Endogenous PGD2 acting on DP2 receptor counter regulates Schistosoma mansoni infection-driven hepatic granulomatous fibrosis
Giovanna N. Pezzella-Ferreira, Camila R. R. Pão, Isaac Bellas, Tatiana Luna-Gomes, Valdirene S. Muniz, Ligia A. Paiva, Natalia R. T. Amorim, Claudio Canetti, Patricia T. Bozza, Bruno L. Diaz, Christianne Bandeira-Melo, Dominique Soldati-Favre, Meera Goh Nair

TL;DR
This study shows that PGD2 helps protect the liver from fibrosis in schistosomiasis, and blocking it may worsen the disease.
Contribution
The study reveals an unexpected protective role of PGD2 in schistosomiasis-induced liver fibrosis through DP2 receptor activation on eosinophils.
Findings
Inhibiting PGD2 or its DP2 receptor increased liver fibrosis and pro-fibrotic cytokines in infected mice.
PGD2/DP2 signaling stimulates anti-fibrogenic cysLTs production by eosinophils, which down-regulates fibrosis.
Eosinophils are the main source of LTC4 in PGD2-mediated schistosomiasis, and their depletion reduces cysLTs.
Abstract
Identifying new molecular therapies targeted at the severe hepatic fibrosis associated with the granulomatous immune response to Schistosoma mansoni infection is essential to reduce fibrosis-related morbidity/mortality in schistosomiasis. In vitro cell activation studies suggested the lipid molecule prostaglandin D2 (PGD2) as a potential pro-fibrotic candidate in schistosomal context, although corroboratory in vivo evidence is still lacking. Here, to investigate the role of PGD2 and its cognate receptor DP2 in vivo, impairment of PGD2 synthesis by HQL-79 (an inhibitor of the H-PGD synthase) or DP2 receptor inhibition by CAY10471 (a selective DP2 antagonist) were used against the fibrotic response of hepatic eosinophilic granulomas of S. mansoni infection in mice. Although studies have postulated PGD2 as a fibrogenic molecule, HQL-79 and CAY10471 amplified, rather than attenuated, the…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
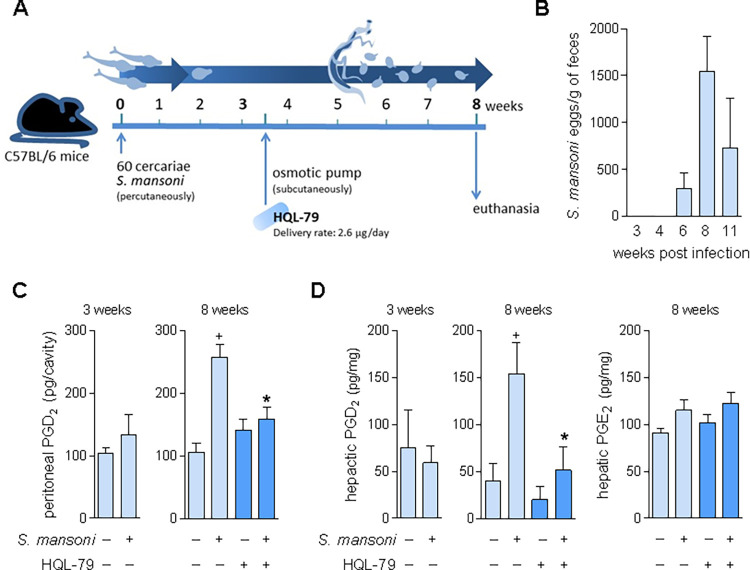 Fig 1
Fig 1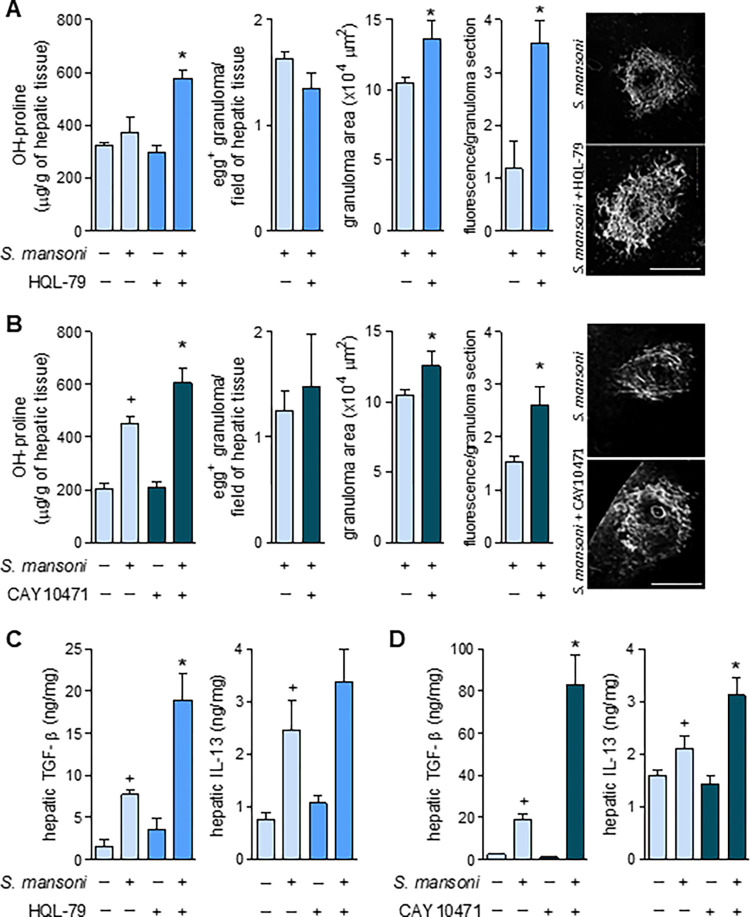 Fig 2
Fig 2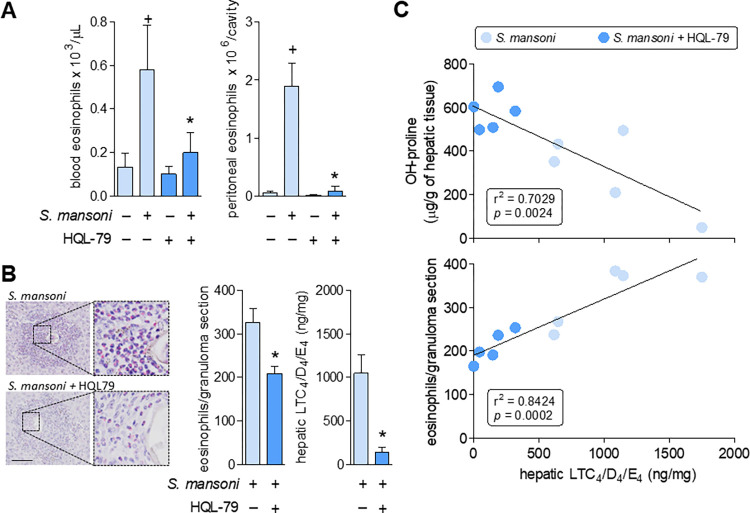 Fig 3
Fig 3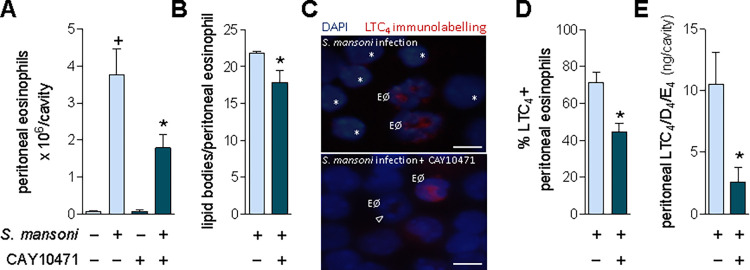 Fig 4
Fig 4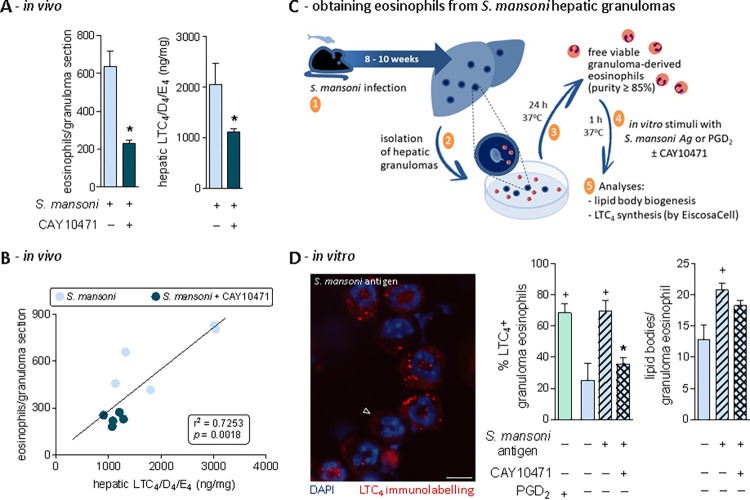 Fig 5
Fig 5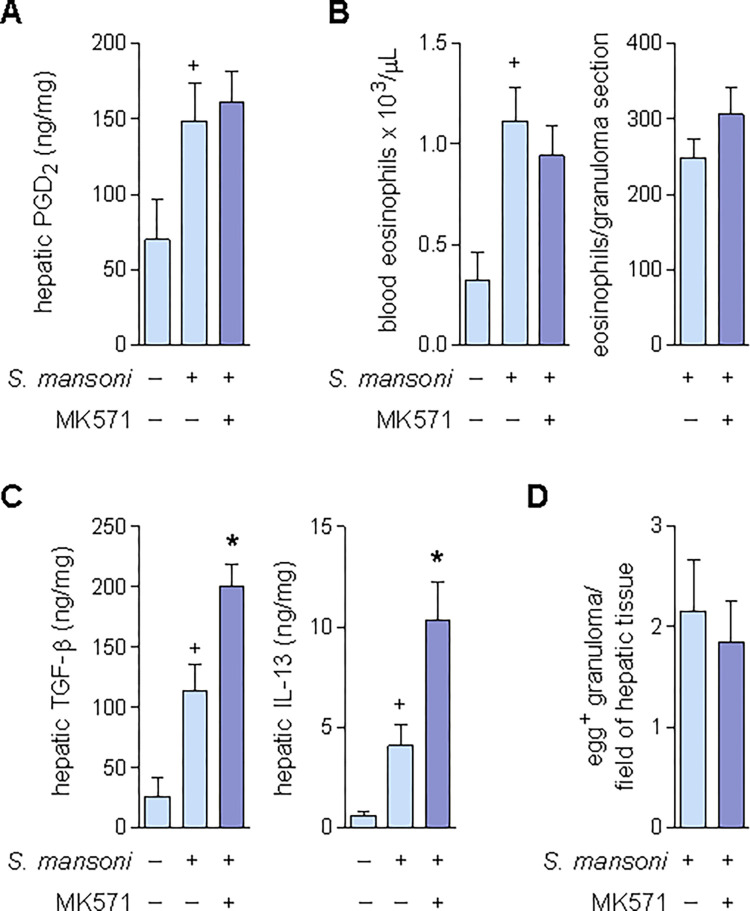 Fig 6
Fig 6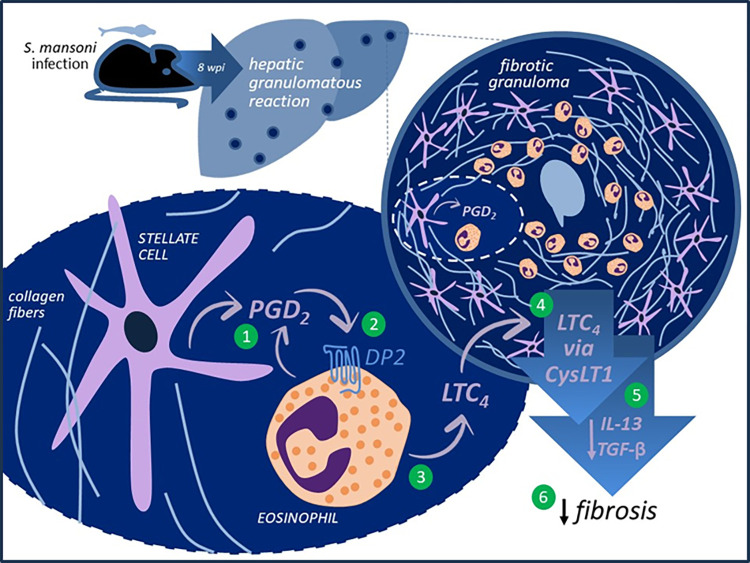 Fig 7
Fig 7- —http://dx.doi.org/10.13039/501100003593Conselho Nacional de Desenvolvimento Científico e Tecnológico
- —http://dx.doi.org/10.13039/501100003593Conselho Nacional de Desenvolvimento Científico e Tecnológico
- —http://dx.doi.org/10.13039/501100004586Fundação Carlos Chagas Filho de Amparo à Pesquisa do Estado do Rio de Janeiro
- —http://dx.doi.org/10.13039/501100004586Fundação Carlos Chagas Filho de Amparo à Pesquisa do Estado do Rio de Janeiro
- —http://dx.doi.org/10.13039/501100004586Fundação Carlos Chagas Filho de Amparo à Pesquisa do Estado do Rio de Janeiro
- —http://dx.doi.org/10.13039/501100004586Fundação Carlos Chagas Filho de Amparo à Pesquisa do Estado do Rio de Janeiro
- —http://dx.doi.org/10.13039/501100002322Coordenação de Aperfeiçoamento de Pessoal de Nível Superior
- —http://dx.doi.org/10.13039/501100002322Coordenação de Aperfeiçoamento de Pessoal de Nível Superior
- —http://dx.doi.org/10.13039/501100003593Conselho Nacional de Desenvolvimento Científico e Tecnológico
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsParasites and Host Interactions · Drug Transport and Resistance Mechanisms · Pneumocystis jirovecii pneumonia detection and treatment
Introduction
Often resulting in portal hypertension and liver failure, liver fibrosis is a collateral outcome of hepatic diseases of varying etiologies, such as schistosomiasis — a neglected tropical immunopathology, whose fibrosis-driven morbidity and mortality affects millions of people infected with Schistosoma spp parasites worldwide. Current anti-schistosomal chemotherapy effectively kills worms resolving the infection with only mild side effects. Yet, controversy persists about whether the present therapeutic approach successfully treats the schistosomiasis-induced liver fibrotic sequelae [1–3].
