The redescription and complete mitogenomes of two Oxycarenus species (Hemiptera, Oxycarenidae) and phylogenetic implications

Abstract
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
 Figure 1
Figure 1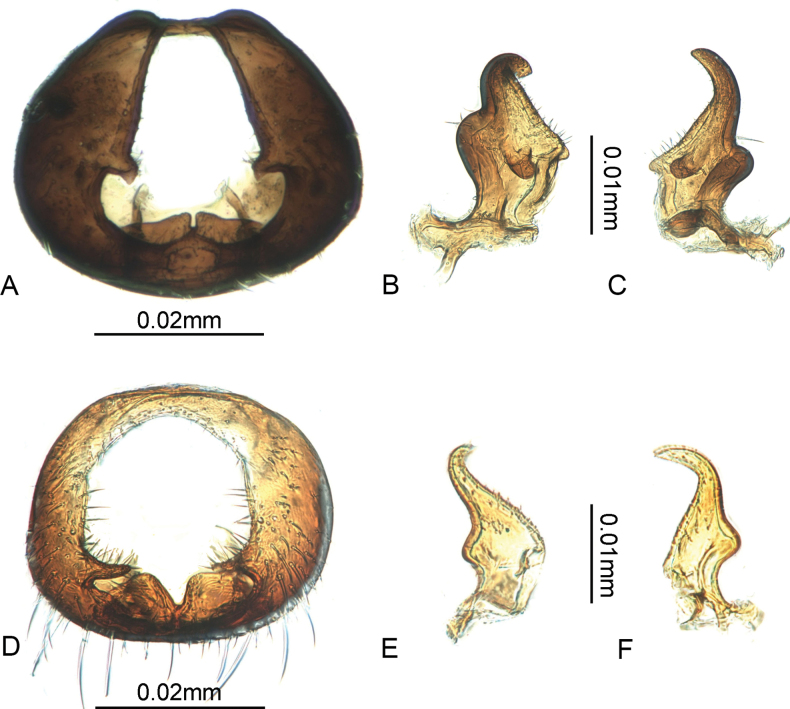 Figure 2
Figure 2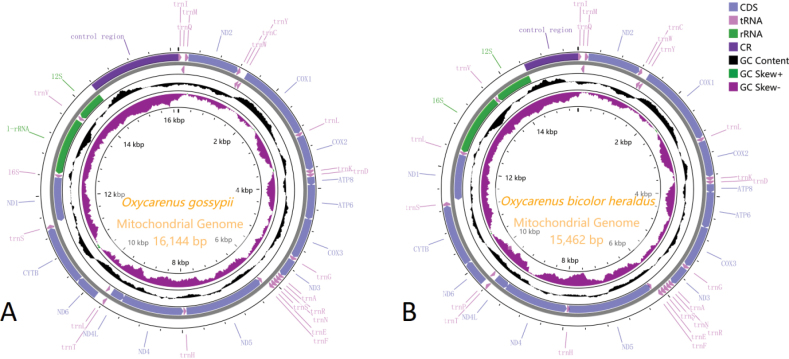 Figure 3
Figure 3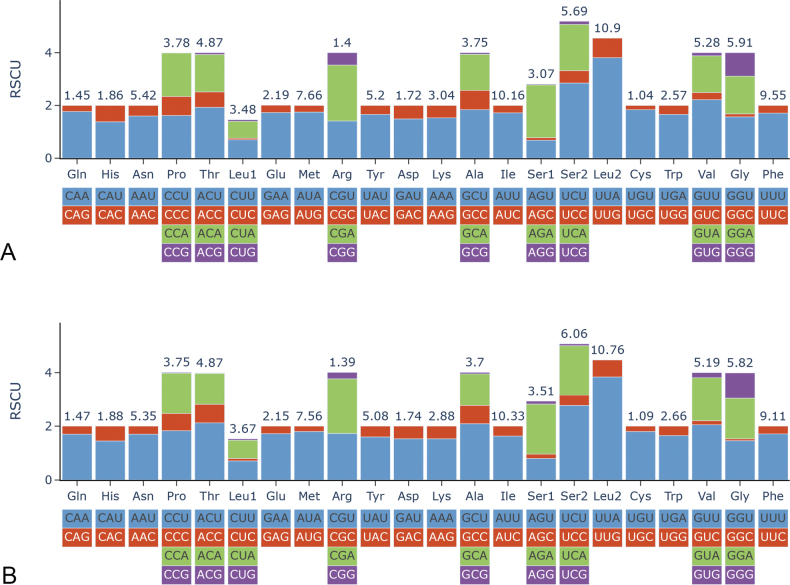 Figure 4
Figure 4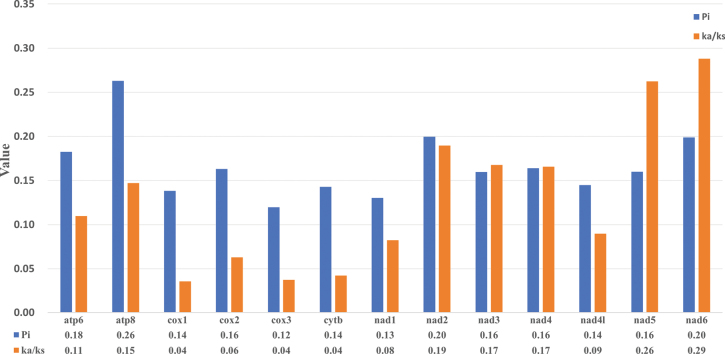 Figure 5
Figure 5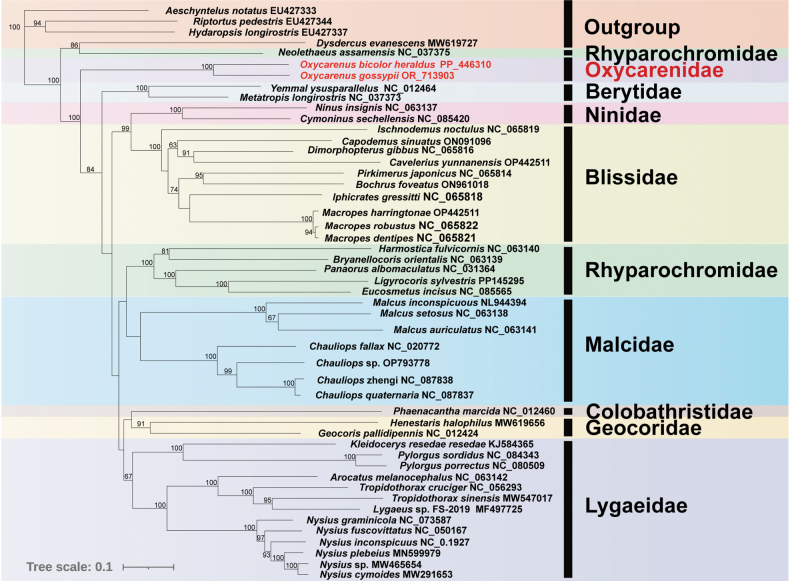 Figure 6
Figure 6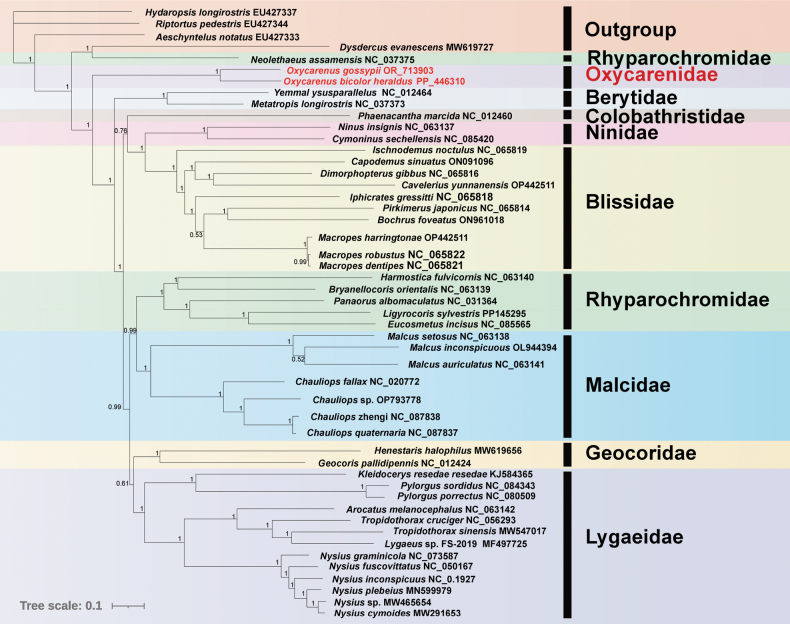 Figure 7
Figure 7| Superfamily | Family | Species | Length (bp) | GenBank No. |
|---|---|---|---|---|
|
|
| 15,744 |
| |
|
| 15,747 |
| ||
|
| 14,738 |
| ||
|
| 15,199 |
| ||
|
| 15,330 |
| ||
|
| 14,988 |
| ||
|
| 15,288 |
| ||
|
| 15,291 |
| ||
|
| 14,923 |
| ||
|
| 15,314 |
| ||
|
| 15,041 |
| ||
|
| 14,540 |
| ||
|
| 14,592 |
| ||
|
| 14,868 |
| ||
|
| 15409 |
| ||
|
| 15,332 |
| ||
|
| 14,688 |
| ||
|
| 15,235 |
| ||
|
| 16,301 |
| ||
|
| 14,575 |
| ||
|
| 16760 |
| ||
|
| 17,367 |
| ||
|
| 16,330 |
| ||
|
| 15,174 |
| ||
|
| 15,399 |
| ||
|
| 15,781 |
| ||
|
| 15,422 |
| ||
|
| 15,739 |
| ||
|
| 15300 |
| ||
|
| 15612 |
| ||
|
| 15507 |
| ||
|
| 15,097 |
| ||
|
| 15,316 |
| ||
|
| 14,894 |
| ||
|
| 15,962 |
| ||
|
| 14,632 |
| ||
|
| 16,144 |
| ||
|
| 15,462 |
| ||
|
| 15,606 |
| ||
|
| 14,562 |
| ||
|
| 15,703 |
| ||
|
| 16,621 |
| ||
|
| 15,067 |
| ||
|
| 16,345 |
| ||
|
|
| 15,635 |
| |
|
|
| 17,191 |
| |
|
| 16,521 |
| ||
|
| 14,532 |
|
| Name | Direction | Position From | Position To | Length (bp) | Intergenic nucleotides | Start/Stop Codons |
|---|---|---|---|---|---|---|
|
| J | 1 | 62 | 62 | 3 | |
|
| N | 60 | 128 | 69 | -1 | |
|
| J | 130 | 197 | 68 | 0 | |
|
| J | 198 | 1187 | 990 | 2 | ATA/TAA |
|
| J | 1186 | 1248 | 63 | 8 | |
|
| N | 1241 | 1302 | 62 | -1 | |
|
| N | 1304 | 1364 | 62 | -1 | |
|
| J | 1366 | 2899 | 1534 | 0 | TTG/T - - |
|
| J | 2900 | 2964 | 65 | 0 | |
|
| J | 2965 | 3640 | 676 | 0 | ATA/T - - |
|
| J | 3641 | 3711 | 71 | 0 | |
|
| J | 3712 | 3777 | 66 | 0 | |
|
| J | 3778 | 3936 | 159 | 7 | ATT/TAA |
|
| J | 3930 | 4595 | 666 | 1 | ATG/TAA |
|
