Thermal Relationship in Tropical Anurans from Two Contrasting Habitats Along an Elevation Gradient in Colombia
Katalina Gutiérrez Hernández, Carlos Alberto Galindo Martínez, Jorge Luis Turriago González, Manuel Hernando Bernal Bautista

TL;DR
This study examines how body temperatures of frogs in different habitats and elevations in Colombia are influenced by environmental temperatures.
Contribution
The study reveals that habitat type strongly influences frog body temperatures at similar elevations, adding to thermal biology understanding.
Findings
Frog body temperatures are more influenced by substrate than air temperature.
Open-habitat frogs have higher body temperatures than forest-dwelling frogs at the same elevation.
Body temperatures of frogs decrease with increasing elevation.
Abstract
Anurans are ectothermic organisms highly susceptible to variations in the environmental temperature that changes with elevation and between habitats in tropical mountains. The aim of this study was to evaluate the variation of body temperature (BT) of nocturnal anurans from two contrasting habitats (open and forest habitats) along an elevation gradient in Colombia. We measured the environmental temperatures (substrate and air) and BT of 135 adult frogs of 11 species from open and forest habitats at three elevational zones of an Andean Mountain. The BT had a positive and significant relationship with environmental temperatures and showed a higher thermal dependence for substrate than air temperature, which suggests that anurans are thermoconformers and potentially tigmotherms. Additionally, BT of anurans from both habitats decreased with the elevation, but species from open habitats had…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
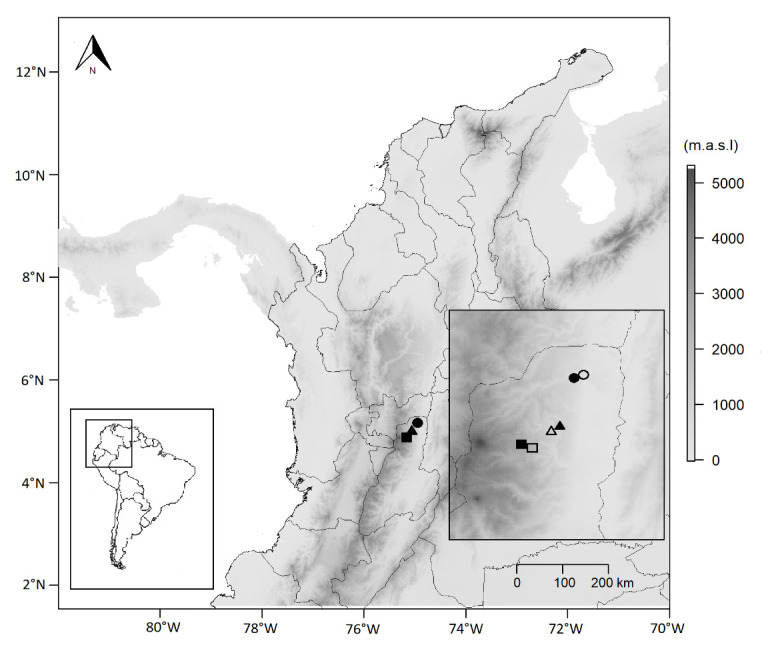 Figure 1
Figure 1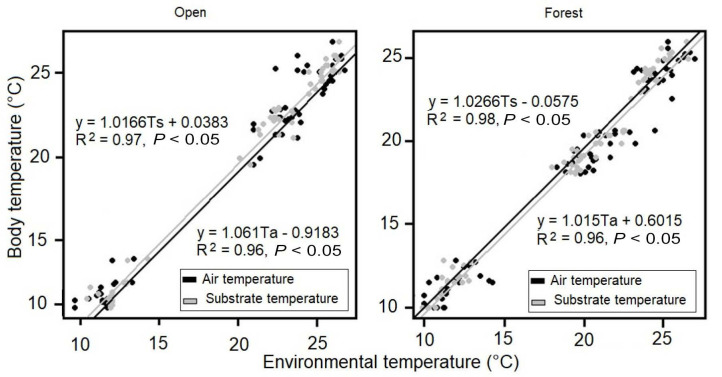 Figure 2
Figure 2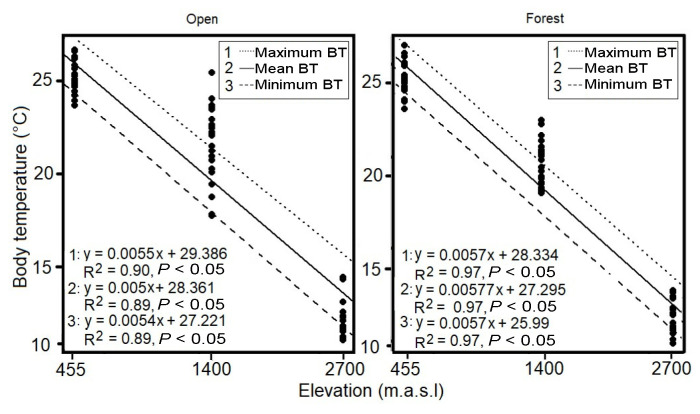 Figure 3
Figure 3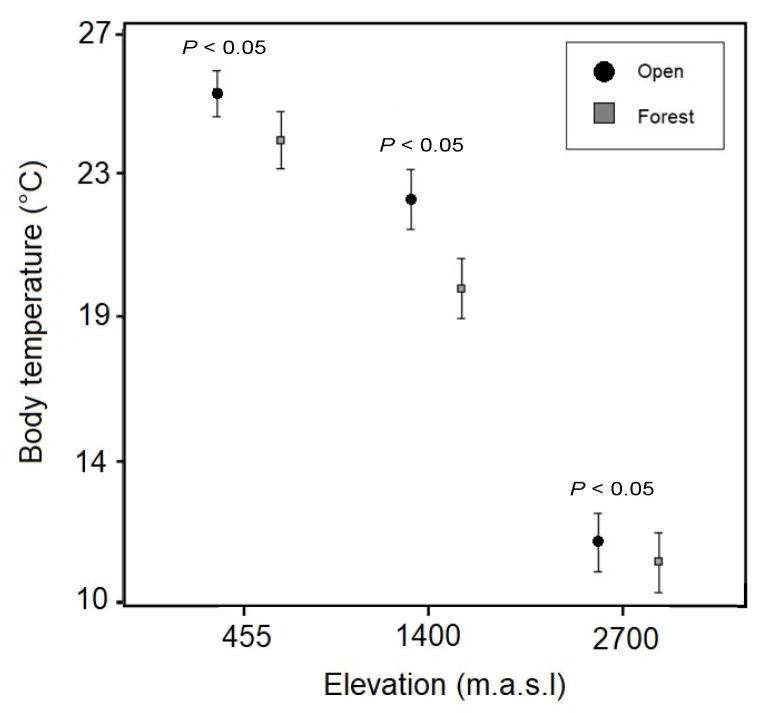 Figure 4
Figure 4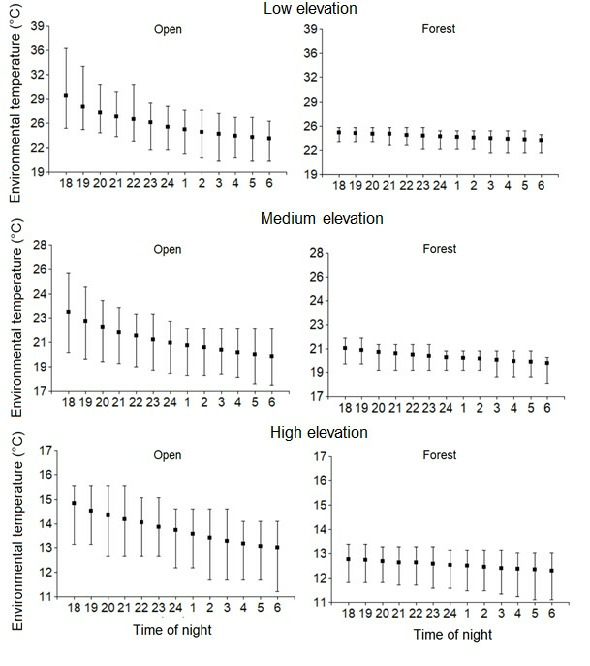 Figure 5
