Lipopolysaccharide Core Truncation in Invasive Escherichia coli O157:H7 ATCC 43895 Impairs Flagella and Curli Biosynthesis and Reduces Cell Invasion Ability
Haiqing Sheng, Robinson J. Ndeddy Aka, Sarah Wu

TL;DR
This study shows that changes in the LPS structure of E. coli O157:H7 reduce its ability to form biofilms and invade host cells.
Contribution
The novel contribution is identifying how specific LPS-core mutations impair flagella and curli biosynthesis, reducing cell invasion.
Findings
LPS-core mutants failed to form biofilms, showing the role of core oligosaccharides in biofilm formation.
Inner-core mutants ΔwaaF and ΔwaaC lost flagella and curli production and showed reduced cell invasion.
Invasion of epithelial cells decreased by 100-fold in inner-core mutants compared to wild type.
Abstract
Escherichia coli O157:H7 (E. coli O157) is known for causing severe foodborne illnesses such as hemorrhagic colitis and hemolytic uremic syndrome. Although E. coli O157 is typically regarded as an extracellular pathogen and a weak biofilm producer, some E. coli O157 strains, including a clinical strain ATCC 43895, exhibit a notable ability to invade bovine crypt cells and other epithelial cells, as well as to form robust biofilm. This invasive strain persists in the bovine host significantly longer than non-invasive strains. Various surface-associated factors, including lipopolysaccharides (LPS), flagella, and other adhesins, likely contribute to this enhanced invasiveness and biofilm formation. In this study, we constructed a series of LPS-core deletion mutations (waaI, waaG, waaF, and waaC) in E. coli O157 ATCC 43895, resulting in stepwise truncations of the LPS. This approach enabled…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
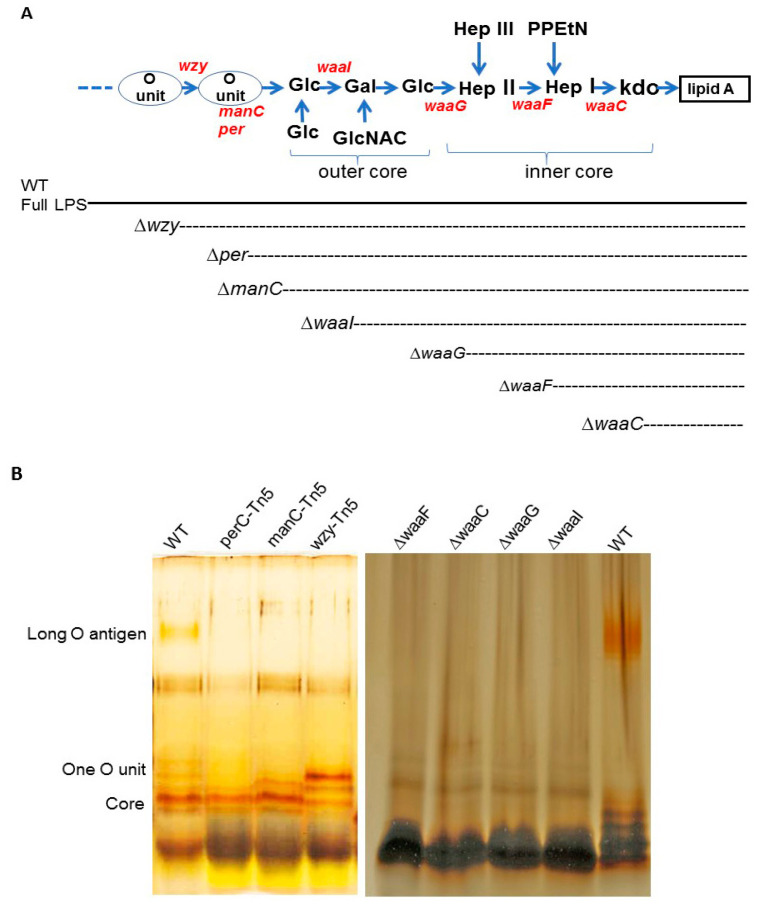 Figure 1
Figure 1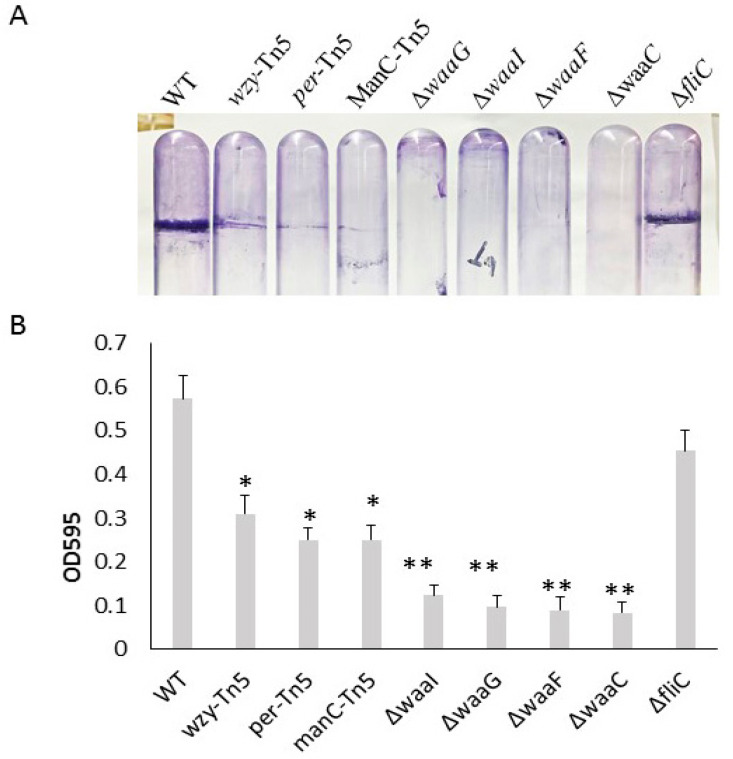 Figure 2
Figure 2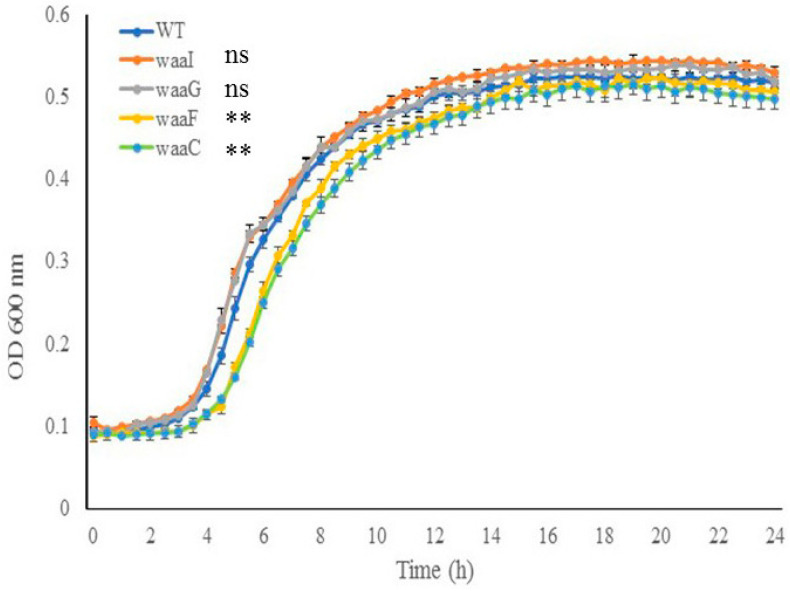 Figure 3
Figure 3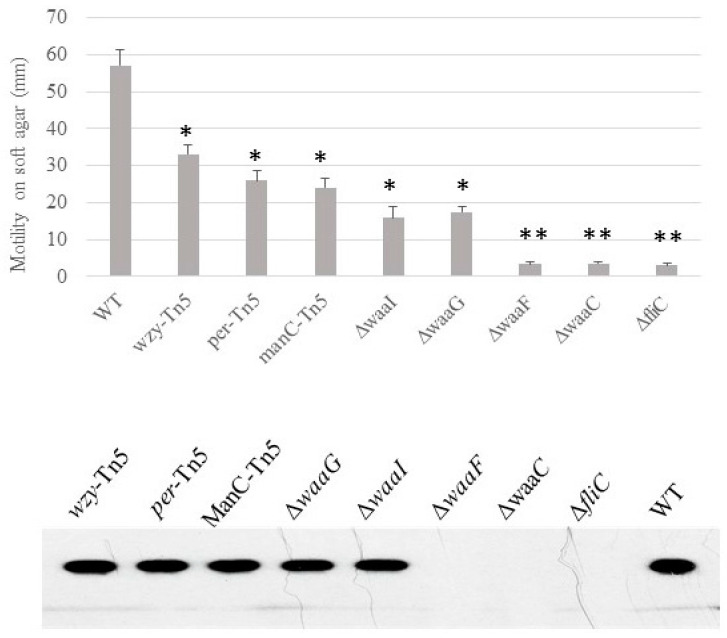 Figure 4
Figure 4 Figure 5
Figure 5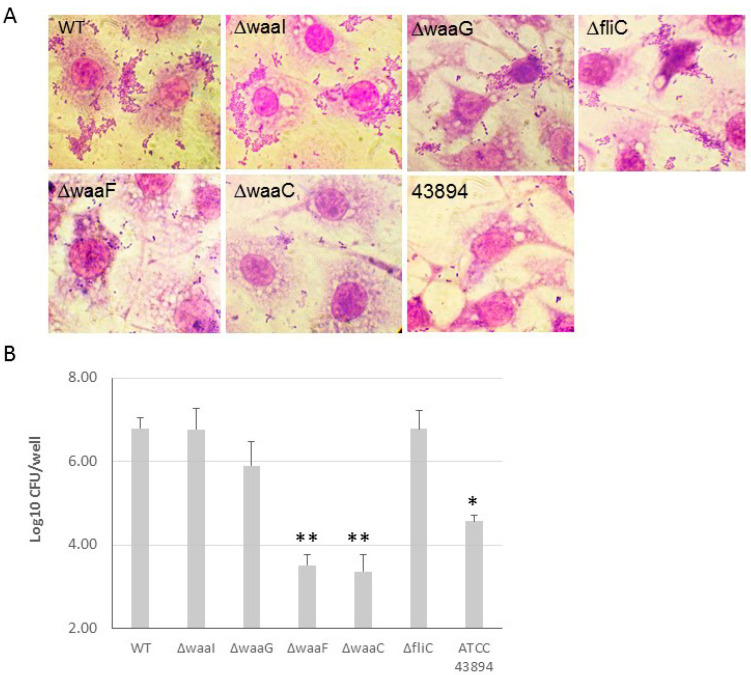 Figure 6
Figure 6- —USDA National Institute for Food and Agriculture Foundational and Applied Science Program
