Leaf Architecture and Genome Size Variation of Durio zibethinus L. from Jelebu, Negeri Sembilan, Malaysia
Kamaruddin Shamin-Shazwan, Rozilawati Shahari, Che Nurul Aini Che Amri, Mohd Razik Midin

TL;DR
This study examines leaf features and genome size differences in durian plants from Jelebu, Malaysia, to aid in their classification and identification.
Contribution
The study provides new taxonomic data on durian accessions from Jelebu using leaf architecture and genome size variations.
Findings
Seven leaf parameters were identified to differentiate durian accessions from Jelebu.
Genome size varied between 1.7433 pg and 1.800 pg among the studied accessions.
Intraspecific variation was observed in leaf architecture and genome size.
Abstract
Durio zibethinus L. is known as the “king of fruit” in Malaysia. Meanwhile, Jelebu, Negeri Sembilan has always become the top choice district to visit for durian lover for its Durian Kampung Jelebu, which possessed good quality on par with top D. zibethinus clones such as D197 Musang King and D24. However, there is still lacking in taxonomic data of D. zibethinus especially from Jelebu. This study aimed to analyse the leaf architecture and genome size variations of selected D. zibethinus accessions from Jelebu. Five D. zibethinus accessions from Jelebu were examined. Thirty-seven parameters of gross leaf morphological characteristics and leaf venation pattern were observed and recorded for identification and classification of D. zibethinus accessions from Jelebu. Seven parameters have been recorded which are petiole length, petiole features, leaf size, leaf shape, leaf base shape, lower…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
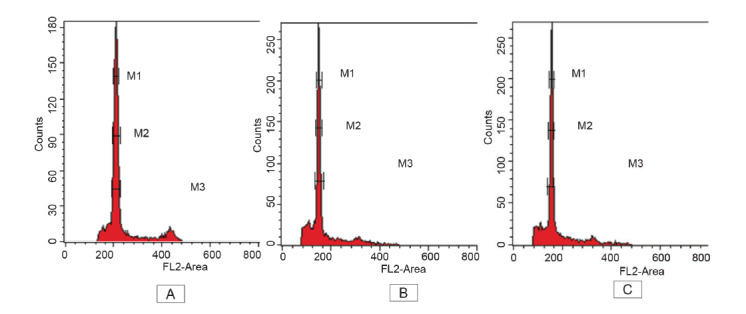 Figure 1
Figure 1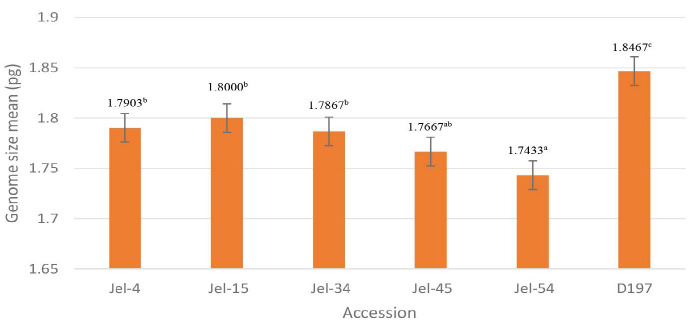 Figure 2
Figure 2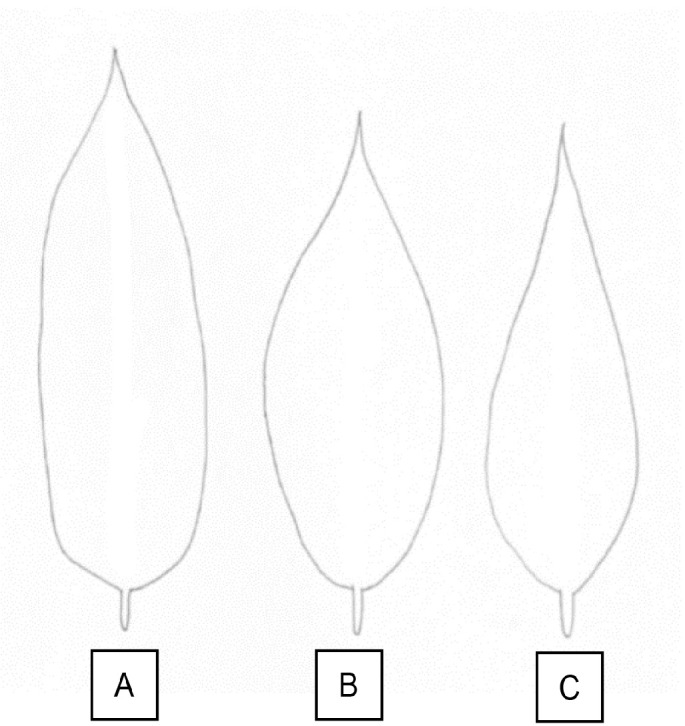 Figure 3
Figure 3 Figure 4
Figure 4 Figure 5
Figure 5 Figure 6
Figure 6- —Ministry of Higher Education
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsLeaf Properties and Growth Measurement · Plant Diversity and Evolution · Seed and Plant Biochemistry
INTRODUCTION
Durio zibethinus L., locally known as durian possessed high value for its delicious and unique aromatic fruit. Malaysia itself has recorded to have at least 200 registered D. zibethinus clones (Department of Agriculture Malaysia [DOA] 2020). D. zibethinus has gained popularity among researchers in determining the more potential possessed by the species in utilising it in various fields. Several previous studies have been conducted on D. zibethinus, including morphology, leaf anatomy, palynology, molecular study and its nutritional values.