| J | 4595 | 5381 | 787 | 0 | ATG/TAA |
|
| J | 5382 | 5447 | 66 | 0 | |
|
| J | 5448 | 5801 | 354 | 0 | ATA/TAA |
|
| J | 5802 | 5864 | 63 | 0 | |
|
| J | 5865 | 5927 | 63 | 0 | |
|
| J | 5928 | 5995 | 68 | 1 | |
|
| J | 5995 | 6063 | 69 | 1 | |
|
| J | 6063 | 6127 | 65 | 0 | |
|
| N | 6128 | 6190 | 63 | 1 | |
|
| N | 6190 | 7899 | 1710 | -3 | ATA/TAA |
|
| N | 7903 | 7964 | 62 | -2 | |
|
| N | 7967 | 9286 | 1320 | 7 | ATG/TAA |
|
| N | 9280 | 9558 | 279 | -5 | ATA/TAA |
|
| J | 9564 | 9625 | 62 | 0 | |
|
| N | 9626 | 9684 | 59 | 4 | |
|
| J | 9781 | 10236 | 456 | 1 | ATA/TAA |
|
| J | 10236 | 11370 | 1135 | 0 | ATG/T - - |
|
| J | 11371 | 11439 | 69 | -16 | |
|
| N | 11456 | 12379 | 924 | 0 | ATT/TAA |
|
| N | 12380 | 12445 | 66 | 0 | |
|
| N | 12464 | 13671 | 1208 | -22 | |
|
| N | 13694 | 13690 | 67 | -4 | |
|
| N | 13765 | 14372 | 608 | 0 |
| Name | Direction | Position From | Position To | Length (bp) | Intergenic nucleotides | Start/Stop Codons |
|---|---|---|---|---|---|---|
|
| J | 1 | 62 | 62 | 3 | |
|
| N | 60 | 128 | 69 | 1 | |
|
| J | 128 | 195 | 68 | 0 | |
|
| J | 196 | 1183 | 988 | 1 | ATA/TAA |
|
| J | 1185 | 1246 | 62 | 8 | |
|
| N | 1239 | 1300 | 62 | 0 | |
|
| N | 1301 | 1363 | 63 | -1 | |
|
| J | 1365 | 2898 | 1581 | 0 | TTG/T - - |
|
| J | 2899 | 2963 | 65 | 0 | |
|
| J | 2964 | 3639 | 699 | 0 | ATA/T - - |
|
| J | 3640 | 3711 | 73 | 0 | |
|
| J | 3712 | 3774 | 63 | 0 | |
|
| J | 3775 | 3933 | 159 | 7 | ATA/TAA |
|
| J | 3927 | 4592 | 666 | 1 | ATG/TAA |
|
| J | 4592 | 5378 | 790 | 0 | ATG/T - - |
|
| J | 5379 | 5443 | 65 | 0 | |
|
| J | 5444 | 5795 | 354 | -1 | ATT/TAG |
|
| J | 5797 | 5859 | 63 | 0 | |
|
| J | 5860 | 5922 | 65 | -1 | |
|
| J | 5924 | 5989 | 66 | 1 | |
|
| J | 5989 | 6057 | 69 | 1 | |
|
| J | 6057 | 6122 | 65 | 0 | |
|
| N | 6123 | 6187 | 63 | 20 | |
|
| N | 6168 | 7922 | 1714 | 24 | ATA/TAA |
|
| N | 7899 | 7960 | 70 | -2 | |
|
| N | 7963 | 9282 | 1320 | 7 | ATG/TAA |
|
| N | 9276 | 9557 | 282 | -2 | ATT/TAA |
|
| J | 9560 | 9621 | 62 | 0 | |
|
| N | 9622 | 9684 | 63 | 3 | |
|
| J | 9692 | 10153 | 462 | 1 | ATT/TAA |
|
| J | 10153 | 11289 | 1137 | 2 | ATG/TAG |
|
| J | 11288 | 11358 | 71 | -17 | |
|