Figure 5- —Vicerrectoría de Investigación-Creación, Innovación, Extensión y Proyección Social (Vice President of Research-Creation, Innovation, Extension and Social Projection) of the Tolima University
- —“Formación de Talento Humano de Alto Nivel” approved by the Fondo de Ciencia, Tecnología e Innovación (CTeI) of Sistema General de Regalías (SGR)
- —Tolima Governorate and Tolima University, Colombia
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsAmphibian and Reptile Biology · Species Distribution and Climate Change · Primate Behavior and Ecology
INTRODUCTION
Environmental temperatures play an important role in the physiological and behavioral functions of amphibians, as they mostly regulate their body temperatures (Angilletta 2009; Duellman & Trueb 1994; Hillman et al. 2008). Additionally, local variation in environmental temperatures may determine the selection of microhabitats where amphibians can carry out successfully their biological activities (Nowakowski et al. 2017). In tropical mountains, the thermal environment is affected by elevation due to the gradual decrease in air temperature as elevation increases, the adiabatic cooling (Ehlers & Poulsen 2009). The thermal environment can be also modified by climatic change and habitat conversion which generally increase the temperatures (Hussain & Pandit 2012; Nowakowski et al. 2017; Nowakowski et al. 2018; Root et al. 2003). Particularly, at a microclimate scale, variations in the cloud and vegetation cover, wind speed and relative humidity, strongly influence the convection and conduction of microenvironmental temperatures (Gates 1980), which are the two main mechanisms of heat exchange in nocturnal amphibians (Pough et al. 2015). Consequently, changes in environmental temperatures may alter the geographical distribution, temporal activity, physiological and behavioural performance of amphibians, predominantly in endemic and thermal specialised species as those that inhabiting the Andean mountains of Colombia (Bernal & Lynch 2013; Lynch & Suárez 2002).
