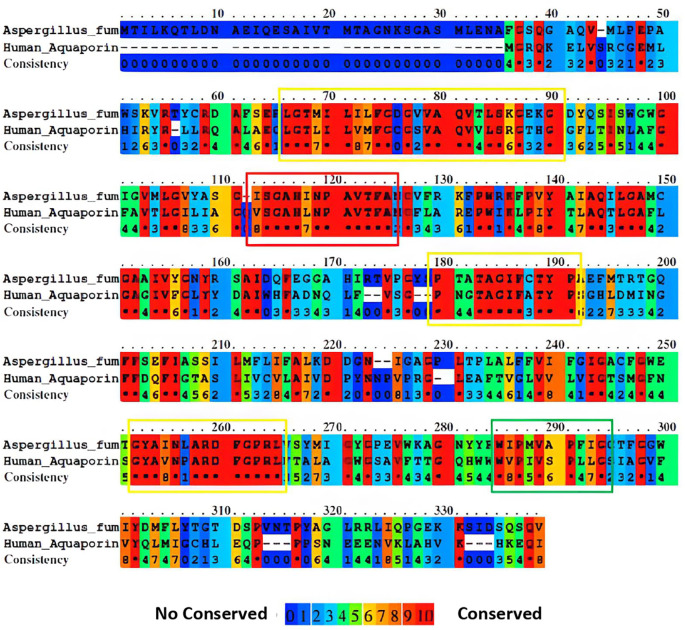 Figure 2
Figure 2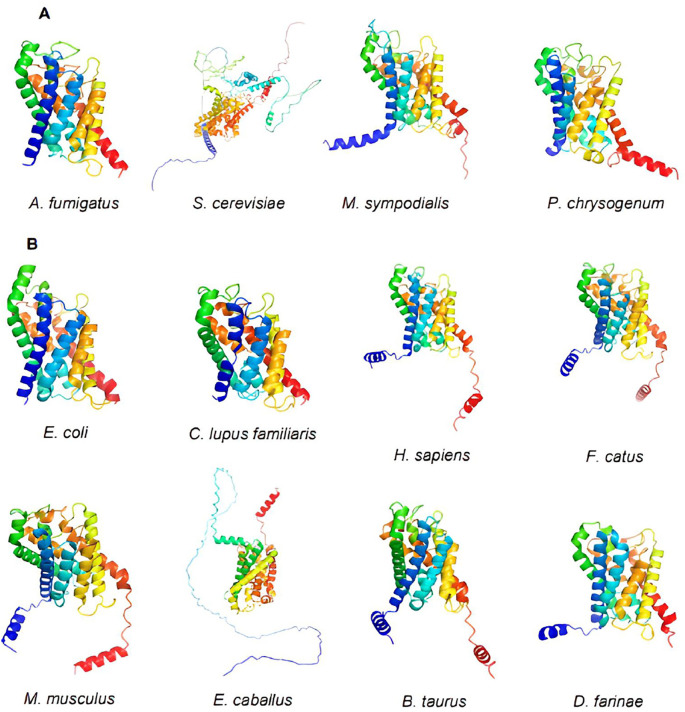 Figure 3
Figure 3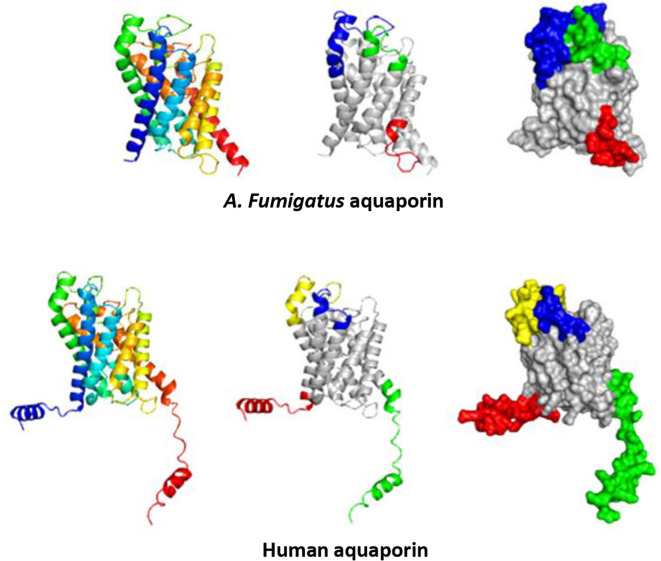 Figure 4
Figure 4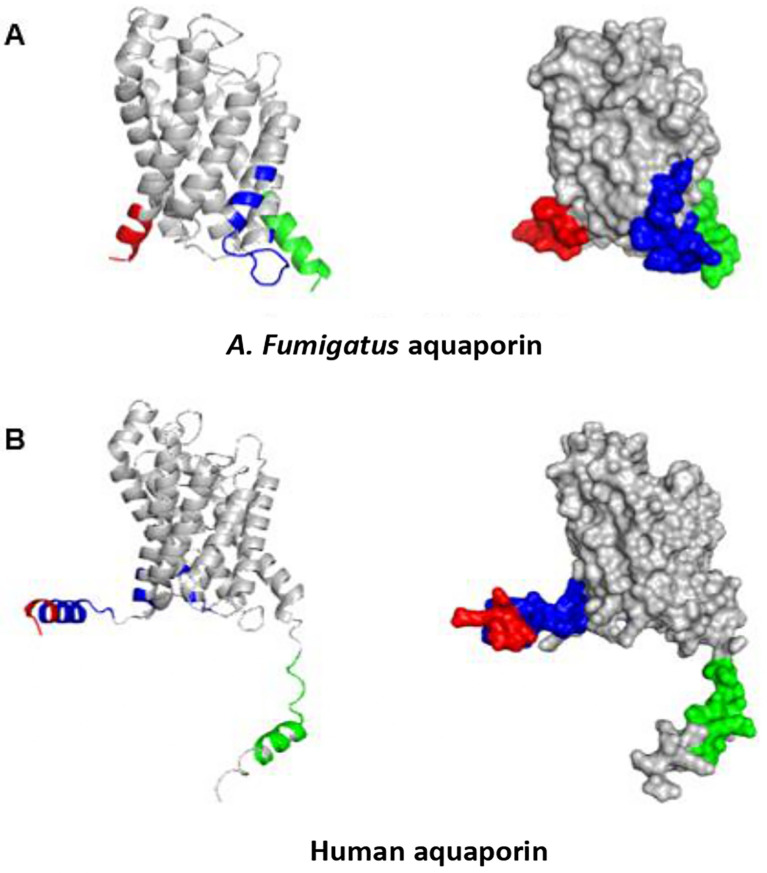 Figure 5
Figure 5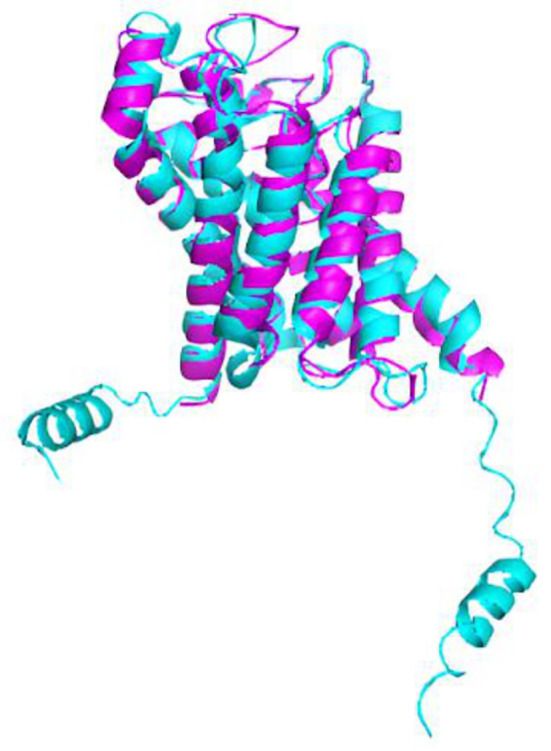 Figure 6
Figure 6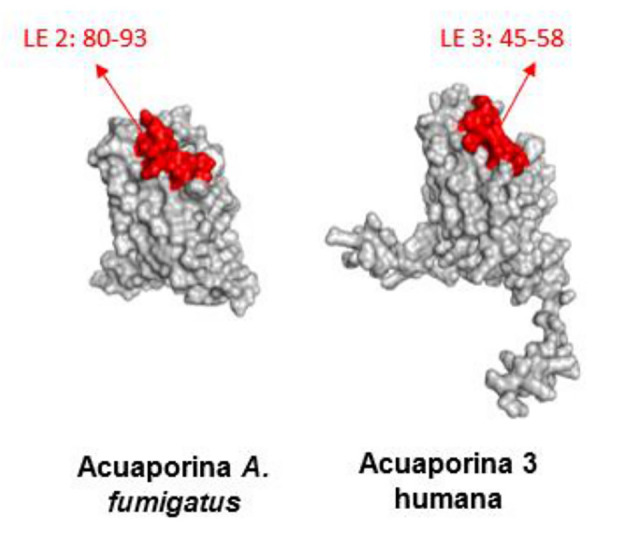 Figure 7
Figure 7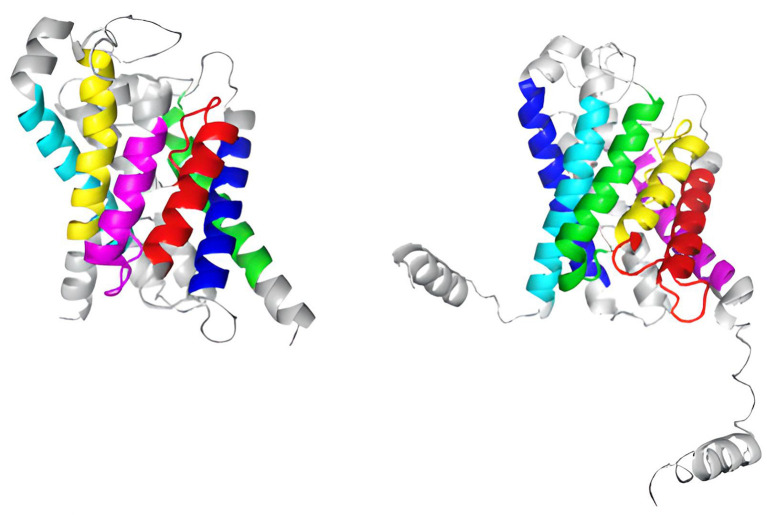 Figure 8
Figure 8| Organisms | Protein | Uniprot/NCBI Entry | Aminoácidos |
|---|---|---|---|
|
| aquaglyceroporin |
| 335 |
|
| Aquaporin-3 |
| 292 |
|
| Aquaporin |
| 265 |
|
| Aquaporin |
| 259 |
|
| Aquaporin |
| 256 |
|
| Aquaporin-like protein |
| 289 |
|
| Aquaporin |
| 144 |
|
| Aquaporin TIP4-1 |
| 196 |
|
| Aquaporin PIP2-1 |
| 283 |
|
| Putative aquaporin TIP13 |
| 251 |
|
| Uncharacterized protein |
| 250 |
|
| Aquaporin TIP3-1 |
| 200 |
|
| Putative glycerol diffusion channel |
| 247 |
|
| Aquaporin |
| 308 |
|
| Aquaporin-3 |
| 292 |
|
| Aquaporin-3 |
| 292 |
|
| Aquaporin-3 |
| 419 |
|
| Aquaporin-3 |
| 292 |
|
| Aquaporin-3 |
| 292 |
|
| Aquaporin |
| 408 |
|
| Putative channel-like protein |
| 291 |
|
| Aquaglycerol porin AQY3 |
| 646 |
|
| Aquaporin-3 |
| 279 |
|
| Pc12g14590 protein |
| 306 |
|
| Glycerol uptake facilitator protein |
| 281 |
|
| Aquaporin |
| 267 |
|
| Glycerol uptake facilitator protein |
| 247 |
| Sources | AQP3 |
| ||||
|---|---|---|---|---|---|---|
| Identity (%) | Similarity (%) | Score | Identity (%) | Similarity (%) | Score | |
|
| 32,6 | 47,5 | 492,5 | - | - | - |
|
| 18,1 | 32,2 | 42,0 | 15,50 | 25,5 | 41 |
|
| 31,5 | 48,7 | 386,0 | 22,90 | 36 | 291 |
|
| 19,6 | 32,7 | 44,5 | 13,70 | 20 | 29 |
|
| 29 | 46 | 360 | 21,80 | 34,6 | 252,5 |
|
| 1,7 | 3,2 | 14,5 | 0,20 | 0,9 | 3 |
|
| 1,7 | 30,5 | 175 | 18,20 | 27,7 | 149,5 |
|
| 21,2 | 32,9 | 218,5 | 24,30 | 38 | 247 |
|
| 25,4 | 38,4 | 193,5 | 18,80 | 29,6 | 167 |
|
| 24,6 | 39 | 190,5 | 19,50 | 34,7 | 167 |
|
| 22,6 | 33,4 | 141,0 | 15,90 | 23,2 | 84,5 |
|
| 27,1 | 44,4 | 332 | 24,20 | 34 | 330 |
|
| 14,9 | 28,5 | 80,5 | 13,00 | 20,2 | 27 |
|
| 72,9 | 75,9 | 1294 | 33,90 | 50,3 | 468,5 |
|
| 95,5 | 97,6 | 1484 | 33,10 | 48,4 | 508,5 |
|
| 66,6 | 68,7 | 1497 | 27,90 | 42,7 | 498 |
