Genome characterization of BI2 subcluster Streptomyces scabiei bacteriophages GoblinVoyage and Doxi13
Hanna Jin, Nihal K. Chana, Annie L. Tang, Paramjit Kaur, Brishti Lamichhane, Sze Ching Leung, Diane Scheiderer, Vighnesh V. Sivaprakasam, Dannah T. Marcelino, Gregory J. Hull, Toma M. Kamara, Maria C. Guimaro, Steven M. Caruso

TL;DR
This paper describes two new bacteriophages, GoblinVoyage and Doxi13, that infect Streptomyces scabiei and have similar genome structures.
Contribution
The study introduces two new siphoviruses in the BI2 cluster with identical 3’ end sticky overhangs and high GC content.
Findings
GoblinVoyage and Doxi13 have genome lengths of 43,540 bp and 43,696 bp, respectively.
Both phages have a GC content of 60.9% and identical 3’ end sticky overhangs.
Abstract
We present the bacteriophages GoblinVoyage and Doxi13, siphoviruses isolated on Streptomyces scabiei RL-34. They belong to the BI2 cluster and have genomes consisting of 60.9% GC content with identical 3’ end sticky overhangs. The genome lengths of GoblinVoyage and Doxi13 are 43,540 bp and 43,696 bp, respectively.
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
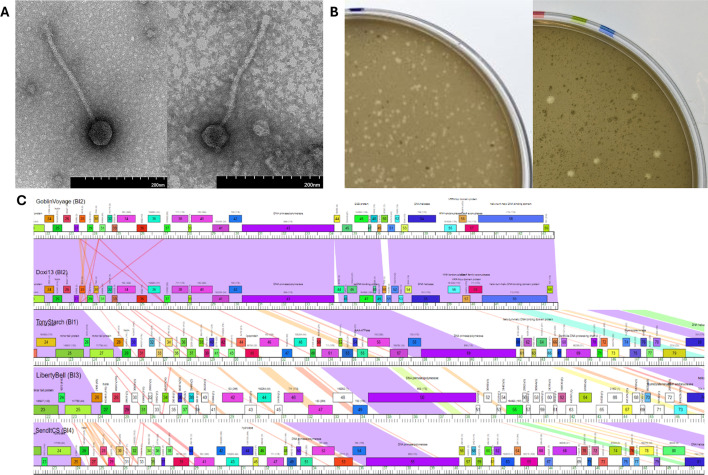 Fig 1
Fig 1| Phage characteristics | GoblinVoyage | Doxi13 |
|---|---|---|
| Isolation host | ||
| Sample type | Dry, dusty soil | Dry, sandy soil |
| Sample location [GPS] | Golansville, Virginia, USA | Laurel, Maryland, USA |
| Approximate shotgun coverage | 1,166 bp | 10,298 bp |
| Total number of reads | 357,901 | 3,008,314 |
| Approximate fold coverage | 1,233 x | 10,327 x |
| Genome length | 43,540 bp | 43,696 bp |
| GC% content of phage | 60.9% | 60.9% |
| Genome end type | 3' sticky overhangs | 3' sticky overhangs |
| Character of genome ends | 5′-CGCCGCCCT-3′ | 5′-CGCCGCCCT-3′ |
| Identified protein coding genes (assigned a function) | 59 (29) | 60 (21) |
| Identified tRNA encoding genes | 0 | 0 |
- —HHS | NIH | National Institute of General Medical Sciences (NIGMS)
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsBacteriophages and microbial interactions · Plant Disease Resistance and Genetics · Genomics and Phylogenetic Studies
ANNOUNCEMENT
Streptomyces scabiei is a Gram-positive phytopathogen, primarily responsible for the common potato scab disease (1). Genomic characterization of bacteriophages that infect S. scabiei reveals roles that they may serve to address the deleterious effects of the infection.
Samples of bacteriophages GoblinVoyage and Doxi13 were collected from soils using S. scabiei RL-34 as a host (Table 1). All protocols were derived from the Science Education Alliance-Phage Hunters Advancing Genomics and Evolutionary Science (SEA-PHAGES) Phage Discovery Guide (2). Soil samples were combined with phage buffer (10 mM [Tris pH 7.5], 10 mM MgSO_4_, 68 mM NaCl, and 1 mM CaCl_2_) and passed through a 0.22-µm filter. Then, 500 µL of the filtrate was added to 250 µL of a 48-hour S. scabiei culture, incubated for 10 minutes, combined with 3 mL of tryptic soy soft agar (BD), then plated onto nutrient agar plates (BD Difco) with Streptomyces phage supplement (10 mM MgCl_2_, 8 mM Ca(NO_3_)2, 0.5% glucose) and incubated for 24–48 hours at 30°C. Phages underwent a minimum of three rounds of plaque purification. In short, picked plaques were serially diluted tenfold and used for plaque assays on the host. The fresh lysate was harvested from plates containing near-confluent lysis for imaging and DNA isolation. Negative-stained transmission electron microscopy revealed siphoviral morphotypes for both GoblinVoyage and Doxi13 (Fig. 1A). Both phages demonstrated a similar plaque morphology (Fig. 1B). Neither phage produced discernable lysogens when tested by spotting dilutions of the lysate on S. mirabilis and incubating for 7 days at 30°C, suggesting they followed a lytic infection cycle.