The thermal ecology of amphibians has been studied extensively for many years (Brattstrom 1963; Iturra et al. 2014; Navas et al. 2013; Taylor et al. 2020). However, there is limited information on the body temperatures (BT) of tropical anurans in relationship to microenvironmental temperatures, which differ significantly from macroclimate data that have been demonstrated to have limited predictive power in small animals, such as amphibians (Sanabria & Quiroga 2019). Therefore, information on average temperature and thermal variation of microhabitats seems indispensable to understand amphibian BT in tropical mountain systems, beyond the known phenomenon of BT decreasing with elevation (Navas et al. 2013). These microclimate data are especially important in the Andes because they are a global hot spots of anuran species richness and endemism (Armesto & Señaris 2017; Ortiz et al. 2013), and because Andean amphibians are very susceptible to decline in response to climate change (Catenazzi et al. 2014; Corn 2005; Delgado & Burrowes 2022; Hussain & Pandit 2012; Iturra et al. 2014; Navas et al. 2013).
In this work, we recorded the environmental temperature and BT of tropical anurans from two contrasting habitats along an elevation gradient in an Andean Mountain of Colombia. Specifically, we evaluated the influence of: (1) air and substrate temperatures (Ta and Ts), (2) elevation, and (3) type of habitat (open vs forest) on BT of nocturnal anurans. As amphibians are considered tigmotherms (Cruz et al. 2016; Duellman & Trueb 1994), we expected that BT would show a stronger association with Ts than Ta, thus indicating to be a better predictor of variations in activity temperature of anurans. We also expected a negative relationship between BT and elevation, in response to environmental temperature decreasing as elevation increases (Navas et al. 2013; Ramírez et al. 2009). Finally, we hypothesised that anurans from open habitats (which are daily exposed to greater solar radiation) would have a higher mean and thermal variation than those from forest habitats along the studied elevational gradient. We hypothesised this result given the thermal stability of forest environments (Bohlman et al. 1995; Navas et al. 2013). Therefore, this paper aims to contribute to a better understanding of the thermal variation in two contrasting habitats (forest and open) across elevation and its impact on anuran BT.
MATERIALS AND METHODS
Study Sites and Species
This work was carried out at three different elevations (455 m.a.s.l, 1,400 m.a.s.l and 2,700 m.a.s.l) along the Cordillera Central of Colombian Andes, at the Tolima department, Colombia. The specific locations studied and their coordinates are detailed in Fig. 1. At each elevation, during three consecutive nights per month, from September to December 2018, two people actively searched adult anurans, based on acoustic and visual encounters (Angulo et al. 2006), both in forests as well as in open habitats, at approximately the same nocturnal hours, from 18:00 h to 24:00 h. Thus, our cumulative sampling effort was approximately of 72 h per person. We defined a forest habitat when the associated vegetation was mainly arboreal, with a close canopy higher than 60% (in which the land surface is covered by tree canopies), while a habitat was considered open when it had a maximum canopy cover (open canopy) up to 40% (FAO 2001; Galeana et al. 2009).