|
| 94,9 | 98,3 | 1486 | 32,80 | 48,1 | 497,5 |
|
| 95,5 | 98,6 | 1493 | 33,10 | 48,1 | 506,5 |
|
| 15,6 | 25,4 | 214 | 14,50 | 22,6 | 205 |
|
| 33,1 | 49,5 | 401,5 | 35,90 | 49,6 | 516 |
|
| 15,8 | 25,1 | 516 | 22,60 | 31,4 | 668,5 |
|
| 33,2 | 50,9 | 377,5 | 3,50 | 43,4 | 477 |
|
| 24,6 | 35,2 | 205 | 20,90 | 32,8 | 144,5 |
|
| 35,6 | 53,2 | 475,5 | 31,70 | 45,2 | 436,5 |
|
| 9,9 | 16,1 | 30,5 | 10,40 | 16,7 | 17,5 |
|
| 31,7 | 43,6 | 377,5 | 27,70 | 37,4 | 337,5 |
|
| ||||||
|---|---|---|---|---|---|---|
| Lineal Epitopes | ||||||
| No. | Chain | Start | Final | Peptide | N. Residues | Score |
| 1 | A | 218 | 231 | DDGNIGAGPLTPLA | 14 | 0,763 |
| 2 | A | 80 | 93 | QVTLSKGEKGDYQS | 13 | 0,7 |
| 3 | A | 151 | 180 | AIVYGNYRSAIDQFEGGAHIRTVPGYSPTA | 30 | 0,7 |
|
| ||||||
| 1 | A | 54 | 61 | A:R54, A:T55, A:Y56, A:C57, A:R58, A:D59, A:A60, A:F61 | 8 | 0,869 |
| 2 | A | 211 | 306 | A:F211, A:F293, A:G295, A:W296, A:I297, A:Y298, A:D299, A:M300, A:F301, A:L302, A:Y303, A:T304, A:G305, A:T306 | 14 | 0,815 |
| 3 | A | 215 | 234 | A:A215, A:D218, A:D219, A:G220, A:N221, A:I222, A:G223, A:A224, A:G225, A:P226, A:L227, A:T228, A:L230, A:A231, A:F234 | 15 | 0,763 |
|
| ||||||
|
| ||||||
| 1 | A | 1 | 23 | MGRQKELVSRCGEMLHIRYRLLR | 23 | 0,872 |
| 2 | A | 259 | 292 | FVYQLMIGCHLEQPPPSNEEENVKLAHVKHKEQI | 34 | 0,856 |
| 3 | A | 45 | 58 | QVVLSRGTHGGFLT | 14 | 0,709 |
| 4 | A | 120 | 143 | FGLYYDAIWHFADNQLFVSGPNGT | 24 | 0,702 |
|
| ||||||
| 1 | A | 1 | 5 | A:M1, A:G2, A:R3, A:Q4, A:K5 | 5 | 0,957 |
| 2 | A | 271 | 285 | A:Q271, A:P272, A:P273, A:P274, A:S275, A:N276, A:E278, A:E279, A:N280, A:V281, A:K282, A:L283, A:A284, A:H285 | 14 | 0,914 |
| 3 | A | 6 | 103 | A:E6, A:L7, A:V8, A:S9, A:R10, A:C11, A:G12, A:E13, A:M14, A:L15, A:H16, A:I17, A:L21, A:L22, A:R23, A:L26, A:E96, A:P97, A:I99, A:P102, A:I103 | 21 | 0,775 |
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsAsthma and respiratory diseases · Protein purification and stability · Food Allergy and Anaphylaxis Research
Introduction