Durio zibethinus leaves are alternately arranged with elliptic, oblong, or lanceolate. The leaves are coriaceous with acuminate apex and obtuse base (Soegeng-Reksodihardjo 1962). The surface of the upper leaves is glossy and velvety while its lower surface is silver-golden due to the overlapping peltate, and dense stellate hairs covered (O’Gara et al. 2004). Previous research on its micromorphological characteristics of its leaves was done by Salma (1999) recorded six different types of trichomes: one glandular trichome and five non-glandular trichomes. Recent publication by Talip and Shamsuddin (2019) reported the characteristics of anatomical structures of the cross-section observation (petiole, midrib, lamina and margin) of D. zibethinus.
Molecular characterisation on D. zibethinus has been widely carried out in several countries such as Indonesia and Thailand, however there is insufficient information on the molecular analysis of D. zibethinus in Malaysia. One of the most popular genetic studies on D. zibethinus is the study of genome that reported by Teh et al. (2017) has found that the genome size of D. zibethinus is about 738 Mb and found the pungent smell of D. zibethinus is caused by methionine gamma lyases (MGL) gene. Various previous research has been done on nutrient content of D. zibethinus as per 100 g of aril such as calories content in between 144 kcal to 153 kcal, 40 glycemic index, 27.1 g of carbohydrates, 3.4 g of fat, 3.8 g of fibre, and 2.7 g of protein (Jennings 2019; DOA 2013; Siddiq & Nasir 2012; Morton 1987).
However, there is lack of information on the taxonomic data of D. zibethinus, especially in Jelebu, Negeri Sembilan, Malaysia. Jelebu has been hailed as one of the best hotspots for durian lovers in every fruiting season and is usually known as Durian Kampung Jelebu. Jelebu has been recognised as “Fruit Valley” by Farmers’ Organisation Authority (Lembaga Pertubuhan Peladang, LPP) and the Ministry of Agriculture and Agro-based Industry, and these government sectors are working together to develop Durian Kampung industries in Jelebu, Negeri Sembilan (Bernama 2018). Therefore, more research needs to be done on Durian Kampung Jelebu to expand the industry of Durian Kampung. One of the most crucial data needed for this effort is providing taxonomic data such as morphological databases for D. zibethinus accessions from Jelebu.
Morphological characterisation is one of the most important elements used by taxonomists in identifying and classifying the plant species for systematic studies. It is the easiest method that can be performed by observing significant characters on the roots, tree habit, leaf, flower and fruit. Today, morphological characterisation has led to more profound and accurate plant identification and classification studies in phenotypic characterisation through plant genetic analysis. Characterisation of D. zibethinus using vegetative structure, such as leaves part could help farmers or D. zibethinus growers identify between clones at the seedling level. In addition to that, these data are achievable, fast and reliable.
Currently, genetic diversity has become one of the important tools to be used in the identification and classification of plant species. Siew et al. (2018) reported that the morphological characteristics of D. zibethinus can be easily changed due to its phenotypic plasticity, which directly affects environmental factors such as climate change, nutrient, and moisture content of soil. Due to this, genetic diversity can be studied as an added value for taxonomic data to reduce the limitation of phenotypic plasticity. Besides, more research is needed through genetic diversity analysis based on genome size to support the identification and classification procedure by analysing the variations in genome size among D. zibethinus accessions.
Therefore, the aim of this study is to examine leaf architecture and genome size variations between selected D. zibethinus accessions from Jelebu. The hypothesis of the study is there might be difference in term of leaves morphological characteristics either in external morphological or micromorphological such as venation pattern that might becoming a significant data in identifying and characterising D. zibethinus accessions from Jelebu. Next, the genome size variations analysis might show variations in between accessions and this data will be beneficial in recognising the D. zibethinus Jelebu accessions.