Morphological and functional characterization of BI2 phages GoblinVoyage and Doxi13. (A) Representative transmission electron microscopy images of GoblinVoyage and Doxi13. The fresh lysate sample was stained with 2% uranyl acetate. TEM imaging revealed GoblinVoyage and Doxi13 are siphoviruses with flexible and noncontractile tails. GoblinVoyage (left) has a capsid diameter of 58 nm (n = 3, SD = 2.5 nm) and tail length 261 nm (n = 3, SD = 1.53). Doxi13 (right) has a capsid diameter of 52 nm (n = 3, SD = 1.5) and tail length 248 nm (n = 3, SD = 12.56). All measurements were taken using ImageJ v1.54i (3). (B) Plaque morphology of GoblinVoyage and Doxi13 (left to right). Both phages showed turbid plaques with an average diameter of 1.10 mm (n = 30, SD = 0.337) and 1.60 mm (n = 16, SD = 0.46), respectively. Doxi13 plaques were observed in S. mirabilis. Plaques were measured after approximately 24 to 48 hours of incubation at 30°C. All measurements were taken using ImageJ v1.54i (3). (C) Comparison of GoblinVoyage and Doxi13 genome size and GoblinVoyage to other BI subcluster phages. From top to bottom: GoblinVoyage (BI2), Doxi13 (BI2), TonyStarch (BI1), LibertyBell (BI3), and SendItCS (BI4). All phages are compared to GoblinVoyage from approximately 21,500 bp. All phages selected are representative or the only member of their subcluster. Phages from subclusters BI5, BI6, and BI7 show similar results and are not included. Image produced using Phamerator vActino_Draft 558 (4).
Phage DNA was extracted from the freshly prepared crude lysate using the Promega Wizard DNA cleanup kit and sequenced at the Pittsburgh Bacteriophage Institute using the NEB Ultra II Library Kit and an Illumina MiSeq with v3 reagents with 150-bp single-end raw reads. The sequencing results are described in Table 1. Newbler v2.9 (5) and Consed v29 (6) were used for assembly, quality verification, and end determination as described (7).
Genome annotation was completed using DNA Master v5.23.6 (8) with internal Glimmer v3.02 (9) and GeneMarkS v2.5 (10, 11) programs. GeneMark.hmm v2.5p (10, 11) was used to confirm these annotations with S. scabiei 87–22 as the species. Putative functions were assigned by assessing amino acid sequence homology using NCBI BLASTp v2.15.0 (12) and Conserved Domain Database v3.19 (13), structural homology with HHpred v57c87 (minimum probability: 90%) (14, 15), and synteny via Phamerator vActino_Draft 558 (4). Default parameters were used for all software, unless otherwise specified. The phages were assigned to cluster BI2 in the Actinobacteriophage Database (PhagesDB) based on gene content as described (16, 17) and the genus Scapunavirus (18). Genomic characterization and annotation results are reported in Table 1.
Annotations of GoblinVoyage and Doxi13 failed to identify genes consistent with a temperate phage, agreeing with the earlier virulent designation and classification of the cluster in PhagesDB. Genomic comparison of phages from BI subclusters shows that BI2 phages have smaller genomes, differing by approximately 10 kbp, including multiple small genes in the center of the genome (Fig. 1C).
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Li Y, Liu J, Díaz-Cruz G, Cheng Z, Bignell DRD. 2019. Virulence mechanisms of plant-pathogenic Streptomyces species: an updated review. Microbiology (Reading) 165:1025–1040. doi:10.1099/mic.0.00081831162023 · doi ↗ · pubmed ↗
- 2Poxleitner M, Pope W, Jacobs-Sera D, Sivanathan V, Hatfull G. 2018. SEA-PHAGES phage discovery guide. HHMI SEA-PHAGES phage discovery guide. Howard Hughes Medical Institute, Chevy Chase.
- 3Collins TJ. 2007. Image J for microscopy. Bio Techniques 43:25–30. doi:10.2144/00011251717936939 · doi ↗ · pubmed ↗
- 4Cresawn SG, Bogel M, Day N, Jacobs-Sera D, Hendrix RW, Hatfull GF. 2011. Phamerator: a bioinformatic tool for comparative bacteriophage genomics. BMC Bioinformatics 12:395. doi:10.1186/1471-2105-12-39521991981 PMC 3233612 · doi ↗ · pubmed ↗
- 5Margulies M, Egholm M, Altman WE, Attiya S, Bader JS, Bemben LA, Berka J, Braverman MS, Chen Y-J, Chen Z, et al.. 2005. Genome sequencing in microfabricated high-density picolitre reactors. Nature 437:376–380. doi:10.1038/nature 0395916056220 PMC 1464427 · doi ↗ · pubmed ↗
- 6Gordon D, Abajian C, Green P. 1998. Consed: a graphical tool for sequence finishing. Genome Res 8:195–202. doi:10.1101/gr.8.3.1959521923 · doi ↗ · pubmed ↗
- 7Russell DA. 2018. Sequencing, assembling, and finishing complete bacteriophage genomes. Methods Mol Biol 1681:109–125. doi:10.1007/978-1-4939-7343-9_929134591 · doi ↗ · pubmed ↗
- 8Pope WH, Jacobs-Sera D. 2018. Annotation of bacteriophage genome sequences using DNA master: an overview. Methods Mol Biol 1681:217–229. doi:10.1007/978-1-4939-7343-9_1629134598 · doi ↗ · pubmed ↗