Atopic dermatitis (AD) is a chronic and recurrent inflammatory skin disorder characterized by symptoms, such as eczema, erythema, persistent itching, and continuous skin damage. Over the past 30 years, the prevalence of AD has shown a notable rise, affecting between 5% and 20% of the child population and approximately 7% of adults in industrialized countries. ^ 1 ^ ^,^ ^ 2 ^ The pathogenesis of AD is not completely clear; however, allergens can activate an intense Th2 response. ^ 3 ^ Allergenic sources can vary depending on the population; for example, house dust mites are the main source of sensitization in tropical countries, whereas in Europe, pollen is the main trigger for allergies. In addition, depending on the environmental conditions, it is possible to be exposed to different species; therefore, epidemiological studies have shown variations in the frequencies of sensitization. ^ 4 ^ The severity of the disease is influenced by several factors, including compromised skin barrier integrity due to reduced levels of filaggrins and ceramides as well as an elevated presence of water channels such as aquaporin, which drives the skin, making the patient more susceptible to atopic dermatitis. ^ 5 ^ In general, allergy management includes allergen avoidance, pharmacotherapy, and immunotherapy, where only the latter has a beneficial effect that lasts for years and allows for a reduction in anti-inflammatory treatment. ^ 6 ^ ^–^ ^ 8 ^
In addition to other environmental factors, the skin microbiome plays a pivotal role in AD development. Aspergillus fumigatus is an opportunistic pathogen in both poikilothermic and homeothermic animals. In humans, this pathogen has been associated with many pathologies such as invasive cutaneous aspergillosis, saprophytic lung colonization, and aspergillomas. In addition, A. fumigatus can induce IgE-mediated allergic diseases including rhinitis, allergic sinusitis, asthma, hypersensitivity pneumonitis, and AD. ^ 9 ^ ^,^ ^ 10 ^ It has been characterized as an important allergenic source, and some allergens, including thioredoxins and cyclophilins, share homology with human proteins and are implicated in the autoreactive response in AD.