- —USDA NIFA Hatch
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsEscherichia coli research studies · Bacterial Genetics and Biotechnology · Vibrio bacteria research studies
1. Introduction
Escherichia coli O157:H7 (E. coli O157) is a Shiga-toxin-producing pathogen that poses significant public health risks due to its potential to cause severe foodborne illnesses, including hemorrhagic colitis and hemolytic uremic syndrome [1,2]. This bacterium colonizes the bovine recto-anal junction in healthy cattle, which act as primary reservoirs and are common source of foodborne infections [3,4]. The economic impact of E. coli O157 contamination in beef and fresh produce is considerable, with estimates suggesting that medical costs and lost productivity in the United States alone may reach USD 400 million annually [5]. Additionally, the economic burden on meat producers and vegetable growers can include product loss and negative publicity associated with product recalls [6]. Attachment to biological surfaces is a crucial first step in colonization, facilitated by various surface-associated factors such as lipopolysaccharides (LPS) [7], fimbriae [8], flagella, and other adhesins such as intimin [9,10,11], autotransporters [12], and curli [13]. These factors contribute to biofilm formation, cell adherence and invasion, and persistence in the bovine host.
LPS is an essential surface component of the outer membrane in Gram-negative bacteria. E. coli LPS is composed of three distinct regions: the O-antigen, the core oligosaccharide (OS), and lipid A [14]. In E. coli O157, the LPS core-OS is of the R3 type, which differs distinctly from that of E. coli K-12 [15]. The inner core of the R3 type comprises 3-deoxy-D-manno-oct-2-ulosonic acid (Kdo) and ADP-heptose residues, while the outer core is constructed of hexoses and 2-acetoamido-2-deoxy-hexoses. ADP-heptose is a crucial component of the LPS inner core, linking the outer part of LPS to Kdo between the Kdo2-lipid A and O-antigen. The R3 core structure involves several genes in the waa operon, including waaC, waaF, waaQ, waaG, waaO, waaR, waaY, waaZ, rfaE, and