| N | 11376 | 12298 | 960 | 0 | ATA/TAA |
|
| N | 12299 | 12365 | 67 | 0 | |
|
| N | 12366 | 13611 | 1253 | 0 | |
|
| N | 13612 | 13678 | 67 | -1 | |
|
| N | 13680 | 14453 | 802 | 0 |
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsPhytoplasmas and Hemiptera pathogens · Legume Nitrogen Fixing Symbiosis · Cocoa and Sweet Potato Agronomy
Introduction
Mitochondrial genome analysis is a powerful tool for elucidating the phylogeny and population genetics of insect taxa (Cameron and Whiting 2008; Li et al. 2023). Insects possess circular double-stranded mitochondrial molecules typically ranging from 14 to 20 kb in length. These genomes encode 37 genes, encompassing 13 protein-coding genes (PCGs), two ribosomal RNA genes (rRNAs), 22 transfer RNA genes (tRNAs), and a single control region. While insect mitochondria generally adhere to a conventional structure, there are exceptions, in certain species of Anoplura (Boore 1999; Shao et al. 2009). Characterized by compactness, insect mitochondrial genomes feature minimal spacer regions or overlapping sequences between adjacent genes (Boore 1999). Notably, they exhibit small size, stable genetic composition, relatively conserved gene sequences, rapid evolutionary rates, and comprehensive molecular information. Consequently, they serve as invaluable tools for investigating molecular evolution, phylogenetics, and population genetic structure (Xue and Bu 2008; Simon and Hadrys 2013; Kocher et al. 2014).
The Lygaeoidea, the second largest superfamily in Pentatomomorpha, comprises over 4,700 described species across 16 families (Weirauch and Schuh 2011; Dellapé and Henry 2024). This widespread terrestrial superfamily primarily includes herbivorous species feeding on plant seeds or sap, with some being economically significant pests (Sweet 2000). Among them, Oxycarenidae species predominantly inhabit the plants of Malvaceae and Sterculiaceae, where they feed on seeds and can inflict substantial damage to cotton and other mallow crops (Sureshan et al. 2021). Currently, over 140 species in 27 genera have been documented worldwide (Henry and Dellapé 2009; Xiao and Gao 2022; Dellapé and Henry 2024).