MATERIALS AND METHODS
Leaves Sampling
The collection of leaves samples was collected in December 2021 from durian orchards in Jelebu, Negeri Sembilan. Five accessions of D. zibethinus accessions from Jelebu were selected for this study as tabulated in Table 1. The leaves samples were then processed into herbarium specimen by following standard herbarium protocol procedure with some modifications (Seshagirirao et al. 2016) and deposited in Herbarium Room, Department of Plant Science, Kulliyyah of Science, International Islamic University Malaysia, Kuantan, Malaysia. Young leaves samples from each selected accession were preserved in a freezer to avoid damage prior before the genome size estimation procedure by flow cytometry (FCM) analysis.
Leaf Morphological Analysis
Data collection for leaf architecture analysis was divided into two parts which are: gross morphological characteristics and leaf venation variations. Firstly, gross morphological characteristics data was collected from the observation of petiole structure, leaf structure and leaf surface. Quantitative data for length was measured by using ruler and angle was measured using protractor.
Secondly, leaf venation variations analysis was undergone sample preparation before observation by adapting simple preparation method by Bulger (2017) with some modifications. Matured and fully developed leaves of each accession were fully soaked under tap water for four to six months, depending on the thickness of leaf epidermis. The leaf was brushed off carefully to remove the epidermis layer and the tap water was replaced every three days until leaf epidermis can be brushed off until clear transparent leaf was obtained. observed and examined under a dissecting microscope. Description of the leaf architecture was followed terminologies by Ellis et al. (2009). Colour chart of the leaf surface was determined by several colour chart from Scheme colour (Shiny Silver Colour Scheme, https://www.schemecolor.com/shiny-silver-color-palette.php), Scheme colour (Light Gold Gradient Colour Scheme. https://www.schemecolor.com/light-gold-gradient.php), and Scheme colour (Green Scale Colour Scheme, https://www.schemecolor.com/green-scale.php).
Genome Size Estimation Analysis
Nuclei suspension preparation
Nuclei were extracted by chopping the young leaf sample in the suitable lysis buffer. LB01 buffer was selected during optimisation process prior to FCM measurement. RNase A and propidium iodide were then added into the samples and the mixture was incubated for 10 min prior to analysis.
The fluorescence intensity of each sample was measured by FCM machine equipped with 15 mW argon ion laser at 488 nm. Histograms were collected over 1,024 channels and for leaf samples 15,000 events were captured.
Genome size measurement
Measurement of genome size was then obtained using Glycine max cv. Polanka (2C = 2.5 pg, seeds were provided by J Doležel, Olomouc University) as an external reference standard. The values of fluorescent intensity peaks and genome sizes of samples were analysed using CellQuest3R software.
Data analysis
Genome size variations were analysed by using SPSS software to study the relationship between samples. Intraspecific species variations between selected D. zibethinus accessions were calculated according to Duncan’s multiple range test.
RESULTS
Leaf Architecture Observation
Based on this study, 37 parameters of leaf architecture were observed and recorded.?
Genome Size Estimation
FCM DNA histogram analysis
The LB01 lysis buffer used in this study generated a good DNA histogram peak with low CV value which is less than 5% and minimum debris background for all accessions (Fig. 1). Glycine max cv. Polanka was used as an external standard as it has established genome size, 2.5 pg and widely used in FCM analysis (Kandaiah et al. 2021; Temsch et al. 2021; Midin et al. 2020). Midin et al. (2020) reported that the DNA histogram peak of G. max was not overlapping to the peak of Garcinia mangostana, determining the suitability of G. max to be used as external reference standard in their study. Fig. 1 demonstrates DNA histogram of G. max, D. zibethinus clone D197, and D. zibethinus Jel-15 which having the lowest CV value (Table 3) and fine peak area of histogram peak among other accessions. Based on the result, DNA peak of G. max nuclei intensity peak was located between channel 200–240 while Jel-15 and D197 was located on the channel 120–160 indicated intraspecific genome size variation.
Genome size determination
Table 3 shows genome size and coefficient of variation-first time used value (CV) for each D. zibethinus accessions. CV value is used to measure and compare the variation between samples among quantitative traits as it is important in understanding the phenotypic plasticity and its evolvability (Pélabon et al. 2020). The bar chart in Fig. 2 illustrates the genome size of each accession for this study. The value on the top of each bar represents the value of genome size of D. zibethinus accessions. Based on statistical analysis used, Duncan analysis shows there are four variations obtained which are 1-a group: Jel-54, 2-ab group: Jel-45, 3-b group: Jel-4, Jel-15, and Jel34, and 4-c group: D197 (Table 3).