rfaD [15]. These genes collectively contribute to adding various sugar residues and modifications to the core oligosaccharide, resulting in a stable and functional LPS molecule. Mutations or deletions in these genes can lead to truncated LPS structures, which impact the overall functionality of the bacterial outer membrane [16,17]. The structural integrity of the outer membrane is crucial for the assembling and functionality of other surface structures such as flagella and curli, and for interactions with the host environment [14,18,19,20]. Disruptions in LPS-core synthesis can lead to significant alterations in bacterial behavior and pathogenic potential. Understanding the roles of these genes can provide valuable insights into how modifications in LPS structure can influence the virulence and survival strategies of E. coli O157.
E. coli O157 is typically regarded as an extracellular pathogen and a weak biofilm producer [21,22]. However, some E. coli O157 strains, including a clinical strain ATCC 43895 (E. coli O157 43895), exhibit a notable ability to invade bovine crypt cells and other epithelial cells, as well as to form robust biofilm at 37 °C [23]. The invasive strain E. coli O157 ATCC persists significantly longer in the bovine host than non-invasive E. coli O157 strains [13]. The biofilm formation of E. coli O157 43895 is associated with LPS and curli. Its curli production promotes cell invasion [13]. Curli fimbriae, composed of polymerized amyloid protein, are produced by many E. coli and Salmonella typhimurium strains, preferentially at relatively lower temperatures (25–30 °C) [24,25]. We previously showed that E. coli O157 43895 cells produce curli at 37 °C [13]. The influence of LPS on surface structures such as flagella and curli, and consequently on invasion ability, is not yet fully understood. Our previous study showed that deletion of the genes involved in biosynthesis of E. coli O157 43895 O-antigen does not affect curli production [13]. However, other LPS biosynthesis genes, such as waaG, rfbH, and lpxM have been implicated as important for curli production [26,27,28].