In Schistosoma mansoni (S. mansoni) infection-driven pathogenesis, hepatic fibrosis is a long-lasting feature of the excessive granulomatous inflammation built around liver-trapped eggs. The assembly of S. mansoni-induced hepatic fibrotic granulomas follows a switch from an initial type 1 to the pro-fibrotic type 2 immune response. And though protective at its onset, the large extensions occupied by the persistent fibrotic granulomatous tissue can culminate with disruption of the normal liver architecture and loss of its essential functions. Initiated by the egg deposition in the liver parenchyma and orchestrated by type 2 cytokines and intense eosinophilic inflammation, the hepatocellular response within schistosomal granulomatous space triggers complex cellular events — remarkably, the transdifferentiation of resident stellate cells into hepatic myofibroblasts [4]. Intra-granulomatous activation of stellate myofibroblast evokes prominent collagen synthesis and periovular deposition in the S. mansoni-infected liver parenchyma [5]. It is well-established that the granulomatous myofibroblast-ruled fibrogenic process is notably promoted by transforming growth factor-β1 (TGF-β1) and IL-13; indeed, the specific blockade of TGF-β1 and IL-13 reduces the schistosomal fibrosis [6,7]. Of note, besides their role in fibrosis as the pivotal collagen-synthesizing cells, S. mansoni infection-activated stellate myofibroblasts also appear to display additional immunomodulatory functions through the release of chemokines, cytokines, and bioactive lipid mediators, such as prostaglandin (PG)D_2_ [8].
PGD_2_ is a downstream metabolite of the arachidonic acid/cyclooxygenase (COX) enzymatic pathway, which is synthesized by the rate-limiting PGD synthase (PGDS) enzymes. In S. mansoni mammal hosts, in addition to the lipocalin-type PGDS highly expressed in the central nervous system, the hematopoietic PGD synthase (hPGDS) is the main terminal enzyme for PGD_2_ synthesis in peripheral tissues [9]. Besides eosinophils and various inflammatory cell types, hepatic stellate myofibroblasts of the S. mansoni-driven granuloma express hPGDS and are notable sources of PGD_2_ [8–11].
Either autocrinally or paracrinally, PGD_2_ exerts most of its effects through the activation of two distinct 7-transmembrane G-protein coupled receptors, DP1 and DP2, co-expressed in cell surfaces. In contrast to the adenylate cyclase-driven inhibitory feature of DP1 activation, DP2 (formerly known as CRTH2) elicits the distinctive downstream signaling classically displayed by chemoattractant receptors. The resultant of concomitant activating both PGD_2_ receptors is a cell type-specific phenomenon [12–14]. In eosinophils, for induction of lipid body-compartmentalized leukotriene (LT)C_4_ synthesis, PGD_2_-induced concurrent DP1/DP2 activation converges toward triggering cooperative signaling and intracellular events [12,15,16]: DP1 brings about the assembly of new active lipid body compartments and DP2 activates LTC_4_ synthesizing machinery within newly formed lipid bodies [15,17].
The link between PGD_2_ and schistosomal immunopathology began 40 years ago with the observation that S. mansoni cercariae can generate PGD_2_ by themselves [18] and has advanced to unveil that (i) not only cercariae but also other S. mansoni life stages (remarkably eggs) produce PGD_2_ via the enzymatic activity of a schistosomal PGDS — the Sm28GST, a 28 kDa glutathione-S-transferase [19–21]; (ii) cercariae-derived PGD_2_ within infected skin elicits cutaneous immune evasion, decreasing the migratory capability of Langerhans cells towards draining lymph nodes [22]; and (iii) by employing a DP1 receptor-deficient mouse, activation of host DP1 receptor during S. mansoni infection was shown to be crucial for the establishment of early type 1 immune response (until 1 week) as well as late worm burden (7 weeks) in the liver [19]. Altogether, the data clearly show PGD_2_’s role in schistosomal immunopathogenesis and its potential as a novel adjuvant target for the current chemotherapy, focusing on quelling the deleterious impact of parasitic infection on the host [23].