Past studies investigating the phylogenetic relationships of Pentatomomorpha have relied on morphological characters for classification (Henry 1997), and more recently, they have incorporated molecular data. The increasing number of Lygaeoidea species documented in recent years underscores the importance of exploring their phylogenetic relationships using mitochondrial DNA, both to validate previous findings and to provide additional insights.
In this study, we redescribe both O.gossypii Horváth, 1926 and Oxycarenusbicolorheraldus Distant, 1904. Additionally, two misidentifications in China are rectified, and the complete mitogenomes of these species are sequenced. Subsequently, we construct phylogenetic trees using the complete mitogenomes of 45 species of Lygaeoidea and four outgroup species. These findings contribute essential data for further investigations into the phylogenetic relationships within Lygaeoidea and Pentatomomorpha.
Materials and methods
Sample collection, identification and DNA extraction
Adult specimens of Oxycarenusbicolorheraldus Distant, 1904 were collected from Xiangshan Park, Pukou District, Nanjing, Jiangsu Province, China, in May 2020. Adult specimens of O.gossypii Horváth, 1926 were collected from Phoenix Airport, Sanya City, Hainan Province, China, in March 2020.
Composite images were obtained using an M205FA Leica stereomicroscope and camera, with the Leica Application Suite v. 4.5.0. Type label data are presented verbatim, with lines on the same label separated by a slash (/), and different labels divided by double slashes (//). Texts printed [pr] and handwritten [hw] are indicated. All measurements provided in the text are expressed in millimetres.
Abbreviations
BMNHNatural History Museum, London, United Kingdom;
IZASInstitute of Zoology, Academia Sinica, Beijing, China;
MSIE Shanghai Institute of Entomology, Shanghai, China;
NKUMInstitute of Entomology, Nankai University, Tianjin, China;
NJFUNanjing Forestry University, Nanjing, Jiangsu.
Genomic DNA were extracted from adult target insects using the Rapid Animal Genomic DNA Isolation Kit (Sangon Biotech, Shanghai, China).
Sequencing, assembly, annotation, and bioinformatics analyses
The mitochondrial genomes of these two species were sequenced using an Illumina MiSeq PE300 platform (Sangon Biotech, Shanghai, China). Subsequently, Fastp v. 0.36 (Chen et al. 2018) was employed to eliminate low-quality and short reads, ensuring the integrity of the data for subsequent analysis. SPAdes v. 3.15 (Bankevich et al. 2012) facilitated the de novo assembly of the high-quality next-generation sequencing data, resulting in the generation of contigs and scaffolds. Rigorous evaluation and quality control measures were applied to the assembly results using PrInSeS-G (Massouras et al. 2010). Potential contamination originating from the host genome was meticulously identified and eliminated, retaining only the scaffolds derived from the organelle genome. Sequence similarity was assessed by comparing the scaffolds with the NCBI library using BLASTn (Ye et al. 2012). Target scaffolds were manually selected based on sequencing depth and coverage information for each scaffold. GapFiller v. 1.11 (Boetzer and Pirovano 2012) was utilized to supplement and rectify obtained alleles by correcting editing errors and filling gaps introduced during splicing, including the insertion or deletion of fragments as needed.
The two mitogenome sequences were annotated using Geneious v. 11.0.2 (Kearse et al. 2012), following the invertebrate mitochondrial genetic code. Circular maps of the mitogenomes were generated using the CGView Server (Grant and Stothard 2008). To ensure annotation accuracy, all tRNA genes were verified using the MITOS Web Server (Bernt et al. 2013), and their secondary structures were predicted using the tRNAscan-SE Server v. 1.21 (Lowe and Chan 2016). PhyloSuite v. 1.2.3 (Xiang et al. 2023) and MEGA X (Kumar et al. 2018) were employed to determine base composition and relative synonymous codon usage (RSCU) values of the two mitogenome sequences. Non-synonymous substitutions (Ka) and synonymous substitutions (Ks) of the 13 PCGs of Oxycarenidae were calculated using DnaSP5 software (Librado and Rozas 2009), and Ka/Ks values were subsequently derived. Nucleotide composition skew was computed using the formulas developed by Perna and Kocher: AT-skew = (A − T) / (A + T) and GC-skew = (G − C) / (G + C). This study aimed to comprehensively examine the evolutionary patterns among mitochondrial protein-coding genes (PCGs) in species of Oxycarenidae.
Phylogenetic analysis
To investigate mitogenome arrangement patterns in Lygaeoidea, we compared the gene orders of all known Lygaeoidea mitogenomes with those of closely related taxa (Table 1). For phylogenetic analyses, we examined a total of 49 mitogenomes (Table 1), which included two newly generated sequences from this study. We standardized all sequences and extracted 13 PCGs using PhyloSuite v. 1.2.2 (Perna and Kocher 1995; Xiang et al. 2023). The 13 PCGs of these species were individually aligned using codon-based multiple alignments with MAFFT v. 7.313 software (Katoh and Standley 2013). The concatenated PCGs were processed with PhyloSuite v. 1.2.3. PartitionFinder2 selected optimal partitioning schemes and evolutionary models for constructing Bayesian-inference (BI) and maximum-likelihood (ML) trees with confidence (Soria-Carrasco et al. 2007; Lanfear et al. 2017). Phylogenetic trees were reconstructed using IQ-TREE v. 1.6.8 (Guindon et al. 2010) and MrBayes v. 3.2.6 (Ronquist et al. 2012) with the assistance of PhyloSuite v. 1.2.2.