Despite the identification of sensitization to extracts from allergenic sources, such as fungi, the recognition of specific IgE sensitization to allergens is necessary to carry out a better diagnosis and immunotherapy. ^ 11 ^ However, this represents a challenge, because most fungal allergens cross-react with other taxonomically distant species, making it difficult to prove the clinical relevance of IgE reactivity against these allergens. In addition, it is important to consider that the evolution of AD varies greatly among patients with this disease and can be triggered by different factors. For example, in A. fumigatus and AD, studies suggest that specific IgE-mediated activation is induced not by multiple allergens and not by a few major allergens, as is the case with asthma and rhinitis. ^ 12 ^ ^,^ ^ 13 ^ Another problem presented by the diagnosis of allergic diseases associated with A. fumigatus is the lack of standardized allergens for preparation of molecular component tests. Therefore, the discovery of new fungal allergens and the identification of the prevalence of these new components may provide an opportunity to learn more about the pathology of atopic dermatitis, improving the diagnosis and treatment of this disease.
In recent years, aquaporin has become a candidate protein for the study of skin and allergic diseases such as AD, given its role in epidermal hydration and the transport of various substances such as glycerol, salts, and exocrine fluids, favoring dry skin and allergen leakage. ^ 14 ^ They have also been associated with various atopic diseases, such as glaucoma, cancer, epilepsy, obesity, epidermal hyperplasia, and neuromyelitis optical, among others. In AD, an increase in the expression of aquaporin 3 has been observed, which may be related to water loss and the severity of the disease, in contrast to healthy individuals who maintain a balanced expression of this protein in the basal, horny, and spinous layers. ^ 5 ^ The consequences of aquaporin 3 (AQP3) overexpression extend beyond cutaneous water retention as it becomes a target for the exacerbated immune response in AD inflammation. This hypothesis suggests that it may be recognized by human autoantibodies, which, due to the persistence of self-protein exposure, contribute to increased inflammation and chronicity of AD.