Measures of Microenvironmental and Anuran Body Temperatures
Temperatures from anuran microhabitats were obtained with six data loggers IButton (DS1921G, accuracy of ± 1°C), which were placed in both the forest and open habitats for each of the three elevational zones studied (2 × 3 = 6 IButtons). Specifically, each data logger we positioned on the ground, in direct contact with the soil surface, where adult anurans were found. The data loggers were programmed to register temperatures each hour from September to December 2018, broadly coinciding with rainy season and frog samplings.
Anuran BT and environmental temperatures (Ts and Ta) were measured simultaneously in the field with two thermometers Extech (421502, precision 0.3°C, 0.05% accuracy) and their two thermocouples type K. The BT was taken over the dorsal part of animal, whereas Ts was recorded directly in the place where the animal was found, such as on the surface of the ground, trunks, leaf litter, soil vegetation (pastures, mosses) or leaves of trees. When some animals were found in water, Ts was recorded as the temperature of the water surface. The Ta was measured 20 cm above the animal. Previously, in the laboratory of Herpetology at the Tolima University, at different hours of the day, we simultaneously measured 10 times the skin temperature and cloacal temperature (with the thermocouple type K and thermometer Extech) of nine animals from four species with different body sizes, so as to evaluate the differences between external and internal temperatures. We did not find significant differences (Repeated Measures ANOVA, p > 0.05 in all cases), as it has been reported in other amphibians (Cruz et al. 2016; Oromí et al. 2010). Therefore, we assumed that skin temperature reported in this study represents the internal body temperature of anurans. We also measured the snout-vent length of anurans (SVL) using a digital caliper (Mitutoyo, precision 0.01 mm).
Statistical Analyses
We used descriptive statistics (mean ± standard deviation, range) to show the thermal environmental variables and BT. We conducted linear regressions and Pearson correlations to evaluate separately the relationship between BT and environmental temperatures (Ta and Ts), as data met the assumption of normality (Shapiro-Wilks test, p > 0.05). Additionally, we performed an ANCOVA to compare the BT of anurans from open vs forest habitats at each of the three elevational zones studied, using the SVL as covariable, and the Levene’s test to compare the variances of BT between these two habitats. All statistical analyses were realised in RStudio version 4.1.2.
We recognise the importance of phylogenetic analyses for interspecific comparative studies; however, some previous data suggest that elevation and natural history may influence BT more than shared ancestry (Navas et al. 2013). In addition, due to the limited number of species studied, our data are likely not suitable for phylogenetic analysis (Blomberg et al. 2003). For that reason, herein we analysed our data employing conventional statistical analyses independent of phylogenetic position.
RESULTS
Data on the species studied, anuran BT and environmental temperatures for the forest and open habitats along the elevational gradient sampled are shown in Table 1. Mean BT varied from 11.0°C in Pristimantis permixtus at 2,700 m.a.s.l to 25.2°C in Rhinella margaritifera at 455 m.a.s.l. The highest BT variation within species was 3.1°C (range: 23.7°C–26.8°C) for R. margaritifera from the open habitat at 455 m.a.s.l, whereas the lowest one was 1.5°C (range: 10.0°C–11.5°C) for P. permixtus from the forest habitat at 2,700 m.a.s.l (Table 1).
In both open and forest habitats, BT had a positive and significant relationship with environmental temperatures (Fig. 2). Particularly, Pearson’s test was higher than 0.65 for the relationship between BT and Ts in all cases, whereas it was lower than 0.65 between BT and Ta (Table 2). Body temperature of anurans (mean, maximum and minimum) decreased significantly with elevation in both open and forest habitats (Fig. 3). Similarly, Pearson correlation coefficient (r) showed a high inverse linear relationship between BT and elevation in both habitats studied (open habitat, r = −0.94; forest habitat, r = −0.99).
Anurans from open habitat had a significantly higher BT than those from forest habitat, controlling for differences in body size (ANCOVA, F = 66.36, p < 0.001). However, the variance of the BT was not significant between open and forest habitats (Levene’s test, p > 0.05). Particularly, comparing the anuran BT between these two habitats but within each elevational zone studied, the difference was statistically significant for the species from both low and medium zones (Tukey test p < 0.05), but not for those from the high elevation (Tukey test p > 0.05) (Fig. 4). Nevertheless, microenvironmental nightly temperatures (from 18:00 h to 6:00 h) showed higher hourly mean temperatures and greater range of variation for open than forest habitats in all three elevational zones studied (Fig. 5).