In this study, we constructed derivative strains of E. coli O157 43895 with a series of ordered LPS-core deletion mutations, leading to stepwise truncations of the LPS. We investigated how truncating the LPS core-OS affects the behaviors of invasive E. coli O157 43895, particularly its interaction with epithelial cells. By deleting specific genes involved in LPS core-OS synthesis and comparing these strains with the wild type and previously created O-antigen mutants, we aimed to understand the impacts on key virulence factors, such as flagella and curli biosynthesis, as well as the bacterium’s ability to invade host cells. The findings offer valuable insights into the molecular mechanisms underlying E. coli O157 persistence and pathogenicity, potentially identifying targets for interventions to reduce bovine carriage and human infections caused by this pathogen.
2. Results
Construction of E. coli O157 43895 LPS mutants and comparison of their LPS structures. We generated a series of LPS mutants of invasive E. coli O157 43895, each exhibiting progressively shorter LPS structures, as illustrated in Figure 1. Four genes involved in core biosynthesis were individually deleted from the chromosome of E. coli O157 43895, resulting in the mutants ΔwaaI, ΔwaaG, ΔwaaF, and ΔwaaC. Additionally, three O-antigen mutants (wzy-Tn5, per-Tn5, and manC-Tn5) generated previously by Tn5 mutagenesis were included in this study for their LPS structural analysis and comparison [13]. The expected structures of LPS in these seven E. coli O157 43895 mutants are shown in Figure 1A.
The mutations commenced with a wzy mutant, characterized by LPS containing only a single O-antigen unit, and culminated in a waaC mutant with an inner core truncated to the Kdo residues (Figure 1A). Each mutation yielded a strain with an LPS length consistent with our expectations. The wild-type strain produced complete LPS molecules with long O-antigens, while the LPS of the waaC mutant comprised only lipid A and Kdo. The LPS synthesized by the other mutants exhibited varying degrees of core and/or O-antigen sugar presence, as visualized in the gel (Figure 1B).
Comparison of biofilm formation of E. coli O157 43895 LPS and flagella mutants. We previously demonstrated that the invasive E. coli O157 43895 forms robust biofilm, even at an elevated temperature of 37 °C, compared with non-invasive strains [13]. In this study, we compared the biofilm-formation abilities of O-antigen, core-OS, and flagella mutants of E. coli O157 43895 with the WT by observing pellicle biofilms and conducting quantification measurement. The WT, along with the O-antigen mutants wzy-Tn5, per-Tn5, and manC-Tn5, the LPS-core mutants ΔwaaI, ΔwaaG, ΔwaaF, and ΔwaaC, and the flagella mutant ΔfliC, were grown in LB broth overnight at 37 °C under static condition. After 48 h of culture, pellicles and rings were visible in the WT and the flagella mutant (Figure 2A). The O-antigen mutants (wzy-Tn5, per-Tn5, and manC-Tn5) also formed visible rings, though they were less pronounced compared with the WT. In contrast, truncation of the LPS core abolished pellicle biofilm formation, as no rings were observed in the cultures of the core mutants (ΔwaaI, ΔwaaG, ΔwaaF, and ΔwaaC) (Figure 2A).
In the quantification assay, the WT and mutant strains were grown in a 96-well microtiter plate for 24 h. Biofilms were stained with crystal violet and measured using a spectrophotometer. The ΔfliC mutant demonstrated a biofilm formation ability with an OD595 reading of 0.46, comparable to the WT’s reading of 0.54 (p > 0.05), suggesting that biofilm formation of E. coli O157 43895 is not dependent on flagella. In contrast, biofilm formation by O-antigen mutants (wzy-Tn5, per-Tn5, and manC-Tn5) was significantly reduced compared with the WT (p ≤ 0.01), with the OD595 reading ranging from 0.25 to 0.31 (Figure 2B). Biofilm formation was even further diminished in the LPS-core mutants (ΔwaaI, ΔwaaG, ΔwaaF, and ΔwaaC), with the OD595 reading in the 0.1 range (p ≤ 0.001). The reduction was consistent with the absence of a pellicle ring in the tube assays, indicating that these LPS-core mutants lost their ability to form biofilm. The significant difference in biofilm formation between the LPS core-OS mutants and the O-antigen mutants highlights the importance of an intact of LPS core for biofilm formation in E. coli O157 43895.
Cell growth of E. coli O157 43895 WT and LPS-core mutants. To investigate the impact of the LPS core on cell growth, core mutants and the WT were grown in MOPS media at 37 °C for 24 h with aeration. As shown in Figure 3, the two outer-core mutants, ΔwaaI and ΔwaaG, displayed growth comparable to the WT strain throughout the 24 h period. In contrast, the two LPS inner-core mutants, ΔwaaF and ΔwaaC, experienced an expanded lag phase and grew significantly slower than the WT during the logarithmic phase (4 h to 12 h) (p < 0.001); however, the two mutants were able to eventually catch up in growth rate during the stationary phase. These results suggest that the absence of the inner LPS core initially affects cell growth.