Results
Taxonomy
Oxycarenus
gossypii
Taxon classificationAnimaliaHemipteraOxycarenidae
Horváth, 1926
7CE1A317-74E0-5474-BBF9-5053F24D28B7
Oxycarenus gossypii : Horváth 1926: 136; Esaki 1926: 161; Slater 1964: 673; Péricart 2001: 115. Oxycarenus laetus : Zheng and Zou 1981: 96. Misidentification.
Material examined.
China • 3♂♂1♀; Yunnan, Yuanjiang; alt. 400 m; 25 Jul. 2006; Weibing Zhu leg. (NKUM) • 1♂2♀♀; Yunnan, Xishuangbanna, Mengsong; alt. 1600 m; 23 Apr. 1958; Xvwu Meng leg. (IZAS) • 1♂; Yunnan, Xishuangbanna, Damenglong; alt. 650 m; 8 Apr. 1958; Leyi Zheng leg. (IZAS) • 5♂♂6♀♀; Hainan, Sanya, Fenghuang airport; 26 Mar. 2020; Bo Cai leg. (NJFU) • 16♂♂5♀♀; Hainan, Jianfengling thermal forestry institute; 21 Apr. 1985; Leyi Zheng leg.; from capsule of Abutilonindicum (NKUM) • 192♂♂183♀♀; Hainan, Sanya; alt. 10 m; 5–6 Apr. 1960; Suofu Li leg. (IZAS) • 26♂♂26♀♀; Hainan, Ledong; 11 Jun. 1960; Xuezhong Zhang leg. (IZAS) • 1♂; Hainan Nada; 27 Apr. 1954; Keren Huang leg. (IZAS).
Dorsal and ventral view of Oxycarenus species sequenced AO.gossypiiB, CO.bicolorheraldus.
Redescription.
Body brown, densely punctate, with white decumbent, erect, and apically enlarged setae. Antennae dark brown. Rostrum extends past anterior margin of abdominal sternite III, up to abdominal sternite V in females. Bucculae yellowish white. Pronotum brown, often lighter at anterior margin and posterior half, densely covered with deep, large punctures, white erect, and apically enlarged setae mixed with decumbent setae; callus area slightly elevated, densely covered with large, dark brown setae. Lateral margins of pronotum slightly sinuate. Scutellum brown, evenly punctate, flattened except basal margin concave, peripherally covered with both decumbent and erect, apically enlarged setae. Clavus brown, with both types of setae mentioned above. Corium yellowish brown, with a conspicuous black spot at distal angle; sparse erect setae, apical margin straight. Membrane smoky brown. Thoracic sternum brown, posterior margins of metapleura pale. Ostiolar peritreme of metathoracic scent gland yellow. Supracoxal lobewhite. Femora dark brown; fore femora beneath with four spines; fore tibiae yellowish brown; mid and hind tibiae pale, both ends brown. Abdominal sterna reddish brown, smooth, impunctate, without erect setae. Male sternites VI and VII with posterior margin with two transverse combs of glandular setae on either side of median line. Female abdominal sterna III to IV fused; ovipositor reaching abdominal sternites V–VII, with sternites V–VII medially strongly narrowed, pushed forward towards base of abdomen.
Pygophore: dorsal opening narrowly triangular (Fig. 2A); lateral projections in basal one third of pygophore openings, projecting obliquely posteriorly, tips truncate; distal margin of cup-like sclerite with a narrow, deep incision (Fig. 2A). Parameres (Fig. 2B, C) with basal shank relatively broad, about twice as wide as blade; outer projection rounded, inner projection more pointed from dorsal view; another finger-like inner projection present on inner side from ventral view.
Genitalia of Oxycarenus species A–CO.gossypiiA pygophore, posterodorsal view B, C left paramere, dorsal and ventral view D–FO.bicolorheraldusD pygophore, posterodorsal view E, F left paramere, dorsal and ventral view.
Measurements (in mm, n = 8). Body length 3.40–4.00, width 1.1–1.30. Head length 0.70–0.72, width across eyes 0.65–0.67; antennal segments I–IV length: 0.27–0.29: 0.56–0.58: 0.45–0.47: 0.52–0.54. Pronotum length 0.78–0.80, width of anterior margin 0.52–0.54, width of posterior margin 1.00–1.02; scutellum length 0.34–0.36, width 0.52–0.54. Distance of apex clavus–apex corium 0.60–0.62; distance of apex corium–apex membrane 0.72–0.74.
Distribution.
China (Hainan, Yunnan, Taiwan); Vietnam.
Remarks.
On review of descriptions and figures, we discovered that Oxycarenusgossypii was erroneously identified as Oxycarenuslaetus (Kirby, 1891) in the study by Zheng and Zou (1981). However, distinct differences exist between these species: the clavus appears brown in O.gossypii, whereas it is pale in O.laetus; the membrane presents a smoky golden-brown hue in O.gossypii (in contrast to the colorless and hyaline membrane of O.laetus); and, while the corium of O.gossypii is pale or slightly smoky except at the base, it consistently remains pale in O.laetus.
Oxycarenus
bicolor heraldus
Taxon classificationAnimaliaHemipteraOxycarenidae
Distant, 1904
8BDACA5D-4334-58B3-A37D-FEEF2B50885F
Oxycarenus heraldus : Distant 1904: 44. Oxycarenus bicolor variety heraldus : Bergroth 1918: 73. Oxycarenus bicolor heraldus : Slater 1964: 670. Oxycarenus lugubris : Zheng and Zou 1981: 97. Misidentification.
Type material examined
(digital photograph). Lectotype: Burma • ♀; Carin Chebà [pr] / 900–1100 m [pr] / L. Fea V XII-88 [pr] // heraldus [hw] / Dist. [hw] // Distant Coll. / 1911–383 // Type [pr, red] // SYN/ TYPE [pr, blue] // Oxycarenus / heraldus / Distant, 1904: 44 [pr] / BMNH(E) / 1340705 [pr] (BMNH).
Paralectotype: same information except: BMNH(E) / 1340706 [pr].
Other material examined.