DISCUSSION
The positive and significant relationship between anuran body and environmental temperatures in two contrasting habitats in an Andean elevational gradient in Colombia (Fig. 2), indicates that frogs have the tendency to thermoconformity (i.e., BT parallels changes in environmental temperatures), as has also been reported in other related amphibian studies (Carvajalino et al. 2011; Iturra et al. 2014; Rueda et al. 2016). This thermal strategy results important because it may reduce the water loss maintaining the skin wet (Duellman & Trueb 1994) while amphibians perform their routine activities, such as foraging, calling, breeding and resting (Sanabria et al. 2011; Sanabria et al. 2014). Additionally, the stronger correlation of BT with Ts than Ta (Table 2), suggests that anurans present a tigmothermal regulation, where the conduction is probably the main way of heat exchange (Lara & Luja 2018; Leyte et al. 2018), as seen in other nocturnal amphibians, such as Pleurodema thaul (Iturra et al. 2014), and nine of the eleven species studied by Lara and Luja (2018). Thus, these data demonstrate that Ts is a better predictor of anuran BT than Ta, similar to that found in other amphibian species, Leptodactylus ocellatus (Sanabria et al. 2003), Pleurodema thaul (Iturra et al. 2014), and most of the anurans studied by Navas et al. (2013). This is an important finding for generating more realistic models about the effects of elevation, or environmental variables, on the physiological and behavioural performance of amphibians, especially when unusual or abrupt thermal changes may have a strong value in terms of thermal ecology and conservation of amphibians.
As expected, BT of anurans decreased through elevation in both habitats (open and forest) (Fig. 3), like other neotropical salamanders (Feder & Lynch 1982), and amphibians from the eastern Nepal Himalaya (Khatiwada et al. 2020), and the tropical Andes (Navas et al. 2013; Pintanel et al. 2019). This trend may be explained by the thermoconformity of amphibians which follows the inverse relationship between microenvironmental temperatures and elevation (Khatiwada et al. 2020; Navas et al. 2013). This result also suggests that elevation may have a stronger impact on BT than phylogeny (Navas et al. 2013), as this comparison was made among different taxa and within the same species located at different elevation, e.g., B. platanera and R. margaritifera, with similar outcomes, a lower anuran BT at the highest elevation (Table 1). Furthermore, the effect of differences in temperatures between two elevations was clearly more noticeable on anuran BT than the effect of differences in temperature between the two microhabitats studied at the same elevation, even though microhabitats can generate small areas with stable environmental conditions which may diverge from this main trend (Jones et al. 2011; Pintanel et al. 2019). For instance, the minimum BT difference between two elevational zones was 3.2°C in open habitats (Mariquita: 455 m.a.s.l and Líbano: 1,400 m.a.s.l: 25.1°C–21.9°C), whereas the maximum BT difference between forest and open habitats was 2.6°C (21.9°C–19.3°C) in Líbano (Tolima), the intermediate elevational zone (Table 1). Then, in this work we detected that changes in elevation caused a higher effect on anuran BT than microhabitat temperatures.
The mean of anuran BT was significantly higher in open than forest habitats at the same elevational level (450 and 1,400 m.a.s.l) (Fig. 4). This result agrees with the information reported by Soto et al. (2017) in the frog Agalychnis dacnicolor, whose BT was significantly higher in grassland than in forest areas. This difference could be explained due to the low cover vegetation in open areas that may allow a major solar radiation and consequently upper temperatures (Pintanel et al. 2019; Scheffers et al. 2017), while the arboreal vegetation in forest habitats may act like a buffer that generates low temperatures (Pincebourde & Suppo 2016; Scheffers et al. 2017). Nevertheless, at the highest elevational zone studied (2,700 m.a.s.l), the difference of anuran BT between open and forest habitats was not statistically significant, even though the environmental temperature in open areas was slightly hotter than in forest habitats (Fig. 5). This similarity between open and forest environments at the highest elevation could be due to the fact that BTs of both studied species, Niceforonia adenobrachia and P. permixtus, were measured in equivalent microhabitat conditions (N. adenobrachia: under rocks, plants or soil vegetation; P. permixtus: on bushes or ground). As opposed to our hypothesis, the thermal variance of anuran BT from forest habitat was not statistically lower than that from open habitat for each elevational level studied (Fig. 4). We consider that this result can be attributed to the most heterogeneous landscape (and thus a slightly wider thermal variance) of the forest habitat where we recorded the anuran BT (on the ground, mosses, tree branches, tree leaves, in water bodies), in comparison to the open habitat that was most homogenous (leaf litter, rocks and the soil vegetation). It has been demonstrated that within a forest, the microclimates change according to features such as topography, landscape composition and local water balance (De Frenne et al. 2021).