Understanding the role of PGD_2_ after the host has been parasitized and before liver fibrosis has fully developed may generate medically relevant insight into disease management. Such studies are particularly relevant due to some divergent evidence for the role of PGD_2_ in fibrotic processes that affect other organs’ fibrogenesis. In renal settings, while in vitro PGD_2_ played an anti-fibrogenic role by suppressing the induction of fibrotic phenotype in cultured kidney cells [24,25]; in vivo, PGD_2_ emerged as pro-fibrogenic since DP2 antagonism downregulates renal fibrosis in a chronic model of kidney inflammation [26]. In the lung, while studies on non-asthmatic models of pulmonary fibrosis acknowledge PGD_2_ itself (via its DP2 receptor) as either a beneficial [27–30] or a deleterious [31] signal against fibrotic development; in the asthmatic lung, however, PGD_2_/DP2 displays undisputable pro-fibrogenic impact [32,33]. Although simple extrapolation of PGD_2_’s role in liver fibrosis based on evidence gathered on different tissues may not be straightforward, it is reasonable to speculate that schistosomiasis-driven fibrosis may be promoted by PGD_2_, in a similar pro-fibrotic fashion as it is in asthma due to the shared type 2-governed eosinophilic environment.
Since PGD_2_ elusive function on fibrogenesis has not been properly validated using in vivo models of hepatic inflammation, the potential link between PGD_2_ and schistosomal hepatic fibrosis is mostly drawn from in vitro studies. While by employing DP1 deficient mice, no impact was observed in the fibrotic feature of hepatic granulomatous reactions around S. mansoni eggs [19], it has been observed that (i) host PGD_2_ synthesized by S. mansoni-driven stellate myofibroblasts in response to in vitro TGF-β autocrinally mediates myofibroblastic secretory activity [34]; (ii) S. mansoni-derived PGD_2_ acting via DP1 triggers in vitro eosinophil activation (lipid body biogenesis) [21] and (iii) induces 15-lypoxygenase-driven synthesis of eoxin C_4_, which in turn promotes TGF-β secretion [21]. Taken together with the pro-fibrotic role in type 2-biased asthma, these pieces of evidence build a case for a PGD_2_ role in promoting S. mansoni-driven fibrogenesis in hepatic granulomas. Moreover, such findings support recommendations of PGD_2_ as a possible target for anti-fibrotic therapies in schistosomiasis [35–37].
Our investigation of the role of PGD_2_ in schistosomal granulomas led to the discovery of unanticipated endogenous counter-regulatory functions required to limit hepatic fibrosis beyond those associated with DP1 receptor activation [19,38,39]. We have unveiled a PGD_2_/DP2 modulatory mechanism that reduces the granuloma-associated fibrotic reaction elicited by S. mansoni infection. Although missing pieces of information do not allow the drawing of a complete picture of the events responsible for PGD_2_-mediated inhibition of S. mansoni infection-elicited hepatic granulomatous fibrosis, our findings unveil that once oviposition starts, PGD_2_ (i) is produced within hepatic granuloma, seemingly, by hPGDS-expressing stellate cells; (ii) activates DP2 receptors to mobilize blood eosinophils to the hepatic granulomas; and (iii) stimulates granuloma-infiltrating eosinophils to synthesize and secrete LTC_4_; which, via CysLT1 receptor activation (iv) reduces hepatic production of fibrogenic TGF-β and IL-13; (v) decreasing synthesis and deposition of collagen around granuloma-trapped eggs.
Results and discussion
S. mansoni infection-induced hepatic granuloma formation triggers hPGDS-driven PGD2 synthesis
In vitro studies have suggested PGD_2_ as a potential molecular target for novel anti-fibrotic therapies in the hepatic granulomatous pathology of S. mansoni infection [8,35,37]. To address such inference from in vitro data, here we first evaluate whether enhanced PGD_2_ production is elicited in an in vivo experimental model of schistosomiasis — C57BL/6 mice infected percutaneously with 60 cercariae of S. mansoni (Fig 1A). As shown in Fig 1B, the infection protocol triggers productive oviposition first detected in feces within 6 weeks, which peaks within 8 weeks post-infection (wpi), while no S. mansoni egg was found in feces as early as 3 weeks. Before oviposition (3 wpi), no increase in PGD_2_ synthesis was detected within peritoneal or hepatic compartments of S. *mansoni-*infected mice (Fig 1C and 1D). However, later at 8 wpi, PGD_2_ levels were elevated in both egg-containing granulomatous liver tissue, as well as, in peritoneal cavities of S. mansoni-infected mice (Fig 1C and 1D). Of note, we have previously demonstrated that parallel to the 8 weeks-related intense egg release in feces (Fig 1B) uncovered here, a type 2 immune response characterized by systemic eosinophilia is associated with the formation of egg-encasing hepatic eosinophilic granulomas with robust fibrosis around S. mansoni eggs [40,41]. Although regulatory roles for PGD_2_ in S. mansoni-induced type 2 immune response-biased pathology had been previously observed in functional studies employing the DP1 receptor-deficient mice [19], enhanced PGD_2_ production in the liver during schistosomiasis had not been demonstrated before.
*Characterization of a post-oviposition and host PGD synthase-mediated PGD2 synthesis during S. mansoni infection in mice.Scheme in A outlines the protocol of S. mansoni infection in mice achieved by active percutaneous penetration by 60 cercariae and the continuous treatment with HQL-79 (2.6 μg/day), which was delivered by subcutaneously implanted osmotic pumps at 3.5 weeks post infection (wpi) with S. mansoni. Peritoneal lavages and livers were collected 3 or 8 wpi. B shows a temporal kinetics of S. mansoni egg detection in feces. In C, PGD2 amounts detected by specific EIA kit in cell-free peritoneal fluids. In D, PGD2 or PGE2 levels in liver homogenates detected by specific EIA kits for each prostanoid. Values are expressed as the mean ± SEM from at least 5 animals per group (experiments were repeated at least once). +p < 0.05 compared to non-infected control group. p < 0.05 compared to infected non-treated group.
Administration of HQL-79 — a specific inhibitor of the hPGDS — effectively inhibited the host PGD_2_ synthesis induced by in vivo S. mansoni infection, thus confirming its adequacy as a pharmacological tool for our aims. As schematized in Fig 1A, to achieve systemic and persistent delivery of HQL-79 treatment during long-lasting S. mansoni infection, osmotic pumps containing HQL-79 (flow delivery rate of 2.65 μg/day; for about 30 days) were subcutaneously implanted at 3.5 wpi — therefore, before either oviposition (Fig 1B) or the onset of S. mansoni infection-induced increase of systemic PGD_2_ levels (Fig 1C and 1D). Validating the systemic reach of this administration strategy, as well the HQL-79 inhibitory specificity towards PGD_2_ synthesis, HQL-79 treatment did impair S. mansoni infection-induced PGD_2_ synthesis detected in both peritoneal cavity and liver (Fig 1C and 1D), while did not alter hepatic tissue PGE_2_ levels (Fig 1D).
Inhibition of PGD2 synthesis or DP2 antagonism aggravates fibrosis of S. mansoni-induced hepatic granulomas
The proposed pro-fibrogenic function of endogenous PGD_2_ in hepatic granulomatous response to S. mansoni infection [35,37] was investigated by employing two complementary pharmacological strategies, including either (i) the proven in vivo PGD_2_ synthesis inhibition by HQL-79 treatment (Fig 1C and 1D); or (ii) the blockade of PGD_2_ receptor DP2 by a selective antagonist, CAY10471, also delivered by osmotic pump system (flow rate of 1.75 μg/day; implanted at 3.5 wpi). As shown in Fig 2, S. mansoni infection triggered an intense periovular granulomatous response in livers, presenting enhanced levels of TGF-β and IL-13, as well as locally deposited collagen around S. mansoni eggs entrapped in hepatic granulomas. In contrast to the hypothesis of a pro-fibrogenic role of PGD_2_ in schistosomiasis derived from in vitro studies, in our experiments, both PGD_2_-targeting treatments aggravated, rather than attenuated, the S. mansoni-induced fibrotic reaction in the granulomatous liver. Such unexpected outcome was evidenced by a variety of fibrosis-related hepatic parameters, which were further enhanced by HQL-79 and CAY10471 treatments (Fig 2), including S. mansoni infection-induced: (i) increased hepatic collagen synthesis (as assessed by total hydroxyproline liver content) (Fig 2A and 2B); (ii) deposition of collagen fibers around S. mansoni eggs in hepatic granulomas (Fig 2A and 2B); (iii) hepatic production of the key pro-fibrogenic cytokine TGF-β (Fig 2C and 2D); and (iv) hepatic levels of the prototypical type 2 immune response cytokine IL-13 (Fig 2C and 2D) which is known to display potent fibrogenic activity in liver as well [6,7]. Either HQL-79 (Fig 2A) or CAY10471 (Fig 2B) seemed to worsen overall hepatic S. mansoni fibrosis by specifically increasing localized collagen synthesis within granulomas which were already in development; since both treatments: failed to modify the overall number of egg-encasing granulomas found 8 wpi within S. mansoni-infected livers, while promoting evident expansion of fibrotic area of individual S. mansoni-driven granulomas (Fig 2A and 2B). Notably, these findings also indicate that DP2 receptors activation by endogenous PGD_2_ production — that occurs between 3.5 and 8 wpi — does not play key roles in controlling total egg production or the hepatic burden of parasites. It is important to highlight that our experimental design does not allow general assumptions on whether DP2 activation has endogenous roles in the initial cutaneous phase, oviposition-onset, or granuloma assembly. It is noteworthy that, inversely, DP1 activation by PGD_2_ during S. mansoni infection appears to negatively impact early parasite survival and the successful establishment of infection [19]; a caveat being that DP1 role on S. mansoni parasitism is derived from studies with genetically modified mice [19]. Based on classical opposing effects typically displayed by DP1 versus DP2 activation [12,42], selective antagonism of DP1 receptor could even trigger beneficial anti-fibrogenic outcomes.