Motility, presence of flagella and curli. We previously demonstrated that the O-antigen mutant Δper of E. coli O157 43895 has impaired motility but can be induced to swim after extended incubation in 0.3% soft agar [7]. Here, we determined the relative ability of LPS-core mutants of E. coli O157 43895 to swim on soft agar and the presence of flagella. After 48 h of prolonged incubation, the O-antigen mutants and one of the outer-core LPS mutants ΔwaaI were able to swim to varying degrees on soft agar, though with impaired motility. In contrast, three core LPS mutants, ΔwaaG, ΔwaaF, and ΔwaaC, lost motility (Figure 4). Western-blot analysis indicated that these three mutants had defective motility due to lack of flagella (Figure 4).
Unlike non-invasive E. coli O157 strains, ATCC 43895 produces curli at 37 °C. To investigate the influence of the core-OS on curli production, we grew the LPS mutants of the invasive strain on CRI plates at 37 °C. After 24 h incubation, the WT and the O-antigen and outer-core mutants (ΔwaaI and ΔwaaG), and flagella mutant (ΔfliC) all produced curli, as evidenced by red coloration of the colonies due to Congo-red binding on the CRI plate. In contrast, the two inner-core LPS mutants (ΔwaaF and ΔwaaC) remained white, indicating a lack of curli production (Figure 5).
The effect of LPS core-OS of O157 43895 on cell adherence and invasion. E. coli O157 43895 has a strong ability to adhere to and invade epithelial cells compared with other E. coli O157 strains. We previously demonstrated that truncation of O-antigen in E. coli O157 43895 (wzy-Tn5, per-Tn5, and manC-Tn5) did not affect its cell adherence and invasion [13]. In this study, we examined the invasion ability of the isogenic LPS-core mutants ΔwaaI, ΔwaaG, ΔwaaF, and ΔwaaC, as well as the flagella mutant ΔfliC. The WT and mutants were co-cultured with MAC-T cells individually. Giemsa staining revealed that the outer-core mutants ΔwaaI and ΔwaaG, and mutant ΔfliC, exhibited a large number of bacteria adhering to the epithelial cells in an aggregative pattern, similar to WT. However, the two inner-core mutants, ΔwaaF and ΔwaaC, showed dramatically reduced adherence to MAC-T cells, with only a few sporadic bacteria adhering to the cells (Figure 6A). Subsequently, we assessed the ability of these mutants to invade epithelial cells. The WT and mutants were individually co-cultured with a monolayer of MAC-T cells in a 24-well plate at MOI of 10 for 3 h. The number of bacteria recovered after 2 h of gentamicin treatment indicated the number of intracellular bacteria. Among the four LPS-core mutants, ΔwaaF and ΔwaaC showed a more than 100-fold and 10-fold reduction in cell invasion, respectively, compared with WT E. coli O157 43895 and curli-deficient E. coli O157:H7 ATCC 43894. The log value of intracellular bacteria for the two inner-core mutants was significantly lower compared with the WT and the outer-core mutants ΔwaaI and ΔwaaG (p < 0.001), as well as to the non-invasive strain ATCC 43894 (p < 0.01).
3. Discussion
The LPS core of E. coli O157 belongs to the R3 type, which is preferentially associated with key virulence determinants such as Shiga toxins [29,30,31] and the adhesin intimin and its receptor Tir [32,33]. In this study, we systematically investigated the relationship between O-antigen/LPS-core length and biofilm formation, flagella and curli production, and cell adherence and invasion. Deletion of the waa or disruption of wba genes targeted in this study resulted in truncated LPS structures (Figure 1). The results demonstrate the critical role of the LPS core in various cellular functions of E. coli O157 43895.
Unlike non-invasive E. coli O157 strains, ATCC 43895 forms a robust pellicle biofilm at 37 °C. Pellicle biofilms are bacterial communities at the air–liquid interface, composed of extracellular polymeric substances such as polysaccharides, proteins, flagella, fimbriae, and nucleic acids secreted by the bacteria [34,35]. In some Gram-negative bacteria, LPS is crucial for biofilm matrix construction [36,37,38]. This extracellular matrix can enhance bacterial adherence to host tissues, facilitating colonization in bovine hosts and potentially leading to deeper tissue invasion [23]. Previous research has shown that disrupting the O-antigen biosynthetic genes (per, manC, and wzy) in E. coli O157 43895 and curli fibers contribute to biofilm formation [13]. In this study, we compared the biofilm formation abilities of O-antigen, LPS-core, and flagella mutants of E. coli O157 43895 with the wild type. We found that truncating the LPS core completely abolished biofilm formation, highlighting the critical role of a complete LPS core in maintaining biofilm integrity. The differences in biofilm formation among LPS-core and O-antigen mutants suggest additional factors beyond LPS-core length. Interestingly, despite lacking motility, the flagella fliC mutant still formed a robust pellicle biofilm, which was unexpected.