Additionally, this protein is present in most organisms with a wide variety of isoforms and functions throughout the taxonomic framework. Recently, aquaporin has been identified in highly allergenic species, such as house dust mites. In a study carried out by Mao et al., describing the Dermatophagoides farinae proteome, two-dimensional immunoblotting was performed, which showed binding with IgE antibodies in a band that has the same isoelectric point and molecular weight of the aquaporin family, suggesting its possible role as an allergen. The objective of this study was to perform an in silico analysis to investigate the potential molecular mimicry between human aquaporins and those from A. fumigatus, as well as various allergenic sources.
Methods
For the selection of aquaporins, the Aspergillus fumigatus (KEY82230) and human aquaporin 3 (Q92482) sequences of interest were compared with existing allergenic protein sequences using Allermatch ( https://allermatch.org/), an allergen comparative server that allows allergen sequence alignment; therefore, those with high identity values were chosen. The sequence pool was obtained from the UniProt ( https://www.uniprot.org/) and National Center for Biotechnology Information (NCBI) databases. Given the diversity in amino acid origins and lengths across the molecules under examination, we employed coverage and identity values to gauge the extent of their relationships. The identity and coverage values between the molecules used in this study were determined using the EMBOSS Needle ( https://www.ebi.ac.uk/Tools/psa/emboss_needle/), specialized for performing paired alignments, and the web server from Praline ( http://www.ibi.vu.nl/), which allowed for paired and multiple alignments. When carrying out the alignment, it was considered that the sequences were different and came from different sources; therefore, the alignment parameters were set to utilize the BLOSUM62 matrix, which primarily assesses evolutionary divergence, along with adjustments to the conditional scoring matrix. ^ 15 ^ ^,^ ^ 16 ^
Phylogenetic analysis
To construct the phylogenetic tree, we conducted a study using “Molecular Evolutionary Genetic Analysis” (MEGA) version 11 program. We employed the neighbor-joining (R) reconstruction method, supported by 100 bootstrap repetitions, to ensure the reliability and robustness of the analysis. Evolutionary distances were computed using the Poisson correction method, which uses a comparison matrix to identify sequence similarities. This matrix was created based on all amino acid sequences of the selected allergens, with all empty gaps removed (full deletions). Branch length summation (SBL) was presented as a measure of overall comparison and homologies, providing insights into the number of nodes, their positions, and the formation of the evolutionarily closest sequence “clusters. ^ 15 ^ ^,^ ^ 17 ^
3D models of proteins
Aquaporin models were acquired from the Protein Data Bank and AlphaFold ( Homo sapiens, Felis catus, Equus caballus, Bos taurus, Mus musculus, Malassezia sympodialis, Saccharomyces cerevisiae, Penicillium chrysogenum, Escherichia coli). For A. fumigatus and Cannis lupus familiaris aquaporin 3D structures were modeled by homology using the Swiss Model server ( https://swissmodel.expasy.org/), and these models were refined in ModRefiner. ( https://zhanggroup.org/ModRefiner/), an algorithm for the fine-grained refinement of protein structures at the atomic level. ^ 18 ^
Epitope prediction
Models were used to locate possible epitope regions predicted by the Ellipro server ( http://tools.iedb.org/ellipro/), which was used to predict linear and discontinuous epitopes in the aquaporins of A. fumigatus, (AQP3), Cannis lupus familiaris, Felis catus, Equus caballus, Bos taurus, Mus musculus, Malassezia sympodialis, Saccharomyces cerevisiae, Penicillium chrysogenum and Escherichia coli. The potential epitopes were selected under the criteria of a score ≥ 7, as this value evaluates how exposed the residue is and the binding capacity of the epitope with the paratope. ^ 15 ^ ^,^ ^ 17 ^
Identification of aquaporin domains
The characterization of the protein domains was carried out using the Interpro web server ( https://www.ebi.ac.uk/interpro/); with this bioinformatic tool we obtained those domains with functional activity present in the protein sequence of the aquaporins studied. Subsequently, a review of the tertiary structure was carried out using the PyMOL program, which functioned as a molecular viewer, allowing identification of the location of the protein domains in the tertiary structure of each one.
Results
Aquaporins sequence alignments
A total of 25 aquaporin sequences from different sources were selected and compared with the A. fumigatus aquaporin and human aquaporin ( Table 1), the identity percentages of the aquaporins from different sources Vs A. fumigatus aquaporin and AQP3 are shown in Table 2. Notably, human AQP3 exhibited maximum values of identity and similarity of 95.5% and 98.6%, respectively, with aquaporin allergy sources. When comparing A. fumigatus aquaporins and AQP3, an identity of 32.6% and a similarity of 47.5% were observed, indicating a significant degree of homology between them. Regarding the AQP of A. fumigatus, the highest identity values were observed in the AQP of M. simpodialies (35%), followed by the mammalian species that also had greater identity with human AQP3, such as C. familiaris (33, 9%), Felis gatus (33, 10%), Mus musculus (33, 10%), and Bos taurus (32, 80%). The lowest identity and similarity results were obtained with apple ( Malus domestica), with values of 0.2% and 0.9%, respectively.
Table 2.: Identity and similarity between AQP3, A. fumigatus aquaporin and aquaporins from allergen sources.