*Inhibition of PGD2 synthesis by HQL-79 or selective antagonism of DP2 receptor by CAY10471 promote amplification of overall fibrotic reaction within S. mansoni-infected hepatic tissue.Continuous treatments (delivered by implanted osmotic pumps) with either HQL-79 (2.6 μg/day) or CAY10471 (1.7 μg/day) were initiated at 3.5 wpi with S. mansoni. Livers were collected 8 wpi. A and B show total amounts of hepatic hydroxyproline, total numbers of egg-encasing hepatic granulomas, and area measurements of individual hepatic granulomas. Images are representative collagen-related fluorescence in hepatic granuloma sections (confocal microscopy; PicroSirius red modified staining). C and D show levels of TGF-β and IL-13 detected by specific ELISA kits in liver homogenates of S. mansoni-infected mice. Values are expressed as the mean ± SEM from at least 5 animals per group from representative experiments for each drug studied. +p < 0.05 compared to non-infected control group. p < 0.05 compared to infected non-treated group.
The unexpected pro-fibrotic effect of both HQL-79 and CAY10471 treatments was accompanied by a further increase of the infection-elicited hepatic levels of the pro-fibrogenic factors, TGF-β and IL-13 (Fig 2C and 2D). So, our findings also indicate that the mechanisms involved in the PGD_2_/DP2-driven down-regulatory function may depend on counterbalancing S. mansoni-induced hepatic production of TGF-β and IL-13 in vivo. Strikingly, as already mentioned previously, in vitro studies showed a diametrically opposite pattern for PGD_2_ impact on TGF-β or IL-13 secretion by isolated cells, for instance, schistosomal granuloma-derived hepatic stellate cells [21,34,43]. The in vitro versus in vivo discrepancies unveiled here for the role of PGD_2_ in S. mansoni-driven fibrotic process are likely due to the single cell type feature of in vitro assays, which lacks the sequential cellular activities performed by multiple cell types (vide infra) working in the more complex in vivo setting of hepatic granulomatous reaction. Of note, besides their significant role in the establishment of fibrosis [4,44], these S. mansoni-driven hepatic granulomatous myofibroblasts are also recognized to contribute to local eosinophilia of hepatic periovular granuloma [45].
Endogenous PGD2 mediates S. mansoni infection-elicited eosinophilic inflammation
Eosinophils have recently emerged as protective agents in hepatic tissue, acting towards reinstating liver homeostasis after diverse types of injuries [46,47]. Hepatic granulomatous eosinophilia is an undisputable hallmark of S. mansoni infection, yet these cells are considered at most minor immunomodulators of the protective type 2 immune response or even simple bystanders of the pathogenesis [48–50]. On the other hand, it is well-established that eosinophils represent key players in PGD_2_-regulated inflammatory responses since they function as both cellular sources and targets of PGD_2_ [11,12,15]. With an experimental design not aiming to re-examine eosinophil’s role in S. mansoni-driven immunopathology, here HQL-79 effect on S. mansoni infection-elicited hepatic eosinophilic reaction was examined seeking novel insights on the molecular/cellular mechanisms involved in the unexpected anti-fibrogenic effect of PGD_2_.
Reproducing clinical schistosomiasis, the mouse experimental model of S. mansoni infection used here exhibits, within 8 wpi, a marked systemic eosinophilia characterized by elevated numbers of circulating eosinophils, as well as infiltrating eosinophils found in both peritoneal cavities and in egg-encasing hepatic granulomas (Fig 3). Eosinophil recruitment from the circulation into S. mansoni infection-elicited inflammatory sites is known to be mediated by a variety of parasite- and host-derived chemoattractant molecules, especially host CCL11 [51]. Based on the potent eosinophilotactic activity of PGD_2_ [14,52], we analyzed whether endogenous PGD_2_ would also contribute to S. mansoni infection-induced eosinophilic reaction. As shown in Fig 3, by systemically inhibiting host PGD_2_ synthesis, HQL-79 was capable to impair the establishment of S. mansoni-driven eosinophilia, decreasing blood eosinophil availability (Fig 3A, left panel), eosinophil migration to peritoneal cavities (Fig 3A, right panel) and eosinophil influx into S. mansoni-induced hepatic granulomas (Fig 3B, left and middle panels). Hence, our findings boost the status of hPGDS-derived PGD_2_ to an important host-derived mediator of S. mansoni infection-elicited systemic eosinophilia.
*HQL-79 inhibits eosinophilic reaction and cysLTs production induced by S. mansoni infection.A shows eosinophil numbers found in either peripheral blood or peritoneal compartments 8 wpi with S. mansoni. B highlights the eosinophilic feature of hepatic granulomas (stained by Sirius red), showing representative images and enumeration of granuloma-infiltrating eosinophils as well as the amounts of cysLTs (LTC4/D4/E4) detected by specific ELISA kits in liver homogenates. C displays linear regression curves illustrating the relationships between cysLTS levels and either collagen synthesis (top panel) or granulomatous eosinophilic reaction (bottom panel) found in hepatic tissues. Values are expressed as the mean ± SEM from at least 5 animals per group (experiment was repeated at least once). +p < 0.05 compared to non-infected control group. p < 0.05 compared to infected non-treated group. R-squared (r2) and p values for each linear regression are shown in each panel.
The clear inhibitory effect of HQL-79 towards the eosinophilic feature of S. mansoni-driven inflammatory response indirectly uncovers at least three mechanistic aspects of PGD_2_-mediated regulation of the fibrogenic process of schistosomal hepatic granulomas.
First, HQL-79-unveiled PGD_2_ role in eosinophilia induced by S. mansoni infection is neither a consequence of S. mansoni parasitism inhibition nor a failure to mount the subsequent type 2 immune response. Therefore, the mechanism involved is distinct from the observed in DP1 receptor-deficient model [19]. The inhibition of PGD_2_ synthesis by HQL-79 diminished S. mansoni infection-triggered eosinophilic reaction, although the magnitude of parasitic burden (number of eggs trapped in hepatic tissue) was unaffected (Fig 2A) and associated with a clear type 2 immune environment. Indeed, besides the slightly elevated hepatic IL-13 levels (although not statistically significant) found under HQL-79 treatment (Fig 2C), the PGD_2_ synthesis inhibitor failed to change the increased serum levels of type 2 cytokines IL-5 and IL-13 (S1A Fig) as well as S. mansoni infection-triggered eosinophil production at mouse bone marrows (S1B Fig), a known IL-5-driven phenomenon.
Second, although well-known producers of PGD_2_ [10], at first glance granulomatous eosinophils did not seem to be the main host cell producers of PGD_2_ at hepatic inflammatory site during S. mansoni infection, since: (i) HQL-79-promoted reduction of PGD_2_ derails the establishment of hepatic granulomatous eosinophilia (Fig 3B), and more importantly (ii) S. mansoni infection-induced hepatic PGD_2_ synthesis (as analyzed at 6 wpi) precedes eosinophil appearance at the granulomatous reaction sites (S2 Fig), occurring despite eosinophils absence. Therefore, the main source for PGD_2_ synthesis within schistosomal granuloma must be a hPGDS-expressing cell type which is structurally present at schistosomal granuloma at 6 wpi and before the PGD_2_-mediated eosinophil arrival at hepatic granulomatous tissue. We have previously shown that the S. mansoni-driven hepatic stellate cells (granulomatous myofibroblasts) are capable to produce PGD_2_ in the schistosomiasis context in an HQL-79-sensitive manner [8]. Here, as shown in S2E Fig, stellate myofibroblasts purified from 6 wpi hepatic granulomas (for cell isolation methods see [34]), when stimulated in vitro with a total lipid extract of S. mansoni parasites (for lipid extraction method see [21]), rapidly synthesize and secrete PGD_2_ within 2 h. Inasmuch as S. mansoni-driven myofibroblasts are also known to regulate eosinophilic reaction [45], we can hypothesize that hPGDS-expressing stellate cells are targeted by HQL-79, culminating with both inhibition of hepatic eosinophilia and augmentation of fibrosis-related parameters.