Some species found at the same elevation in both forest and open habitats (Pristimantis taeniatus and R. margaritifera) had similar BTs (Table 1). Contrarily, B. platanera and R. margaritifera sampled along two elevational zones showed BTs more associated to their elevation than their habitat (Table 1). Consequently, variations in both microhabitat and elevational environmental temperatures are good predictors of BTs in anurans and then their knowledge is crucial to determine risks associated to climate changes and habitat loss, which are considered key threatening processes for the biodiversity, as they cause thermal stress and droughts (Mantyka et al. 2011). For instance, the rapid decline in a population of the salamander Aneides aeneus within a highly fragmented habitat has been linked with an increase in the environmental temperature (Corser 2001). This information is also particularly valuable in tropical habitats that are frequently exposed to rapid fragmentation and destruction (Alroy 2017), as many anuran species are adapted to the specific temperature of their microhabitats (Bernal & Lynch 2013), where narrow changes in these temperatures may seriously affect the survival of species (Turriago et al. 2015), some of them likely stenothermic (i.e., organisms that cannot tolerate wide ranges of environmental temperatures) (Navas et al. 2013).
CONCLUSION
In this work, we found that:
Substrate temperature (Ts) is notably a better predictor of tropical anuran BT than a close air temperature (Ta) (approximately 20 cm above anurans). This information is important to be more accurate in predicting the risk of amphibians due to the global warming as many forecasts are obtained from macroenvironmental temperatures rather than those from microenvironmental habitats (Navas et al. 2013; Pintanel et al. 2019).Anurans from high elevation (2,700 m.a.s.l), have a lower BT than those from lowlands (455 m.a.s.l), as expected by the temperature lapse rate. This result demonstrates the thermoconformity of anuran BT along elevational gradients in the Colombian Andes.Anurans from open habitats have a greater mean, although not necessarily a wider BT variation, than the species from forest habitats. These data show that the temperature of habitats can change even within the same elevational level, microhabitats, and thus, the anuran BT.
This reinforces the importance of the records of microhabitat temperatures to a better estimation of anuran BTs, and the physiological and behavioural effects of habitat modification (e.g., deforestation, habitat conversion) on these ectotherms. Finally, given that anurans from forest habitat have a lower mean BT than species from open habitat, it would be interesting to test if anurans from these habitats are more thermally specialists with a lower thermal tolerance than those from open areas (see Pintanel et al. [2019] for an example with Pristimantis frogs). This may have conservation implications considering the substantial heating found in converted tropical forest and threatened environments including tropical Andes (Senior et al. 2017).
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Angilletta MJ 2009 Thermal adaptation a theoretical and empirical synthesis New York Oxford University Press 10.1093/acprof:oso/9780198570875.001.1 · doi ↗
- 2Alroy J 2017 Effects of habitat disturbance on tropical forest biodiversity Proceedings of the National Academy of Sciences 114236056606110.1073/pnas.1611855114 PMC 546868428461482 · doi ↗ · pubmed ↗
- 3Angulo A Rueda J Rodriguez J La Marca E 2006 Técnicas de inventario y monitoreo para los anfibios de la Región Tropical Andina Inventory and monitoring techniques for amphibians of the Tropical Andean Region Bogotá DC Conservación Internacional
- 4Armesto LO Señaris JC 2017 Anuros del norte de los andes: Patrones de riqueza de especies y estado de conservación Northern Andean anurans: Patterns of species richness and conservation status Papéis Avulsos de Zoologia 573949152610.11606/0031-1049.2017.57.39 · doi ↗
- 5Bernal MH Lynch JD 2013 Thermal tolerance in anuran embryos with different reproductive modes: Relationship to altitude The Scientific World Journal 20131710.1155/2013/183212 PMC 367697323766678 · doi ↗ · pubmed ↗
- 6Blomberg SP Garland T Ives AR 2003 Testing for phylogenetic signal in comparative data: Behavioral traits are more labile Evolution 57471774510.1111/j.0014-3820.2003.tb 00285.x 12778543 · doi ↗ · pubmed ↗
- 7Bohlman S Matelson T Nadkarni N 1995 Moisture and temperature patterns of canopy humus and forest floor soil of a montane cloud forest, Costa Rica Biotropica 27131910.2307/2388898 · doi ↗
- 8Brattstrom BH 1963 A preliminary review of the thermal requirements of amphibians Ecology 44223825510.2307/1932171 · doi ↗