China • 2♀♀; Gansu, Wen county, Fanba; 30 Jul. 1988; collected from capsule of Abutilontheophrasti (NKUM) • 20♂♂15♀♀; Jiangsu, Nanjing, Laoshan; 20 Jun. 2021; collected from capsule of Hibiscusmutabilis (NJFU) • 6♂♂7♀♀; Sichuan, Qingchengshan; 16 Aug. 1956; Leyi Zheng leg. (NKUM) • 20♂♂25♀♀; Yunnan, Dali, Cangshan; 19 Aug. 2006; Zhonghua Fan leg. (NKUM) • 300♂♂242♀♀; Yunnan, Menglong, Banna, Mengsong, alt. 1600 m; 23 Apr. 1958; Xvwu Meng leg. (IZAS) • 40♂♂32♀♀; Yunnan, Pingbian; alt. 1300 m; 22 Jun. 1956; Keren Huang leg. (MSIE).
Redescription.
Head dark, blackish brown or black, densely coarsely punctate, with white, flat, decumbent setae and sparser erect, apically enlarged, long setae. Antennae dark, blackish brown or black, with segment I extending to tip of clypeus. Head ventrally densely covered with silvery-white, flat setae. Rostrum extends to hind coxae or middle of abdominal sternite III. Bucculae dark. Pronotum brown with a black transverse stripe at callus area. Covered with coarse punctures and sparsely erect and apically enlarged long setae, with slightly sinuate lateral margins. Scutellum dark blackish brown or black, similar setae to pronutum, punctuated, with a sunken base and a slightly elevated middle. Clavus dark brown to blackish brown, possessing three lines of punctures, with middle row incomplete. Corium with exocorium, basal 1/3 of inner corium, and distal angle yellowish white, about middle 1/3 of inner corium blackish brown, not reaching exocorium; sometimes, extreme distal angles of corium slightly darkened, but not with obvious small black spots; distal margin of corium straight; clavus and corium with sparse pale erect setae. Membrane dark blackish brown, with basal margin adjoining distal margin of corium narrowly white. Head and prothorax ventrally densely covered with silvery-white, decumbent setae; thoracic sternites and pleurae black or dark blackish brown, glossy, except supracoxal lobe and posterior margin of metapleura pale; ostiolar peritreme of metathoracic scent gland strongly protruding, basally brown and distally yellow. Femora blackish brown, slightly thickened; fore femora with four spines; tibiae yellow with both ends dark blackish brown, and fore tibiae darker. Abdomen reddish brown to blackish brown. Posterior margin of sternites VI and VII in males with two conspicuous transverse combs of glandular setae on either side of median line. Female abdominal sterna III–IV fused; ovipositor reaching abdominal sternites V–VII, with sternites V–VII medially strongly narrowed and pushed forward towards abdominal sternites V.
Pygophore: dorsal opening broadly rounded; lateral projections finger-like, slightly inclined posteriorly and internally; distal margin of cup-like sclerite bifurcate (Fig. 2D). Parameres with blade falcate and curved; outer projection rounded; inner projection projecting dorsoventrally, more square (Fig. 2E, F).
Measurements (in mm, n = 8). Body length 3.80–4.30, width 1.10–1.40. Head length 0.71–0.73, width across eyes 0.72–0.73; antennal segments I–IV length: 0.28–0.30: 0.61–0.63: 0.47–0.49: 0.58–0.60. Pronotum length 0.83–0.85, width of anterior margin 0.58–0.60, width of posterior margin 1.10–1.11; scutellum length 0.41–0.43, width 0.54–0.55. Distance of apex clavus–apex corium 0.89–0.90; distance of apex corium–apex membrane 0.67–0.69.
Distribution.
China (Gansu, Jiangsu, Hubei, Sichuan, Yunnan); Burma.
Remarks.
The specific status of Oxycarenusheraldus Distant, 1904 was previously reduced to Oxycarenusbicolorvar.heraldus by Bergroth (1918), and later treated as subspecies Oxycarenusbicolorheraldus by Slater (1964).
Oxycarenusbicolorheraldus shares similar coloration with Oxycarenusbicolorbicolor, but there are notable differences. Unlike Oxycarenusbicolorbicolor, the brown spots on the hemelytra of Oxycarenusbicolorheraldus do not reach the exocorium (the brown spots on the hemelytra extend to the lateral margin of the corium in O.bicolorbicolor). Furthermore, the body size of O.bicolorheraldus is larger (3.80–4.30 mm) compared to O.bicolor (which is smaller, approximately 3.0–3.4 mm), and while the postero-lateral angles of the corium in O.bicolorheraldus may be slightly darkened, but they lack the distinct small black spots that are present in O.bicolorbicolor.
Oxycarenusbicolorheraldus is a common species in China, but it has long been misidentified as Oxycarenuslugubris (Motschulsky, 1859) (Zheng and Zou 1981). In comparison with O.lugubris, the pronotum of O.bicolorheraldus is brown with a black transverse stripe, whereas in O.lugubris, it is entirely black. Furthermore, only the middle 1/3 of the inner corium is blackish brown in O.bicolorheraldus, with the basal membrane narrowly white, while the distal 2/3 of the inner corium is entirely black, and the base of the membrane is also black in O.lugubris. Although both the species are distributed in China, O.lugubris has only been recorded from Taiwan and Hong Kong according to the data available on the iNaturalist website.
Genome structure and base composition
We have sequenced and annotated the complete mitogenomes of O.gossypii and O.bicolorheraldus, which were 16,144 bp and 15,462 bp in length, respectively (Table 1). These mitogenome sequences consist of the 37 typical insect mitochondrial genes, including 13 protein-coding genes (PCGs), 22 transfer RNA genes (tRNAs), and two ribosomal RNA genes (rRNAs), along with an AT-rich region known as the control region (CR), forming a double-stranded ring structure (Fig. 3). The N-strand encodes 14 genes, while the J-strand encodes 23 genes, consistent with the mitochondrial gene arrangement observed in known Lygaeoidea species and the classical insect Drosophilayakuba (Burla, 1954) (Clary and Wolstenholme 1985; Hua et al. 2008; Küechler et al. 2010; Cao et al. 2020).