Third, eosinophil does not seem to be the cell source of TGF-β or IL-13 in S. mansoni-infected livers. Even though eosinophils are known cell reservoirs of preformed TGF-β and IL-13 [48,53], HQL-79 decreased eosinophil presence (Fig 3B), whilst increasing the fibrogenic cytokines (Fig 2C) in S. mansoni-driven hepatic granulomas. Such inverse relationship indicates that, once in place within hepatic granulomas, eosinophils may suppress, rather than secrete TGF-β and IL-13 themselves or further their release by other granuloma cell types. At the granulomatous microenvironment, a paracrine downregulatory impact, likely orchestrated by secretory activities of PGD_2_-stimulated eosinophils, may indeed ensure the lower release of TGF-β and IL-13 by other cell sources.
PGD2 elicits LTC4 production during S. mansoni infection: role of eosinophils as LTC4 synthesizing cells
The lipid molecules LTC_4_, LTD_4,_ and LTE_4_, collectively known as cysteinyl leukotrienes (cysLTs) are notoriously pro-fibrogenic mediators in a variety of lung conditions [54–56] and similar pro-fibrogenic activities for cysLTs can be anticipated in other organs. However, for hepatic fibrosis of any etiology, the role of these molecules is yet uncharacterized; a notable exception is a study performed by Toffoli and coworkers (2006) showing that 5-LO-derived metabolites of unknown cellular origin seem to act as suppressing signals of hepatic granulomatous fibrosis in S. mansoni infection [57,58]. Here, HQL-79 treatment decreased total amounts of cysLTs found in S. *mansoni-*infected hepatic tissue (Fig 3B). Concurring with a cause/effect relationship, and therefore a likely anti-fibrogenic function for these lipid molecules in schistosomal granuloma of infected livers, Fig 3C (top panel) shows a significative inverse correlation (r^2^ = 0.7029; p ≤ 0.05) between hepatic levels of cysLTs and the fibrosis marker hydroxyproline.
S. mansoni infection-induced egg-entrapping hepatic granulomas are comprised of various functionally active cells, including hepatic stellate myofibroblasts, mast cells, macrophages, and eosinophils [41,51,59]. In addition to S. mansoni eggs themselves, all these granuloma-associated cells present the ability to synthesize LTC_4_ [60]. However, cysLTs production is not a ubiquitous cellular activity, it is rather a highly regulated, stimulus-specific, and cell-restricted phenomenon dependent on, for instance, cellular expression and proper intracellular localization of the limiting LTC_4_ synthase enzyme. Eosinophils express the entire LTC_4_ synthesizing enzymatic machinery [61,62], which is promptly activated and compartmentalized within cytoplasmic lipid bodies under PGD_2_ stimulation [15]. Therefore, at this point, we investigated whether (i) S. mansoni infection-induced hepatic LTC_4_ synthesis may take place within PGD_2_-stimulated eosinophils infiltrating hepatic granuloma (vide infra); and (ii) the reduction of hepatic cysLTs observed under HQL-79 may be in part due to the decreased numbers of LTC_4_-synthesizing eosinophils within hepatic granulomas. Both assumptions are strongly supported by the significative positive correlation (r^2^ = 0.8424; p ≤ 0.05) between the magnitude of eosinophil presence and the levels of LTC_4_ found in each granuloma-enriched hepatic tissue of S. mansoni infected mice, treated or not with HQL-79 (Fig 3C; bottom panel). Accordingly, analysis of an early time point (6 wpi) in the S. mansoni infection-driven hepatic granuloma development revealed that, before eosinophil infiltration, the hepatic granulomatous reaction does not show LTC_4_ synthesis (S2D Fig). So far, infiltrating eosinophils appear to synthesize/release LTC_4_ under in situ stimulation by granulomatous PGD_2_ and may represent the main responsible for the 8 wpi increased hepatic levels of cysLTs. Whether activation of eosinophil DP2 receptors mediates this phenomenon was investigated next.
Production of LTC4 by S. mansoni infection-driven eosinophils upon PGD2 activation of DP2 receptors
Besides its eosinophilotactic activity, DP2 receptor is a required element for the successful induction of LTC_4_ synthesis by PGD_2_ in eosinophils — a cooperative intracellular process that demands simultaneous activation of both DP1 and DP2 receptors and takes place within the discrete cytoplasmic lipid body organelles [11,12,15,17]. As shown in Fig 4 and exactly like the inhibition of PGD_2_ synthesis by HQL-79, the selective antagonism of DP2 receptor by CAY10471 did decrease the number of infiltrating eosinophils found 8 wpi within the peritoneal cavity of S. mansoni-infected mice (Fig 4A). Infiltrating peritoneal eosinophils of non-treated infected mice displayed high numbers of cytoplasmic lipid bodies (Fig 4B) as well as showed intracellular immunolabelling for newly synthesized LTC_4_ (Fig 4C and 4D). Antagonism of DP2 by CAY10471 diminished the cytoplasmic content of lipid bodies within infiltrating peritoneal eosinophils (Fig 4B), shut down the ability of some peritoneal infiltrating eosinophils to synthesize LTC_4_ (Fig 4C and 4D), and therefore, partially impaired the overall peritoneal production of cysLTs (Fig 4E) found 8 wpi with S. mansoni. In CAY10471-treated mice, residual peritoneal eosinophilia is still detected (Fig 4A) exhibiting a reduced (38.6 ± 6% of inhibition; n = 5, p ≤ 0.05) LTC_4_ synthesizing ability (Fig 4D and 4E). Together these findings identify eosinophils as, at least, one of the cell types actively participating in in vivo PGD_2_/DP2 receptor-driven stimulation of LTC_4_ synthesis during S. mansoni infection.
*S. mansoni infection induces a PGD2/DP2-mediated cysLTs production by infiltrating peritoneal eosinophils.A shows peritoneal eosinophil counts while B displays the numbers of cytoplasmic lipid body organelles found into peritoneal eosinophils 8 wpi with S. mansoni. C displays representative images of EicosaCell preparations showing intracellular immunolabelled LTC4 (in red; LTC4+ eosinophils). Cellular nuclei are labelled with DAPI (in blue); bar = 10 μm; “EØ” identifies eosinophils among peritoneal cellular population; arrowhead indicates a LTC4- eosinophil (remaining LTC4- cells are mononuclear cells indicated by asterisks). In D, the percentage of peritoneal eosinophil population exhibiting cytoplasmic red immunostaining for LTC4 (LTC4+ eosinophils) is shown. E shows the total amounts of cysLTs (LTC4/D4/E4) detected by specific ELISA kit in peritoneal fluid supernatants. Values are expressed as the mean ± SEM from at least 5 animals per group (each experiment was repeated at least once). +p < 0.05 compared to non-infected control group. p < 0.05 compared to infected non-treated group.
As depicted in Figs 4C and S3, the peritoneal cell population found 8 wpi is not formed only by eosinophils, but by a mixed cell population of eosinophils and mononuclear cells. However, distinct from infiltrating eosinophils, the S. mansoni infection-elicited peritoneal mononuclear cells did not display LTC_4_ synthesizing activity (Fig 4C top image; LTC_4_^-^ mononuclear cells indicated by asterisks). In fact, DP2 receptors appear to regulate neither migration nor activation of peritoneal mononuclear cells in this mice model of S. mansoni infection. CAY10471 treatment did not alter S. mansoni infection-associated numbers of peritoneal mononuclear cells (mixture of mast cells, monocytes/macrophages and lymphocytes) found 8 wpi (S3A Fig), the scarce cytoplasmic lipid bodies found in peritoneal mononuclear cells (S3B Fig) compared to elevated content within eosinophils (Fig 4B), and the lack of intracellular immunolabelling for newly synthesized LTC_4_ (Fig 4C). Therefore, eosinophils seem to represent the cell source of PGD_2_/DP2 receptor-driven cysLTs produced in the peritoneal compartment of S. mansoni-infected animals.
Moving from the peritoneal compartment to the granulomatous liver of S. mansoni-infected mice, CAY10471 produced an even more robust inhibition than those observed with HQL-79 treatment on the magnitude of eosinophilia within hepatic granulomas found 8 wpi (Fig 5A). More than simply ratifying HQL-79 data, and in clear contrast to data showing enhanced allergic pulmonary eosinophilia in DP2 deficient mice [63], CAY10471-driven findings upgraded DP2 to one of the key eosinophilotactic receptors in schistosomiasis, like CCR3 [51].