Phylogenetic analysis
Phylogenetic relationships were determined using the neighbor-joining method, incorporating all 27 amino acid sequences. The resulting phylogenetic tree is presented in a circular format in Figure 1. Evolutionary distances were calculated using the Poisson correction method and were exprex|ssed as the number of amino acid substitutions per site. The tree visually represents the grouping and arrangement of the analyzed aquaporins, facilitating a comprehensive comparison and revealing their interrelatedness.
Phylogenetic tree based on amino acid sequences of the studied aquaporins.The sums of branch lengths for an optimal tree totaled 12.5. The tree reveals the formation of five groups, designated as letters from A to E.
Based on branch distances, the phylogenetic tree presented the formation of five distinct groups, denoted as letters A to E. Multiple sequence alignments were performed using the Praline server to further explore the similarities within each group. Group A consisted of fungal species with a shared identity of 39%, whereas group B comprised mammalian aquaporins with a high identity of 95%. Group C was composed of two mite species with an identity of 53%, group D encompassed plant sources with 40% identity, and group E included pathogens and mites with an identity of 31%. Notably, this analysis revealed significant allergen sources in tropical regions, such as D. pteronissinus and B. tropicalis.
Species such as P. americana, S. aureus, E. coli, B. germanica, and M. domestica did not fit within any specific group because of their substantial branch distances from the root of clustering. Consequently, they were categorized into independent groups. Among the identified groups, those most closely related to the aquaporins of A. fumigatus and humans were found in groups A and B. Additionally, noteworthy proximity was observed between E. coli and D. farinae in group B.
The guidance provided by the phylogenetic tree and the results of paired alignments of the EMBOSS needle and aquaporin sequences were selected with a score ≥380. This threshold ensured an identity greater than 30% for each aquaporin sequence. Subsequently, paired alignments of the selected aquaporins against A. fumigatus were performed using Praline. This approach provides a more visually informative result, enabling a better understanding of the conserved regions in each sequence ( Figure 2). By employing this screening process, we can identify more conserved sequences, thus increasing the likelihood of identifying regions that may contain potential epitopes.
Sequence alignment of aquaporins from A. fumigatus and humans was performed using Praline.The colors used in the alignment represent the level of sequence conservation, with blue indicating lower conservation and red indicating higher conservation. In the accompanying image, highlighted boxes denote the most conserved regions in the sequence, with each color (red, yellow, and green) representing the degree of conservation in descending order.
Aquaporin 3D models
Using PyMOL, we visualized the structures of the 12 selected aquaporin proteins. They were represented in the form of “cartoons” and were color-coded using the spectrum. These models exhibit the characteristic structure of aquaporins, consisting of six alpha helices connected by extracellular and intracellular “loops” that form the water and substance transport channel ( Figure 3).
Structures of aquaporins obtained from sources including PDB, Uniprot, AlphaFold, and Swiss-model. Their clustering corresponds to the arrangement in the phylogenetic tree. (A) This cluster comprises aquaporins from fungi. (B) Aquaporins from animals, except for E. coli, belong to this group.
Moreover, certain structures display distinctive extensions in their amino acid chains, resulting in variations in their tertiary structures. Examples include S. cerevisiae and E. caballus. Consequently, during epitope exploration, most of these regions were exposed, leading to the exclusion of these sequences as potential allergenic sources due to the absence of epitopes in the conserved regions of interest. This minimized the likelihood of cross-reactivity.
Prediction of linear and discontinuous epitopes
Using the 3D structures of A. fumigatus, Homo sapiens, Dermatophagoides farinae, Canis lupus familiaris, Felis catus, Equus caballus, Bos taurus, Mus musculus, Malassezia Sympodialis, Saccharomyces cerevisiae, Penicillium chrysogenum, and Escherichia coli, we predicted the linear and discontinuous epitopes for each aquaporin ( Table 3). We utilized PyMOL to analyze the identified linear and conformational epitopes, and thoroughly examined the distinct regions within each of these 3D structures. These epitopes are marked with the representative colors in the 3D structure. Although this characterization was performed for all 12 studied aquaporins, this work focused only on the epitopes found in the aquaporins of interest ( A. fumigatus and AQP3), as shown in Figures 4 and 5.
Table 3.: Lineal and discontinuous epitotpes from A. fumigatus and human aquaporin 3 (AQP3).
Depiction of predicted linear epitopes.All epitopes are color-coded on the surface models. Red, green, blue, and yellow correspond to the highest values, respectively.
Representation of predicted discontinuous epitopes from A. fumigatus aquaporin and AQP3. All epitopes are highlighted with color on the surface models, where red, green, and blue signify the highest values, respectively.
Furthermore, the PyMOL visualization program allowed us to calculate the root mean standard deviation (RMSD), which measures the structural similarity between two aligned objects, specifically the proteins of interest ( Figure 6). The closer the RMSD value is to zero, the stronger the molecular similarity between the proteins. We obtained an RMSD value of 1.003, indicating a significant molecular similarity.
The structural overlap of A. fumigatus aquaporin (magenta) and human aquaporin AQP3 (cyan) is depicted in a cartoon model. The root-mean-square deviation (RMSD) is 1.003.
In addition, we identified epitopes that contained conserved sequences. In the case of fungal aquaporins, Linear Epitope (LE) 2 was identified, whereas in human aquaporins, LE 3 was found. Individual analysis revealed highly conserved fragments with more than four residues. For fungal aquaporins, the conserved residue is 80QVTLSKG86, whereas for human aquaporins, it is 45QVVLSRG52. The percentage of identity and similarity between these fragments was 71.4% and 85.7%, respectively, suggesting that these residues were highly likely to be antigenic patches ( Figures 7 and 8).