We analyzed the mobility and flagella presence in O-antigen and LPS-core mutants of invasive E. coli O157 43895. All mutants exhibited significantly reduced motility, with ΔwaaG, ΔwaaF, and ΔwaaC mutants completely losing motility. Western-blot analysis revealed that these mutants were unable to produce flagella. Similar findings were observed in E. coli K-12; LPS-core mutants showed no flagella on their cell surfaces under electron microscopy compared with the wild type [39]. Non-motile phenotypes in LPS-deficient strains have also been reported in other Gram-negative bacteria. For instance, deep rough mutants ΔrfaG and ΔrfaD in Salmonella, which lack outer- and inner-core components, respectively, showed dramatically impaired motility [40,41]. These results suggest that truncation of the LPS core affects flagella assembly in both E. coli and Salmonella. In contrast, truncated LPS-core mutants of Pseudomonas aeruginosa PAO1, such as those with rmlC, migA, and wapR deletions, exhibited reduced motility, but flagella assembly appeared intact [42].
We also assessed curli production in the O-antigen and LPS-core mutants using a Congo-red-binding assay. Compared with O-antigen and outer-core mutants, as well as the wild type, the ΔwaaF and ΔwaaC mutants, which are defective in the inner core, failed to produce curli on their surfaces. This finding is consistent with an earlier study that screened the Keio collection of single-gene deletions in E. coli K-12 using Congo-red indicator plates and identified LPS-core mutants of E. coli K-12 that were defective in curli production [43]. These results support the notion that LPS integrity is crucial for maintaining outer membrane stability and the proper assembly of cell-surface components. Truncation of the LPS core disrupts the outer membrane, leading to leakage of periplasmic contents into the extracellular space and alterations in outer membrane composition [44].
It remains unclear whether the altered LPS structure or the mutated genes involved in LPS-core biosynthesis directly caused the defects in flagella and curli fimbriae. The truncation of the LPS core might induce general cell-envelope stress, downregulating flagella and curli assembly in mutants like ΔwaaF and ΔwaaC, which could affect interactions with epithelial cells. Future studies will focus on these aspects.
We observed that ΔwaaF and ΔwaaC mutants, which lack heptose, experienced an extended lag phage at 37 °C, suggesting that heptose incorporation into the inner core is crucial for optimal cell growth, particularly during the early stages. Interestingly, Murata et al. reported that certain LPS synthetic genes involved in heptose biosynthesis (gmhA, gmhB, and gmhD) or its incorporation into the LPS inner core (waaC and waaF) are essential for bacterial growth at critical high temperatures in E. coli K-12 [45].
Lastly, the reduced adherence and invasion abilities of inner-core mutants ΔwaaF and ΔwaaC underline the necessity of a complete LPS core for effective interaction with host cells. These findings suggest that the LPS-core structure not only contributes to production of flagella and curli and supports growth but also plays a crucial role in pathogenesis by mediating adherence and invasion in invasive E. coli O157 43895.
In summary, this study underscores the multifaceted role of the LPS core in invasive E. coli O157 43895, affecting growth, motility, biofilm formation, and host cell interactions, with significant implications for understanding bacterial persistence and developing targeted interventions.
4. Materials and Methods
Bacterial strains, plasmids, media, and growth conditions. The bacterial strains, plasmids, and primers used in this study are listed in Table 1 and Table 2. Bacteria were grown in Luria–Bertani (LB) broth or agar at 37 °C, unless otherwise stated. When required, antibiotics (Sigma-Aldrich, St. Louis, MO, USA) kanamycin (Kan, 50 mg/mL), chloramphenicol (Cm, 30 mg/mL), or ampicillin (Amp, 100 mg/mL) were added to the media. Congo-red-dye-binding of curli was monitored on Congo-red-indicator (CRI) agar: 10 g Casamino acids, 1 g yeast extract, 20 g agar, 20 mg Congo red, and 10 mg Coomassie brilliant blue G-250/L (Sigma-Aldrich).