*DP2 receptor activation triggers S. mansoni granuloma eosinophil response: PGD2-stimulated eosinophils as the cellular source of DP2-driven de novo synthesized LTC4.A shows enumeration of granuloma-infiltrating eosinophils as well as the release of cysLTs (LTC4/D4/E4) detected by specific ELISA kits in liver homogenates. Values are expressed as the mean ± SEM from at least 5 animals per group (experiment was repeated at least once). +p < 0.05 compared to non-infected control group. *p < 0.05 compared to infected non-treated group. B displays linear regression curve illustrating the relationship between cysLTS levels and granulomatous eosinophilic reaction found in hepatic tissues. R-squared (r2) and p values for the linear regression are shown. Panel C schematizes protocol for the isolation of eosinophils from schistosomal hepatic granulomas (a cellular suspension displaying purity of about 85 to 90%). Image in D shows a representative image of intracellularly immunolabeled LTC4 (in red; for EicosaCell preparation vide Methods) within granuloma-isolated eosinophils stimulated in vitro for 1 h with S. mansoni antigen (0.5 μg/mL). Cellular nuclei are labeled with DAPI (in blue); bar = 10 μm; arrowhead indicates an LTC4- eosinophil. Graphs in D show the percentage of in vitro stimulated eosinophils (as indicated) exhibiting cytoplasmic immunostaining for LTC4 (LTC4+ eosinophils) as well as the cytoplasmic numbers of eosinophil lipid body organelles (evaluated in osmium-stained cells). Values are expressed as the mean ± SEM from 5 preparations of granuloma-isolated eosinophils. +p < 0.05 compared to non-stimulated eosinophils. p < 0.05 compared to S. mansoni antigen-stimulated eosinophils.
Besides inhibition of eosinophil accumulation, CAY10471 also promoted the reduction of total hepatic cysLTs (LTC_4_/D_4_/E_4_) content within the S. mansoni infection-induced granuloma-enriched liver (Fig 5A). A positive correlation (r^2^ = 0.7253; p ≤ 0.0018) derived from these data indicates a product/producer relationship between cysLTs and granulomatous eosinophils under CAY10471 treatment (Fig 5B). Taking together with HQL-79 data (Fig 3B), we speculate that an S. mansoni infection-induced DP2 activation by PGD_2_ stimulates in situ LTC_4_ synthesis/release by eosinophils infiltrating hepatic granulomas, therefore bringing about the increased liver content of cysLTs.
Despite the correlation between eosinophil infiltration and cysLT content in the liver, a demonstration that granulomatous eosinophils indeed generate LTC_4_ was still to be determined. To investigate whether the eosinophils present in the fibrotic hepatic granulomas were LTC_4_ synthesizing cells accountable for the increased cysLTs levels found in schistosomal livers, we isolated eosinophils from the S. mansoni hepatic granulomas, stimulated them in vitro with S. mansoni antigen or PGD_2_ for 1 h (with or without CAY10471) and then analyzed lipid body biogenesis as well as intracellular LTC_4_ synthesis (Fig 5C). In this in vitro experimental setting, direct stimulation with PGD_2_ (25 nM) triggered rapid (1 h) intracellular LTC_4_ synthesis within about 70% of granuloma-isolated eosinophils (Fig 5D). So, hepatic granulomatous eosinophils under PGD_2_ stimulation are indeed capable of LTC_4_ synthesis. Moreover, in vitro co-incubation of granuloma-isolated eosinophils with CAY10471 blocked S. mansoni antigen ability to induce LTC_4_ synthesis (Fig 5D). So, at least in vitro, an autocrine/paracrine phenomenon takes place to regulate LTC_4_ synthesis by S. mansoni granuloma-derived eosinophils. Concurring, rapid synthesis of PGD_2_ (1 h) by granuloma-derived eosinophils is also triggered by in vitro stimulation with S. mansoni antigen (S4 Fig). It is noteworthy that all non-eosinophils cells present after processing of the granulomas were negative for PGD_2_. Therefore, one can speculate that, once eosinophils are within hepatic granulomatous environment 8 wpi, they may also assist stellate cells in synthesizing PGD_2_.
For either human blood or in murine pleural fluid cells, eosinophil lipid bodies are the primary subcellular site of PGD_2_-induced LTC_4_ synthesis [11,12,64]. As shown in Fig 5D, immunolabelling of newly synthesized LTC_4_ was in a punctate pattern, with cytoplasmic distribution apart from the perinuclear membrane and fully consistent in size and numbers with eosinophil lipid bodies. As confirmed here (Fig 5D, right graph), in vitro co-incubation with CAY10471 does not modify S. mansoni antigen-triggered lipid body biogenesis, a PGD_2_-induced intracellular event downstream to DP1 receptor activation, rather than DP2 [15]. Of note, in response to PGD_2_, eosinophils are the only characterized cells that synthesize LTC_4_ within these cytoplasmic organelles [10].
Activation of CysLT1 attenuates S. mansoni infection-driven granulomatous fibrosis
The two direct attempts to define eosinophil function in schistosomiasis employing mouse models of eosinophil lineage deficiency, including the studies by Swartz et al. (2006) and de Oliveira et al. (2022), did not uphold the original assumption of eosinophil effector cytotoxic function towards parasites. Instead, these pivotal studies have unveiled a more subtle immunomodulatory role for eosinophils in S. mansoni-evoked granulomatous pathogenesis [49,50]. Particularly regarding S. mansoni infection-triggered hepatic fibrosis within eosinophilic granulomas, de Oliveira and coauthors (2022) revealed that in the absence of eosinophils, reductions in TGF-β and IL-13 levels were accompanied by attenuation of hepatic fibrosis [49], thus indicating a pro-fibrogenic role for eosinophils in the disease. These findings are in clear contrast with the inverse relationship between S. mansoni infection-related eosinophilia and fibrogenesis observed here, whereby decreasing PGD_2_-driven eosinophilia (Figs 3 and 4), HQL-79 and CAY-10471 did increase the fibrogenic cytokines as well as hepatic fibrosis (Fig 2). It seems that targeting specifically PGD_2_-regulated eosinophil functions after the establishment of initial events of infection, rather than the entire eosinophil population from the beginning, appears to promote a distinct pattern of response in schistosomiasis.
We have identified at least one of these PGD_2_/DP2-activated eosinophil effector functions that mediate the endogenous PGD_2_/DP2-driven counter regulatory impact on fibrosis unveiled here: production of LTC_4_ — a molecule with putative anti-fibrogenic effects on S. mansoni infection-induced hepatic granulomatous fibrosis [57]. To directly verify whether hepatic cysLTs down-modulate S. mansoni-induced granulomatous fibrosis, we employed the pharmacological blockade of cysLTs receptor CysLT1 by its selective antagonist, MK571; also delivered by osmotic pump system (flow rate of 2.6 μg/day; implanted at 3.5 wpi). As shown in Fig 6, even though the treatment with MK571 did not interfere with S. mansoni infection-induced PGD_2_ production (Fig 6A) or eosinophil accumulation (Fig 6B) in hepatic granulomatous tissue 8 wpi, the antagonist did enhance the hepatic levels of pro-fibrogenic cytokines TGF-β and IL-13 (Fig 6C), without affecting the number of hepatic granulomas (Fig 6D). So, without interfering with the LTC_4_ cell source or its molecular trigger, i.e. DP2-expressing eosinophils and PGD_2_, the antagonism of CysLT1 receptors reproduced the HQL-79 or CAY10471 pro-fibrogenic effect. This final finding indicates that activation of CysLT1 by eosinophil-derived LTC_4_ and its active metabolites within the schistosomal granuloma represents a mechanism by which endogenous PGD_2_/DP2 axes restrains the buildup of fibrosis through hepatic parenchyma of infected animals.
*MK571 potentiates fibrogenic reaction within S. mansoni infection-driven hepatic granuloma, without interfering with PGD2 synthesis or the eosinophilic reaction.Continuous treatment (delivered by implanted osmotic pumps) with MK571 (2.6 μg/day) was initiated at 3.5 wpi with S. mansoni. Blood samples and livers were collected 8 wpi. A shows PGD2 levels in liver homogenates detected by specific EIA kits. B shows eosinophil numbers found in either peripheral blood or hepatic granulomas. C shows levels of TGF-β and IL-13 detected by specific ELISA kits in liver homogenates. D shows total numbers of egg-encasing hepatic granulomas. Values are expressed as the mean ± SEM from at least 7 animals per group. +p < 0.05 compared to non-infected control group. p < 0.05 compared to infected non-treated group.
Therefore, as schematized in Fig 7, our data uncover an endogenous PGD_2_-driven counter regulatory pathway, which intrinsically limits the development of schistosomal hepatic fibrosis. Stepwise, by the onset of hepatic granulomatous reaction formed around S. mansoni eggs, hPGDS-expressing structural stellate cells of schistosomal granulomas synthesize PGD_2_. Via activation of DP2 receptors, stellate cells-derived PGD_2_ both chemoattracts and activates eosinophils to synthesize/release LTC_4_. Of note, by 8 wpi, PGD_2_ synthesis may also be performed by newly arrived eosinophils, which autocrinally stimulate LTC_4_ release. Acting on CysLT1 receptors-expressing cells, intragranulomatous LTC_4_/D_4_/E_4_ decrease the fibrogenic cytokines TGF-β and IL-13, attenuating the development of hepatic granulomatous fibrosis around the S. mansoni egg.