Potential similar antigenic patches in linear epitopes of A. fumigatus aquaporin and AQP3.
Identification of transmembrane functional activity domains in the aquaporin proteins of human AQP3 and the fungus A. fumigatus through color-coded assignment in their secondary structure.
Discussion
Numerous diseases, including atopic dermatitis, bullous pemphigoid, chronic spontaneous urticaria, and multiple sclerosis, are associated with the presence of IgE autoantibodies. In the context of allergic responses intertwined with autoimmunity, researchers have categorized autoantigens that bind to IgE into three functional groups: 1) autoantigens that share sequence homology with environmental allergens, 2) autoantigens that lack sequence homology with known environmental allergens, and 3) chemically modified autoantigens. However, the identification of environmental allergens with homology to human proteins and their influence on the allergic response vary among these autoantigens, and their clinical significance often remains unclear. ^ 19 ^
Aquaporin is a promising protein candidate for studying cutaneous, autoimmune, and allergic diseases, including atopic dermatitis (AD), because of its involvement in epidermal hydration and transportation of various substances such as glycerol, salts, and exocrine fluids. These functions contribute to skin dryness, which, in turn, facilitates the filtration of allergens. ^ 14 ^ This protein is widely present in numerous organisms, and exhibits a diverse range of isoforms and functions across different taxonomic groups.
Recent research has identified the presence of aquaporins in allergenic species such as dust mites. ^ 20 ^ Mao et al. characterized the proteome of Dermatophagoides farinae, and two-dimensional immunoblotting revealed the binding of IgE antibodies to a band with the same isoelectric point and molecular weight as the aquaporin family, suggesting its potential role as an allergen. ^ 21 ^
On the other hand, in a systematic review, a total of 32 articles were examined to assess the role of the microbiota in atopic dermatitis. The microbiome profile revealed substantial variation in bacterial diversity along with an increased presence of fungi. Notably, there was diversification and expansion of species belonging to the genus Aspergillus within the fungal group, whereas a reduction in the number of Malassezia spp. was observed. Research has also demonstrated that A. fumigatus is a prominent source of sensitization in patients with severe atopic dermatitis. Additionally, the presence of new allergens from this species is believed to have a significant impact on the severity of the condition. ^ 22 ^
The comprehensive analysis of aquaporins provides guidance for considering other potential allergenic sources beyond the main aquaporins studied, namely AQP3 and A. fumigatus aquaporin. We conducted a comparative analysis of 25 aquaporin sequences from various sources, including A. fumigatus aquaporin and human aquaporin 3 (AQP3). A comparison between A. fumigatus aquaporin and AQP3 revealed an identity of 32.6% and similarity of 47.5%, indicating a relevant level of homology between these two proteins despite the evolutionary gap and taxonomic diversification among species. Notably, human AQP3 exhibited the maximum identity and similarity values of 95.5% and 98.6%, respectively. These high values could be attributed to the inclusion of aquaporins from mammalian allergen sources. Given that these species belong to a taxonomic group closer to humans, it is understandable that the APQs are more conserved.
Five clades were constructed for the phylogenetic analysis. A and B showed the highest identity with A. fumigatus aquaporins and AQP3. The alignment results suggest that divergence among aquaporins primarily occurs in smaller species, while it remains conserved in mammals. In addition, the presence of aquaporin isoforms, each with special functions and locations, such as AQP3, indicates functional and phenotypic variability among different cell types within the same organism. However, despite this diversity, the identity of aquaporins within the clades revealed values higher than 30%, particularly in clade B (95%), indicating that if the protein is recognized by antibodies, potential cross-reactivity among these species can occur.
Only a limited number of studies have investigated the relationship between aquaporins derived from allergenic sources and their potential relevance in human diseases. Zhou et al. conducted an assessment using transcriptomics to identify potential AQPs in Blomia tropicalis using molecular cloning techniques, followed by a comparative analysis of identity among mite aquaporin sequences and human aquaporins. The author identified five putative AQP-coding sequences, known as BlotAQP1-5, which were indexed into all three subgroups: AQPs, aquaglyceroporins, and superAQPs. BlotAQP1, BlotAQP2, and BlotAQP5 were clustered with the aquaglyceroporins hAQP3, hAQP7, hAQP9, and hAQP10, with a common sequence consensus ‘N-G-NPSRD-PRL.’ The molecular weight and isoelectric point of these molecules were consistent with the findings reported by Mao et al. in their two-dimensional electrophoresis study. ^ 23 ^ This observation suggests the potential recognition of aquaporins, specifically aquaglycerins such as AQP3, by IgE in patients with allergies. These results are consistent with those presented in our findings, where despite the evolutionary distance between species, there is an identity exceeding 30% and consensus sequences that may serve as potential antigenic patches.