Circular map of the complete mitogenome of Oxycarenus species AO.gossypiiBO.bicolorheraldus. Different colors indicate different types of genes and regions. Genes in the outer circle are located on the J-strand, and genes in the inner circle are located on the N-strand.
The nucleotide composition of the O.gossypii mitogenome was as follows: A = 41.35%, T = 32.82%, C = 15.33%, and G = 10.50%, while that of O.bicolorheraldus was A = 40.86%, T = 33.11%, C = 15.68%, and G = 10.35%. Both mitogenomes exhibited a high AT content, with O.gossypii at 74.17% and O.bicolorheraldus at 73.97%. Additionally, both mitogenomes displayed a slightly positive AT-skew (0.11 and 0.10) and a negative GC-skew (−0.18 and −0.20), indicating a bias towards A and T nucleotides. The study identified 15 gaps in the two mitogenome sequences, ranging from 1 bp to 22 bp, with the longest intergenic spacer being 22 bp, found between rrnL and trnV in O.gossypii (Table 2). Moreover, there were 25 overlapping gene regions, with lengths ranging from 1 bp to 24 bp, and the longest overlap of 24 bp was observed between nad5 and trnH in O.bicolorheraldus (Table 3).
Protein-coding genes
The concatenated length of the 13 protein-coding genes (PCGs) of O.gossypii was 10,990 bp, encoding 3,663 amino acid residues. Similarly, the concatenated length of the 13 PCGs of O.bicolorheraldus was 11,112 bp, encoding 3,702 amino acids. Both species share the same arrangement in their mitochondrial genomes. The majority of PCGs initiate translation using the start codon ATN, except for cox1, which starts with TTG. There are three types of stop codons: TAA, TAG, and an incomplete stop codon T that is completed by the addition of 3′A residues to the mRNA.
The Relative Synonymous Codon Usage (RSCU) of the two Oxycarenidae species was computed and depicted in Fig. 4. Among the codons utilized, CGA-Arg, GCU-Ala, UCU-Ser, UUA-Leu, and GUU-Val were the most frequently employed. Particularly, UUA emerged as the most preferred codon. Moreover, a pronounced bias toward A/T nucleotides was evident across the Protein-Coding Genes (PCGs). Nucleotide diversity (Pi) and the ratios of Ka/Ks for the two species were calculated based on the 13 PCGs, as illustrated in Fig. 5. Pi values ranged from 0.12 to 0.26, with the highest values observed in atp8 and the lowest in cox3, underscoring cox3’s role as the most conserved gene in Oxycarenidae. All Ka/Ks ratios were below 1, varying from 0.04 to 0.29, indicative of purifying selection acting on the genes. Particularly noteworthy was nad6’s highest Ka/Ks values, suggesting rapid evolution, while cox1 and cytb exhibited the slowest evolution, with the lowest values.
RSCU values of Oxycarenus species AO.gossypiiBO.bicolorheraldus. The ordinate represents the RSCU (the number of times a certain synonymous codon is used/the average number of times that all codons encoding the amino acid are used). The abscissa represents different amino acids. The number above the bar graph represents the ratio of amino acids (number of certain amino acids/total number of all amino acids).
Nucleotide diversity (Pi) and nonsynonymous (Ka)/synonymous (Ks) mutation rate ratios of 13 PCGs of Oxycarenidae species (the Pi and Ka/Ks values of each PCG are shown under the gene name).
RNA
The rRNA genes were positioned between the AT-rich region and trnL1, separated by trnV. Their total length ranged from 1816 bp to 1840 bp. In both species, the collective length of the 22 tRNA genes was 1433 bp, with individual tRNA genes varying from 61 bp to 71 bp. Notably, eight tRNA genes were encoded on the N-strand, while the remaining 14 genes were encoded on the J-strand, consistent with previous findings (Bernt et al. 2013; Cao et al. 2020).
Most tRNA genes exhibited a typical cloverleaf secondary structure, featuring a TΨC arm, an amino acid acceptor arm, an anticodon arm, and a dihydrouridine arm. However, an exception was observed in trnS1, where the dihydrouridine arm was absent in O.gossypii, forming a loop. Additionally, trnS1 of O.bicolorheraldus displayed an atypical cloverleaf structure, as depicted in Suppl. material 1, a pattern also observed in other species (Zhao et al. 2018).
Phylogenetic analysis
Phylogenetic relationships within Lygaeoidea were elucidated through the reconstruction of mitochondrial 13 PCGs using both BI and ML methods (Figs 6, 7). A total of 45 Lygaeoidea species were selected as the ingroup, with four species from Coreoidea and Pyrrhocoroidea serving as the outgroup. The resulting ML and BI trees exhibited slightly different topologies. Most families were consistently identified as monophyletic, except for Rhyparochromidae, which was paraphyletic. Dysdercusevanescens (Pyrrhocoroidea: Pyrrhocoridae) and Neolethaeusassamensis (Lygaeoidea: Rhyparochromidae) clustered together in both ML and BI trees (Figs 6, 7). The position of Colobathristidae proved to be unstable in the phylogenetic trees. In one instance, it clustered with Geocoridae with relatively low nodal support (Fig. 6), while another result indicated that Colobathristidae, Ninidae, and Blissidae formed a monophyletic group (Fig. 7). Furthermore, the two sequenced species of Oxycarenidae formed a single clade with a high support value.
Phylogenetic tree of Lygaeoidea inferred from ML based on 13 PCGs. The numbers on the branches show bootstrap values (values >60% are shown). Two Oxycarenidae species in this study are marked in red.