Endogenous PGD2/DP2-driven counter regulatory impact on the development of schistosomal hepatic fibrosis.The induction of liver granulomatous reaction in an experimental mouse model of S. mansoni infection, is characterized by an initial synthesis of PGD2 by hPGDS-expressing hepatic stellate cells (after later establishment of granulomatous eosinophilia, eosinophils contribute to PGD2 production) (1). Activation of DP2 receptors in granulomatous eosinophils (2) triggers the induction of LTC4 synthesis (3). Released LTC4 and/or its metabolites activate CysLT1 receptors-expressing cells (4), promoting the reduction of fibrogenic cytokines TGF-β and IL-13 (5) that culminates with counter regulation of the development of hepatic granulomatous fibrosis around the S. mansoni egg (6).
Conclusions
The detrimental effect of both HQL-79 and CAY10471 on S. mansoni infection-driven hepatic granulomatous inflammation uncovered here has disclosed an unanticipated endogenous protective mechanism of the hepatic tissue — the in vivo anti-fibrogenic activity of hPGDS-derived PGD_2_ via DP2 activation. In addition, our pharmacological approaches have also unveiled a potential novel PGD_2_-driven regulatory mechanism: DP2 receptors-elicited hepatic granulomatous eosinophils as a major cellular source of anti-fibrogenic LTC_4_ in S. mansoni infection. Noteworthy, since LTC_4_ is known to function also as an intracrine signal capable of triggering IL-4 secretion from eosinophils [65,66], cysLTs may emerge as immunoregulators of the IL-4-mediated type 2 immune response of schistosomiasis.
A main driver for the current study was the in vitro data on PGD_2_ role in hepatic tissue-related fibrosis suggesting that PGD_2_ displays deleterious functions in the context of S. mansoni infection [8,21]; and more importantly, the resulting recommendations for PGD_2_-targeting therapeutic maneuvers in schistosomiasis [35–37]. Because presently there are other clinically approved uses for DP2 receptor antagonism in type 2 immune response-biased pathologies [i.e., ramatroban (Baynas) in allergic rhinitis], our current study compels that drug repurposing should not arise directly from the in vitro studies, and most importantly should be avoided in this case. Concurring, the mechanism involved in DP2 receptor anti-fibrogenic function has recently emerged as a far more complex activity. For instance, it has been shown that the DP2 molecule appears to traffic back and anchor to endoplasmic reticulum membranes where, in a fashion non-related to PGD_2_, DP2 evokes degradation of collagen mRNA and decreases intracellular collagen biosynthesis [67]. Also, data derived from genetically deficient animals should be carefully analyzed before being incorporated into treatment strategies as they may not reflect the proper window of opportunity for the infected patient at risk of developing complications due to hepatic fibrosis. Indeed, based on our current in vivo findings, promoting pharmacological DP2 selective agonism (not antagonism) may be the right track to achieve anti-fibrogenic effects.
Materials and methods
Ethics statement
Male C57BL/6 mice weighing 20 to 25 g were obtained from either CCS/UFRJ or FIOCRUZ breeding centers, raised, and maintained under the same housing conditions. Animals were housed in a temperature-controlled room, under a 12∶12 h light cycle with access to filtered water and chow ad libitum. All animal care and experimental protocols were conducted following the guidelines of the Brazilian Council for Care and Use of Experimentation Animals (CONCEA) in accordance with Brazilian Federal Law number 11.794/2008, which regulates the scientific use of animals (license numbers CEUA115/14 and CEUA139/21 by the Committee for Ethics in Animal Experimentation of Federal University of Rio de Janeiro).
S. mansoni infection and treatments
Mice were infected by active percutaneous penetration of S. mansoni infective stage (BH strain; Institute Oswaldo Cruz, FIOCRUZ, RJ), by exposition of washed mouse paws to 60 alive cercariae for 40 min. Uninfected age-matched mice were used as controls. At 3.5 weeks post infection (wpi), osmotic pumps (Alzet pump; flow rate of 0.11 μL/hour — as stated by the manufacturer) — containing 100 μL of either HQL-79 (1 mg/mL), CAY10471 (670 μg/mL), or MK571 (1 mg/mL) solutions producing a cumulative dose of 2.6, 1.7, or 2.6 μg/day, respectively — were implanted subcutaneously in infected and non-infected mice. All three drugs were diluted with 0.1% DMSO in sterile saline. Osmotic pumps containing vehicle solution implanted subcutaneously did not cause changes in mice survival rate, hepatic dysfunction, or inflammatory alterations within implantation sites, peripheral blood or peritoneal cavities of either non-infected or infected mice (S1 Table). All animals were euthanized under anesthesia after 3, 6, or 8 wpi and during this time, they were maintained under the same housing conditions described before.
Analysis of parasitological parameters
To ascertain the establishment of the infection and characterize oviposition onset, the presence of S. mansoni eggs in feces and hepatic tissue were determined. S. mansoni eggs were identified by morphology (containing sharp lateral spines) and enumerated by copro-parasitological thick-smear Kato-Katz method (Helm-Test Biomanguinhos; FIOCRUZ), while the presence of S. mansoni eggs found trapped in hepatic granulomas was analyzed in liver histological preparations (as described below).
Histopathological analysis
Liver samples were washed in saline, fixed in 10% buffered formalin, dehydrated in alcohol, and embedded in paraffin. Sections (5 μm of thickness) were stained with (i) hematoxylin-eosin for enumeration and area analysis of granulomas with entrapped egg; (ii) modified PicroSirius technique for collagen fibers deposition detection; or (iii) Sirius Red for granuloma-infiltrating eosinophils quantification. Acquisition of digital photographs and quantitative analysis of hepatic tissue sections were carried out using a slider scanner (Pannoramic Midi 3DHistec or Motic Easy Scan) and a computer-assisted image analyzer (Slide Viewer 2.4 or Qu Path 0.4.3). All evaluations were performed by two different blinded observers.
Under light microscopy, the areas of hepatic granulomas (20 granulomas per animal) were determined in digital images (acquired at 20x magnification) of histological sections, containing central eggs, randomly chosen, using the measurement tool of the image analyzer. Egg-encasing granulomas counting was carried out at a low power (20x) magnification under light microscopy. The mean number of egg-encasing granulomas per field was calculated for each infected mouse. By confocal microscopy, total fluorescent intensity within digital images of individual granuloma areas (10 granulomas per animal) was quantified using Image J software. All eosinophils identified in individual granuloma areas were counted in digital images acquired under 80x magnification light microscopy. Ten random granulomas per animal were analyzed and results were expressed as eosinophils/granuloma.
Peripheral blood, peritoneal, and bone marrow eosinophilia
Blood eosinophilia was analyzed by light microscopy of blood smears stained with Panoptic kit. To evaluate peritoneal and bone marrow eosinophilia, cells from peritoneal cavities and bone marrows of removed femurs were harvested with RPMI 1640 medium (Sigma) and cytospun towards slides. Total leukocyte enumeration was performed in Neubauer chambers and differential eosinophil count in Panoptic kit-stained slides under light microscopy.
Eosinophils isolation from S. mansoni-driven hepatic granuloma
Mice livers recovered from 8 to 10 wpi with S. mansoni were homogenized in RPMI 1640 medium. Intact hepatic granulomas were allowed to sediment, washed 3 times with RPMI, and then incubated overnight (37°C; 5% CO_2_). In culture bottles, granuloma-released eosinophils are non-adherent cells in suspension, while other cells are found still within granulomas or attached to the plastic. Recovered cells were analyzed under light microscopy after Panoptic kit staining to determine the percentage of eosinophils in suspension. The eosinophil fraction used in in vitro experiments was composed by 80 to 90% purified eosinophils.
Isolated eosinophils (3 x 10^6^ cells/mL) were co-incubated or not with CAY10471 (200 nM; Cayman Chemicals) and then stimulated for 1 h (37°C; 5% CO_2_) with S. mansoni antigen (0.5 μg/mL; Cusabio) or PGD_2_ (25 nM; Cayman Chemicals). After incubation, eosinophil samples were promptly fixed with PFO for osmium staining or placed in EDAC solution for EicosaCell assay (see below).
Evaluation of hepatic collagen synthesis
As an indirect quantitative assay determining amounts of collagen molecules, hepatic fibrosis was also evaluated by measuring hydroxyproline levels in liver homogenates. Hydroxyproline content was determined in dehydrated/hydrolyzed liver fragments by a colorimetric method in chloramine-T buffer (Sigma, USA) with Ehrlich’s reagent (Sigma)/perchloric acid (Merck) and absorbance was measured at 557 nm. Results were expressed as ng of hydroxyproline per mg of hepatic tissue.