Mutant-strain construction. To create a mutant deficient in expression of LPS core-OS in E. coli O157 43895, the Lambda Red recombinase system [46] was used for gene deletion, as previously described (7). Oligonucleotide primers were purchased from Invitrogen (Carlsbad, CA, USA). The genes of the LPS core-OS of E. coli O157 43895 were independently replaced by a chloramphenicol resistance cassette (the cat gene from pKD3). The genes waaI, waaG, waaF, and waaC were selected for deletion based on their position in the waa cluster. As shown in Figure 1A, waaC encodes a heptosyltransferase I enzyme, which adds the first L-glycero-D-manno-heptose (Hep) residue to the Kdo (3-deoxy-D-manno-oct-2-ulosonic acid) molecule in the LPS core-OS; waaF encodes a heptosyltransferase II enzyme, responsible for the addition of the second Hep residue to the growing LPS core-OS oligosaccharide; both waaG and waaI encode a glucosyltransferase that adds a glucose (Glc) residue to the LPS core-OS (Figure 1A). In addition, H7 flagella mutant was made by deleting fliC for comparison. PCR primers for deletions of waaI, waaG, waaF, and waaC were designed based on the sequence of the waa cluster in the E. coli O157 43895 genome (accession number NZ_CP008957). Chloramphenicol-resistant (Cm^R^) recombinants were selected on LB agar plates containing chloramphenicol. The O-antigen mutants manC-Tn5, per-Tn5, and manC-Tn5*,* generated previously, were included in this study for comparison [13]. The failure of the LPS core-OS mutants to express the O157 antigen was confirmed by anti-O157 latex agglutination (Pro-Lab Diagnostics, Toronto, ON, Canada). The chloramphenicol resistance gene was eliminated. The replacement of each of the targeted genes by a nonpolar scar structure [46] and the mutations were verified by PCR.
LPS analysis. To confirm the loss of LPS core-OS in the mutants, LPS was isolated and analyzed using a previously described method [47]. Briefly, cells were grown in 3 mL LB broth to an optical density at 600 nm (OD600) of 1.0. The cells were then harvested by centrifugation, resuspended in 1.0 mL of phosphate-buffered saline, and incubated at 60 °C for 30 min. The suspension was then centrifuged at 12,000× g for 30 min, and the supernatant was mixed with an equal volume of Tricine sample buffer (Bio-Rad Laboratories, Hercules, CA, USA) and boiled for 10 min. Proteinase K was added to a final concentration of 0.5 mg/mL, and the sample was incubated at 60 °C for 60 min before being centrifuged at 16,000× g for 30 min. LPS was then analyzed by Tricine–SDS-PAGE and visualized by silver staining, as previously described [7].
Crystal violet (CV) biofilm assay. To assess the ability of E. coli O157 mutants to produce a pellicle biofilm at the air–liquid interface, a fresh colony was grown in LB broth overnight at 37 °C. This culture was then diluted 500-fold with fresh 4 mL LB in a 15 mL polystyrene tube and incubated overnight at 37 °C under static conditions. The content of each tube was removed. The tubes were stained with 5 mL of 1% crystal violet solution for 5 min. The dye was then removed, and the tubes were rinsed with water and air dried.
Biofilm quantification assays were performed using a standard microtiter assay [48]. The overnight culture was inoculated into duplicate 96-well microtiter plates containing minimal salt medium (MSM) with 0.04% glucose and incubated for 24 h without agitation at 37 °C. Each isolate was inoculated in triplicate. After incubation, the titer plates were washed with water and stained with 1% crystal violet for 15 min at room temperature. Plates were then rinsed vigorously with water again to remove unattached cells and residual dye. Biofilm formation was evaluated by measuring the absorbance of the solubilized dye in 95% ethanol at a wavelength of 595 nm using a PowerWave XS reader (Bio-Tek, Winooski, VT, USA).
Growth curves in MOPS media. Three isolates of each strain were cultured overnight in LB broth at 37 °C with shaking at 150 rpm. Overnight cultures were diluted with LB to 0.1 OD600 and grown for 4 h at 37 °C. Cells were pelleted then washed twice with 1× PBS and resuspended in MOPS media [49] (1× MOPS supplemented with 0.5% glucose, 1 µg/mL thiamine, 0.1 mM K2HPO4). The cells in MOPS were diluted to approximately 0.1 OD600. Two hundred microliters of cells was pipetted in technical replicates of 3 in a microtiter plate and growth was monitored using the PowerWave XS reader at 37 °C with continuous shaking. Optical density measurements were collected at a wavelength of 600 nm every 30 min, 5 s after shaking stopped for 24 h.
Motility assays. The swimming motility of the wild-type and the mutant strains was assessed using tryptone swarm plates containing 1% Bacto Tryptone, 0.5% NaCl, and 0.3% agar. Five microliters of an overnight culture was point-inoculated into swarm plates and incubated at 37 °C for up to 40 h. The diameter of the motility halo was measured.
Adherence and invasion assays. The bacterial internalization of MAC-T cells was measured using a standard gentamicin protection assay. First, 24-well tissue culture plates (Corning Costar, NY, USA) were seeded with 10^4^ MAC-T cells and incubated in 5% CO_2_ at 37 °C until cells were confluent. The MAC-T cell monolayers were then washed twice with Hank’s buffered saline solution (HBSS), and approximately 2 × 10^6^ E. coli O157 cells (multiplicity of infection, MOI, 10:1) in cell culture medium without antibiotics or fetal bovine serum (1 mL) were added. After 3 h at 37 °C, unattached extracellular bacteria were removed by suction, and the monolayers were washed three times with HBSS. Fresh medium containing 100 µg/mL gentamicin was added to kill extracellular bacteria. After an additional 2 h incubation, the monolayers were washed three times with HBSS without Ca^2+^ or Mg^2+^. Epithelial cells were lysed by adding 100 µL 0.5% trypsin-EDTA and 900 µL 0.05% Triton X-100/well for 5 min. Bacterial invasion was quantified by counting the CFUs recovered/well on LB agar. To microscopically view the interaction of the bacteria with MAC-T cells, the cells on cover slips were fixed with methanol, stained with Giemsa stain (0.4% w/v, Sigma-Aldrich), and examined microscopically under oil immersion at 100X.