In contrast, in regions where higher conservation was observed among the sequences, neither linear nor conformational epitopes were identified. This is attributed to the fact that these sequences are in the transmembrane region of the protein, which limits their accessibility to antibodies. However, despite the absence of epitopes, these conserved regions are interesting. During cell lysis caused by mechanical injury or microbial effects, these proteins can be linearly exposed, leading to the emergence of neoepitopes that can be recognized by IgE or phagocytic cells. ^ 24 ^
Although little is known about the role of AQP3 as an allergen or autoantigen, other aquaporins have been studied for their relevance as antigens in autoimmune diseases. Sagan et al. investigated the relevance of AQP4 as an autoantigen in neuromyelitis optica, recognized by T lymphocytes, in an animal model. ^ 25 ^ This aquaporin is a water channel expressed in astrocytes in areas in contact with the blood-brain barrier, and 75% of patients with neuromyelitis have antibodies capable of recognizing AQP4. Therefore, the relevance of human aquaporins found in other tissues, such as AQP3 in the skin, can be considered as potential autoantigens and, through molecular mimicry, triggers relevant skin diseases such as atopic dermatitis. ^ 26 ^ Evidence indicates cross-reactivity with aquaporins expressed by bacteria and mycobacteria, including Escherichia coli and Mycobacterium species, in relation to this specific autoantigen. ^ 27 ^ This indicated that antibodies targeting aquaporins from A. fumigatus could potentially recognize and target this autoantigen. In a previous study, we found that AQP4 exhibits only one predicted cross-reactive epitope. It has been postulated that the recognition of a single epitope by autoantibodies could trigger an epitope spreading mechanism, leading to the involvement of additional autoantigens in the immune response. ^ 28 ^ For both A. fumigatus aquaporins and AQP3, a total of four linear epitopes and three discontinuous epitopes were described. Structural superimposition highlighted a notable degree of similarity between the proteins, indicating that the epitopes could reside within the corresponding antigenic regions. This observation supports the idea of a potential molecular mimicry between these proteins.
It is important to acknowledge that our study has several limitations. In silico modeling and epitope prediction analyses are not definitive, and there may be variations in the actual structure compared with our proposed models. Nonetheless, bioinformatic analyses offer several advantages in directing research resources efficiently. They play a crucial role in the preliminary assessment of hypotheses and help to determine whether it is justified to proceed with in vitro or ex vivo experiments.
Conclusion
During alignment of aquaporin sequences from A. fumigatus and AQP3, specific conserved epitopes were observed. Analysis revealed that the local sequence identity exceeded 70%, indicating molecular mimicry in these regions of both aquaporins. Further investigation of epitopes and domain identification revealed the most conserved region in transmembrane domains 12 and 14 of the respective aquaporins. This study represents a groundbreaking contribution to the exploration and analysis of aquaporin as a potential allergen and autoallergen in atopic dermatitis, as it is the first known study to focus on this subject. To confirm cross-reactivity, future in vitro studies are needed to demonstrate the IgE-binding capacity in these regions.
By acquiring knowledge regarding the sequences and structures of potential epitopes from different allergenic sources, we can modify and synthesize these molecules for future in vitro and in vivo studies. This not only expands the literature in the field of immunotherapy but also facilitates the diagnosis and treatment of autoimmune diseases such as atopic dermatitis. It is important to note that in silico tests optimize these studies by providing rapid results and circumventing the high costs associated with laboratory testing.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Múnera M Sanchez A Buendía E : Autoantigens in atopic dermatitis: The characterization of autoantigens and their diagnostic value. Translational Autoimmunity. Elsevier;2023; pp.37–48. 10.1016/B 978-0-323-85389-7.00019-3 · doi ↗
- 2Mc Kenzie C Silverberg JI : The prevalence and persistence of atopic dermatitis in urban United States children. Ann. Allergy Asthma Immunol. 2019;123(2):173–178.e 1. 10.1016/j.anai.2019.05.014 31128232 · doi ↗ · pubmed ↗
- 3Luger T Amagai M Dreno B : Atopic dermatitis: Role of the skin barrier, environment, microbiome, and therapeutic agents. J. Dermatol. Sci. 2021;102(3):142–157. 10.1016/j.jdermsci.2021.04.007 34116898 · doi ↗ · pubmed ↗
- 4Sánchez J Calvo V Sánchez A : Sensitization to 10 mites in a tropic area. Der p and Der f are important risk factor for sensitization to other mites from Pyroglyphidae, Acaridae, Chortoglyphidae, and Glyciphagidae families. Rev. Alerg. Mex. 2017;64(2):153–162. 10.29262/ram.v 64i 2.243 28658723 · doi ↗ · pubmed ↗
- 5Olsson M Broberg A Jernås M : Increased expression of aquaporin 3 in atopic eczema. Allergy. 2006;61(9):1132–1137. 10.1111/j.1398-9995.2006.01151.x 16918518 · doi ↗ · pubmed ↗
- 6Sánchez J Páez B Macías-Weinmann A : Puntos clave en el tratamiento de la dermatitis en Latinoamérica. El Consenso SLAAI. Rev. Alerg. Mex. 2015;62(3):226–233. 10.29262/ram.v 62i 3.87 26239333 · doi ↗ · pubmed ↗
- 7Cardona R Sánchez A Larenas-Linnemann D : Extractos alergénicos para inmunoterapia en Latinoamérica. Rev. Alerg. Mex. 2018;65(1):25–40. 10.29262/ram.v 65i 1.287 29723939 · doi ↗ · pubmed ↗
- 8Sánchez J Sánchez A Cardona R : Critical review of ISAAC results for atopic dermatitis in tropical cities. Rev. Alerg. Mex. 2018;65(4):389–399. 10.29262/ram.v 65i 4.341 30602209 · doi ↗ · pubmed ↗