Analysis of lipid mediators production
Amounts of eicosanoids PGD_2_, PGE_2_, and cysteinyl leukotrienes (LTC_4_/D_4_/E_4_) found in peritoneal fluid or homogenized liver fragments were measured by specific commercial EIA kits, according to the manufacturer’s instructions (Cayman Chemicals).
For cysLTs production, intracellular LTC_4_ synthesis was alternatively analyzed by the EicosaCell methodology [68] — an immuno-assay that immobilizes the newly formed eicosanoid within active synthesizing cells. Briefly, peritoneal cells or in vitro-stimulated granuloma-derived eosinophils (cell suspensions in RPMI) were immediately mixed with an equal volume of a 0.2% solution of 1-ethyl-3-(3-dimethylamino-propyl) carbodiimide (EDAC; in PBS), used to crosslink eicosanoid carboxyl groups to amines in proteins. After a 10 min incubation at room temperature with EDAC, eosinophils were washed with PBS, cytospun onto glass slides, fixed with PFO (2%), incubated with PBS with 1% BSA for 15 min, and then incubated with a rabbit anti-LTC_4_ antibody (Cayman Chemicals) overnight. The cells were washed with PBS containing 1% BSA (3 times 10 min) and then incubated with Alexa594 donkey anti-rabbit secondary IgG (Jackson) for 1 h. Nuclear visualization by DAPI staining was employed to distinguish polymorphonuclear eosinophils from mononuclear cells. EicosaCell images were obtained using an Olympus BX51 fluorescence microscope, equipped with a 100X objective in conjunction with LAS-AF 2.2.0 Software.
Cytokines measurements
Amounts of TGF-β, IL-5, and IL-13 in liver fragments or in serum samples were measured by commercial ELISA kits, according to the manufacturer’s instructions (R&D Systems and/or Peprotech).
Lipid body staining and enumeration
For lipid body counting, cells in cytospin slides were fixed in 3.7% paraformaldehyde and stained with 1.5% OsO4 in 0.1 M cacodylate buffer, as previously described [64]. By bright field microscopy, fifty consecutively eosinophils/slide were evaluated in a blinded fashion by more than one observer.
Statistical analysis
Results are expressed as means ± SEM (standard error of the mean) and were analyzed by one-way ANOVA, followed by Student-Newman-Keuls test or by Student’s t test, using Prism software (GraphPad Software, Inc., San Diego, CA, USA). Differences were considered significant when p < 0.05. Two independent experiments were performed for each PGD_2_-targeting treatment studied: HQL-79 and CAY10471.
Supporting information
S1 FigInhibition of PGD_2_ synthesis by HQL-79 treatment did not affect S. mansoni infection-induced immune polarization to a type 2-biased response in mice.A shows serum levels of IL-5 and IL-13. B shows eosinophil numbers found at bone marrow. Values are expressed as the mean ± SEM from at least 5 animals per group (experiment was repeated at least once). ^+^p < 0.05 compared to non-infected control group.(TIF)
S2 FigPGD_2_ synthesis within liver of S. mansoni infected mice precedes installation of hepatic granulomatous eosinophilia: hepatic stellate cells as PGD_2_ cellular source.Peritoneal lavages and livers were collected 6 wpi with S. mansoni cercariae. A shows PGD_2_ amounts detected by specific EIA kit in cell-free peritoneal fluids and in liver homogenates. In B, total numbers of egg-encasing hepatic granulomas are shown. C shows eosinophil numbers found in either peritoneal compartment, hepatic granuloma, peripheral blood, or bone marrow space. D shows peritoneal and liver LTC_4_ levels. Values are expressed as the mean ± SEM from at least 5 animals per group. ^+^p < 0.05 compared to non-infected control group. In E, hepatic granuloma-derived stellate cells were isolated 6 wpi from S. mansoni-infected mice and then stimulated in vitro for 2 h with total lipids extracted from isolated parasites; PGD_2_ synthesis was quantified in cell-free supernatants. Values are expressed as the mean ± SEM from 2 preparations of hepatic granuloma-derived stellate cells.(TIF)
S3 FigAntagonism of DP2 receptor by CAY10471 treatment did not affect S. mansoni infection-elicited population of peritoneal mononuclear cells.A shows numbers of mononuclear cells found at peritoneal space as well as representative images of peritoneal cells 8 wpi of S. mansoni-infected mice (top image) and in mice treated with CAY10471 and S. mansoni infected (bottom image). B shows numbers of cytoplasmic lipid body organelles found in peritoneal mononuclear cells. Values are expressed as the mean ± SEM from at least 5 animals per group (experiment was repeated at least once). ^+^p < 0.05 compared to non-infected control group.(TIF)
S4 FigIn vitro stimulated hepatic granuloma-isolated eosinophils synthesize PGD_2_.Eosinophils were isolated from 8 wpi schistosomal hepatic granulomas (purity about 90%) and stimulated in vitro for 1 h with S. mansoni antigen (0.5 μg/mL). Percentage of in vitro stimulated eosinophils (as indicated) exhibiting cytoplasmic immunostaining for LTC_4_ (LTC_4_^+^ eosinophils) under fluorescence microscopy in Eicosacell preparations are shown. Values are expressed as the mean ± SEM from 3 preparations of granuloma-isolated eosinophils. ^+^p < 0.05 compared to non-stimulated eosinophils.(TIF)
S1 TableLack of impact of 0.1% DMSO solution (the vehicle solution employed for HQL-79, CAY10471 or MK571 treatments delivered by osmotic pumps) on non-infected or S. mansoni-infected animals.S. mansoni infection in mice was achieved by active percutaneous penetration by 60 cercariae. Osmotic pumps containing a 0.1% DMSO solution were subcutaneously implanted (3.5 wpi) in both non-infected and S. mansoni-infected animals (columns labelled “DMSO“). Animals that were not implanted with subcutaneous pumps were used as controls (columns labelled “-“). All animals were alive at 8 wpi, when they were euthanized. Livers of all non-infected animals were normal, showing no macroscopic or histopathological alterations. Individual values and mean ± SEM from 2 or 3 animals per group are displayed.(DOCX)
S2 TableRaw data for the graphs.The data for each group presented in the manuscript is listed in individual tables for each graph indicated by Figure number and letter. Mean ± SEM for each group is provided in the bottom line.(DOCX)
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Zwang J, Olliaro P. Efficacy and safety of praziquantel 40 mg/kg in preschool-aged and school-aged children: a meta-analysis. Parasit Vectors. 2017;10: 47. doi: 10.1186/s 13071-016-1958-7 28126024 PMC 5270314 · doi ↗ · pubmed ↗
- 2Zaparina O, Rakhmetova AS, Kolosova NG, Cheng G, Mordvinov VA, Pakharukova MY. Antioxidants resveratrol and Sk Q 1 attenuate praziquantel adverse effects on the liver in Opisthorchis felineus infected hamsters. Acta Trop. 2021;220: 105954. doi: 10.1016/j.actatropica.2021.105954 33979641 · doi ↗ · pubmed ↗
- 3Niu X, Hu T, Hong Y, Li X, Shen Y. The Role of Praziquantel in the Prevention and Treatment of Fibrosis Associated with Schistosomiasis: A Review. J Trop Med. 2022;2022: 1413711. doi: 10.1155/2022/1413711 36313856 PMC 9616668 · doi ↗ · pubmed ↗
- 4Mederacke I, Hsu CC, Troeger JS, Huebener P, Mu X, Dapito DH, et al. Fate tracing reveals hepatic stellate cells as dominant contributors to liver fibrosis independent of its aetiology. Nat Commun. 2013;4: 2823. doi: 10.1038/ncomms 3823 24264436 PMC 4059406 · doi ↗ · pubmed ↗
- 5Anthony B, Mathieson W, de Castro-Borges W, Allen J. Schistosoma mansoni: egg-induced downregulation of hepatic stellate cell activation and fibrogenesis. Exp Parasitol. 2010;124: 409–420. doi: 10.1016/j.exppara.2009.12.009 20045695 · doi ↗ · pubmed ↗
- 6Chiaramonte MG, Cheever AW, Malley JD, Donaldson DD, Wynn TA. Studies of murine schistosomiasis reveal interleukin-13 blockade as a treatment for established and progressive liver fibrosis. Hepatology. 2001;34: 273–282. doi: 10.1053/jhep.2001.26376 11481612 · doi ↗ · pubmed ↗
- 7Chiaramonte MG, Donaldson DD, Cheever AW, Wynn TA. An IL-13 inhibitor blocks the development of hepatic fibrosis during a T-helper type 2-dominated inflammatory response. J Clin Invest. 1999;104: 777–785. doi: 10.1172/JCI 7325 10491413 PMC 408441 · doi ↗ · pubmed ↗
- 8Paiva LA, Coelho KA, Luna-Gomes T, El-Cheikh MC, Borojevic R, Perez SA, et al. Schistosome infection-derived Hepatic Stellate Cells are cellular source of prostaglandin D₂: role in TGF-β-stimulated VEGF production. Prostaglandins Leukot Essent Fatty Acids. 2015;95: 57–62. doi: 10.1016/j.plefa.2015.01.004 25687497 · doi ↗ · pubmed ↗