Phylogenetic tree of Lygaeoidea inferred from BI based on 13 PCGs. The numbers on the branches show posterior probabilities (values >0.50 are shown). Two Oxycarenidae species in this study are marked in red.
Discussion and conclusion
In this study, we redescribed two Oxycarenidae species: Oxycarenusgossypii and O.bicolorheraldus. We also detected misidentifications of two species in China. However, the sheer abundance and morphological similarities amongst oxycarenid species present challenges to providing an accurate morphology alone based classification.
The mitochondrial genomes of O.gossypii and O.bicolorheraldus were sequenced and analyzed, revealing a shared structural similarity. Both genomes exhibited a typical double-stranded ring structure housing 37 genes, including a non-coding control region. Remarkably, neither genome displayed any gene rearrangement, consistent with known genomic arrangements (Ding et al. 2023). The AT content significantly outweighed the CG content, showing a strong AT bias, a trait observed across various families within Pentatomomorpha (Guo and Yuan 2016). Our analysis of relative synonymous codon usage unveiled a prevalent preference for A/T codons, particularly at the termini of protein-coding genes, a phenomenon observed across all sequenced Pentatomomorpha (Hassanin et al. 2005; Guo and Yuan 2016). This nucleotide composition bias is believed to stem from a combination of mutational pressure and natural selection. The KA/KS analysis identified cox1 and cytb as the most conserved genes, whereas nad6 exhibited relatively higher evolutionary rates. Most protein-coding genes initiated translation using the start codon ATN, with the exception of cox1 (TTG). Additionally, three types of stop codons were identified: TAA, TAG, and an incomplete stop codon T. While most tRNA molecules exhibited a typical cloverleaf structure, trnS1 displayed an atypical cloverleaf structure in both species.
The monophyly of most families within Lygaeoidea was strongly supported, except for Rhyparochromidae, marking a deviation from Henry’s findings (1997). Neolethaeusassamensis (Lygaeoidea: Rhyparochromidae) clustering with Dysdercusevanescens (Pyrrhocoroidea: Pyrrhocoridae) in both ML and BI trees mirrored Gao and Dong’s (2023) results. The branches of Ischnorhynchinae, Lygaeinae, and Orsillinae formed a cohesive group designated as Lygaeidae, aligning with Gao and Dong’s findings (Gao and Dong 2023). The sister group relationship between Henestarishalophilus and Geocorispallidipennis supported Henry’s (1997) earlier assertion. However, the phylogenetic position of Colobathristidae remained unstable in our PCG-based tree, in contrast to Ye et al.’s (2022) findings. Moreover, our results did not support the hypothesis that Colobathristidae and Berytidae formed sister groups, nor did they form the “malcid line” with Malcidae and Cymidae as proposed by Henry (1997). The formation of a monophyletic group by Blissidae and Ninidae, excluding Berytidae, diverged from the inferred relationship based on 18S rRNA (Xie et al. 2005). However, our examination validated the hypothesis that the two sequenced Oxycarenidae species constituted a single clade, with the position of Oxycarenidae remaining stable in both ML and BI phylogenetic trees. While our findings enrich the structural information of mitochondrial genomes, a comprehensive discussion on the phylogenetic relationships within Lygaeoidea remains challenging. For a deeper understanding of their evolutionary history, it is imperative that more Lygaeoidea species are sequenced in future studies.
Supplementary Material
XML Treatment for Oxycarenus gossypii
XML Treatment for Oxycarenus bicolor heraldus
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Bankevich A Nurk S Antipov D Gurevich AA Dvorkin M Kulikov AS Lesin VM Nikolenko SI Pham S Prjibelski AD Pyshkin AV Sirotkin AV Vyahhi N Tesler G Alekseyev MA Pevzner PA (2012) SP Ades: a new genome assembly algorithm and its applications to single–cell sequencing.Journal of Computational Biology 19(5): 455–477. 10.1089/cmb.2012.002122506599 PMC 3342519 · doi ↗ · pubmed ↗
- 2Bergroth E (1918) Studies in Philippine Heteroptera, I.Philippine Journal of Science 13(2): 75–126.
- 3Bernt M Donath AJühling F Externbrink F Florentz C Fritzsch GPütz J Middendorf M Stadler PF (2013) MITOS: improved de novo metazoan mitochondrial genome annotation.Molecular Phylogenetics and Evolution 69(2): 313–319. 10.1016/j.ympev.2012.08.02322982435 · doi ↗ · pubmed ↗
- 4Boetzer M Pirovano W (2012) Toward almost closed genomes with Gap Filler. Genome Biology 13(6): R 56. 10.1186/gb-2012-13-6-r 56PMC 344632222731987 · doi ↗ · pubmed ↗
- 5Boore JL (1999) Animal mitochondrial genomes.Nucleic acids research 27(8): 1767–1780. 10.1093/nar/27.8.176710101183 PMC 148383 · doi ↗ · pubmed ↗
- 6Cameron SL Whiting MF (2008) The complete mitochondrial genome of the tobacco hornworm, Manducasexta, (Insecta: Lepidoptera: Sphingidae), and an examination of mitochondrial gene variability within butterflies and moths.Gene 408(1–2): 112–123. 10.1016/j.gene.2007.10.02318065166 · doi ↗ · pubmed ↗
- 7Cao Y Wu HT Li M Chen WT Yuan ML (2020) The complete mitochondrial genome of Nysiusfuscovittatus (Hemiptera: Lygaeidae).Mitochondrial DNA Part B 5(3): 3483–3484. 10.1080/23802359.2020.182706233458212 PMC 7783115 · doi ↗ · pubmed ↗
- 8Chen SF Zhou YQ Chen YR Gu J (2018) Fastp: an ultra-fast all-in-one FASTQ preprocessor.Bioinformatics 34(17): 884–890. 10.1093/bioinformatics/bty 56030423086 PMC 6129281 · doi ↗ · pubmed ↗