Flagella isolation and immunoblotting. Flagella were sheared from E. coli O157 derivatives using a standard protocol [50] with slight modification. Briefly, E. coli O157 strains were cultured on motility agar for 40 h, as mentioned above. LB was inoculated with 20 µL agar plugs from the edge of the motility halo and incubated at 37 °C with shaking for 30 h. Bacterial pellets were collected by centrifugation (5000× g) for 15 min at 4° C, and resuspended with 1 mL cold PBS with protease inhibitor (Sigma-Aldrich) at appropriate concentration. The microtubes with the bacterial suspension were put on ice for 15 min, and placed a FastPrep FB120 Cell Disruptor (Abiogene, CA, USA) at a 5.5 m/sec speed for 30 s. The vigorous vortex step was repeated 4 times and the tubes were put on ice for 10 min at each interval. The supernatants were transferred into clean tubes after centrifuge (8000× g) for 15 min at 4 °C. The cell fibers were pelleted by centrifugation (41,000× g for 3 h at 4 °C). To verify the presence of flagella, sheared fibers were monomerized according to a published protocol [51]. Flagella preparations were incubated at 70 °C for 15 min and then were examined by Western blotting with the 15D8 monoclonal antibody specific for enteric flagella [52]. Cell suspensions from overnight cultures grown in LB were extracted by boiling, fractionated by SDS-PAGE gel electrophoresis, transferred onto PVDF membranes using a Mini Trans-Blot electrophoretic transfer cell (Bio-Rad, Redmond, WA, USA), and probed with 15D8 (1:2000).
Statistical analysis. Statistical analyses were performed by using GraphPad Prism software 7 (San Diego, CA, USA). Details of sample size, test used, error bars and statistically significant cutoff are presented in the text or figure legends.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Paton J.C. Paton A.W. Pathogenesis and diagnosis of Shiga toxin-producing Escherichia coli infections Clin. Microbiol. Rev.19981145047910.1128/CMR.11.3.4509665978 PMC 88891 · doi ↗ · pubmed ↗
- 2Karmali M.A. Steele B.T. Petric M. Lim C. Sporadic cases of haemolytic-uraemic syndrome associated with faecal cytotoxin and cytotoxin-producing Escherichia coli in stools Lancet 1983161962010.1016/S 0140-6736(83)91795-66131302 · doi ↗ · pubmed ↗
- 3Naylor S.W. Low J.C. Besser T.E. Mahajan A. Gunn G.J. Pearce M.C. Mc Kendrick I.J. Smith D.G. Gally D.L. Lymphoid Follicle-Dense Mucosa at the Terminal Rectum Is the Principal Site of Colonization of Enterohemorrhagic Escherichia coli O 157:H 7 in the Bovine Host Infect. Immun.2003711505151210.1128/IAI.71.3.1505-1512.200312595469 PMC 148874 · doi ↗ · pubmed ↗
- 4Sheng H. Davis M.A. Knecht H.J. Hovde C.J. Rectal administration of Escherichia coli O 157:H 7: Novel model for colonization of ruminants Appl. Environ. Microbiol.2004704588459510.1128/AEM.70.8.4588-4595.200415294790 PMC 492365 · doi ↗ · pubmed ↗
- 5Frenzen P.D. Drake A. Angulo F.J. Economic cost of illness due to Escherichia coli O 157 infections in the United States J. Food Prot.2005682623263010.4315/0362-028X-68.12.262316355834 · doi ↗ · pubmed ↗
- 6Buzby J.C. Roberts T. The economics of enteric infections: Human foodborne disease costs Gastroenterology 20091361851186210.1053/j.gastro.2009.01.07419457414 · doi ↗ · pubmed ↗
- 7Sheng H. Lim J.Y. Watkins M.K. Minnich S.A. Hovde C.J. Characterization of an Escherichia coli O 157:H 7 O-antigen deletion mutant and effect of the deletion on bacterial persistence in the mouse intestine and colonization at the bovine terminal rectal mucosa Appl. Environ. Microbiol.2008745015502210.1128/AEM.00743-0818552194 PMC 2519267 · doi ↗ · pubmed ↗
- 8Vogeleer P. Tremblay Y.D.N. Jubelin G. Jacques M. Harel J. Biofilm-Forming Abilities of Shiga Toxin-Producing Escherichia coli Isolates Associated with Human Infections Appl. Environ. Microbiol.2015821448145810.1128/AEM.02983-1526712549 PMC 4771338 · doi ↗ · pubmed ↗