DISCUSSION
Leaf Architecture
Leaf architecture observations were divided into four mains parts which are leaf attachment, leaf structure, leaf surface and leaf venation.
Leaf attachment
All accessions shared common characteristics by having petiole, alternate arrangement, and position of petiolar attachment is at marginal. The similarities exist proved all five accessions belong to the same D. zibethinus species. These characteristics are common and like all D. zibethinus accessions as reported by Effendi (2013).
There are variations in petiole length between accessions. Jel-4, Jel-34 and Jel-45 were recorded to have short petiole with less than 1.80 cm while Jel-15 and Jel-54 has long petiole more than 1.80 cm. Differences in petiole length could determine the variations among D. zibethinus existed. Length of petiole is also observed by other researchers for various identification and classification purposes especially in taxonomic study. For example, Ackerfield and Wen (2002) have used petiole length as one of the parameters in morphometric analysis between Hedera L. species. In addition, Hussain et al. (2008) mentioned petiole length is heritable and found that petiole should become the parameter for cotton breeding research in improving crop quality. This study also found all D. zibethinus accessions from Jelebu have similar characteristics with rounded petiole shape. Rounded petiole shape is also observed by Effendi (2013) study on two leaves of D. zibethinus clone Sunan and Brongkol.
Leaf structure
Leaves of all accessions are simple and symmetric as observed and reported by Idris (2011). Simple leaf organisation is one common characteristic of D. zibethinus. The leaf organisation parameter is also used by Mishra et al. (2010) in identifying Coffea arabica cultivar whereas they also found all cultivars also shared the same characteristics of simple leaf structure.
Variations of leaf size were observed for all accessions. This study found that the leaf length varies between 10.90 cm and 14.87 cm and width varies between 3.67 cm and 4.93 cm. Leaf size is important for identification of species since each species possessed unique characteristics. Further leaf length and width classification has been done based on the data of this study. Jel-15 is the only accession with a short leaf, while the others are long leaves and Jel-4 is the only accession with a broad leaf in contrast to the other accessions with narrow leaf width. Ardiyani (2015) has utilised leaf size of newly found species of Zingiberaceae for the species identification and Liu and Hong (2016) are used leaf size data as one of dichotomous key parameters in revising taxonomic data of Pourthiaea villosa. These two studies proved that leaf size is significant in species identification and classification that can be applied on D. zibethinus accessions under study. There are three main variations of leaf shape which are ovate, elliptic, oblong as illustrated in Fig. 3. Jel-15 and Jel-34 have elliptic leaf shape, and Jel-45 with ovate shape. However, there is a unique variation on Jel-54 with the only accession that shows variation in its leaf shape with elliptic to oblong shape. These characteristics have also been observed on several D. zibethinus accessions (Pratiwi et al. 2018). Thus, leaf shape characteristics possessed significant value in taxonomic study in early identification and classification of D. zibethinus accessions.
All accessions recorded to have acuminate shape with less than 90° apex angle (Fig. 4). However, variations exist for base shape characteristics of D. zibethinus. Three types of leaf base observed were acute, obtuse, and rounded with the same base angle of more than 90° (Fig. 5). Only Jel-34 is observed to have an acute to obtuse base, Jel-4 is the only accession with acute base, Jel-15 possessed obtuse leaf base and Jel-45 and Jel-54 with rounded base. Based on this study, Jel-34 showed two types of base shape. Apex and base characteristics might have value on plant classification as these plant parts are used also by Hernandez et al. (2020) for analysing leaf architecture variations existed among Dipterocarpus species. Based on the study, they highlighted base shape as one of significant parameters to be used in taxonomic study.
All accessions observed also to have entire and unlobed margin type. These two parameters are also important to be studied in identifying D. zibethinus as they might provide data for greater identification to find variations between accessions. However, this study found no variation in the margin characteristics and the findings are similar to the previous study done by Effendi (2013). Thus, these parameters could be considered as common characteristics of D. zibethinus accessions.
Leaf surface
All accessions have green shiny on the upper surface. This is common for all D. zibethinus accessions from Jelebu and is easy to identify the species by early observation and the same observation reported by Talip and Shamsudin (2019). Other than that, all accessions are also densely covered by trichomes on the lower side of the leaves. The presence of dense trichome on the lower surface of the leaves is one of significant characteristics possessed by every D. zibethinus accessions which also mentioned by Talip and Shamsudin (2019), and Effendi (2013).
There is a variation on the colour of the lower surface of the leaves, either silvery grey or golden brownish in colour. Characteristics of leaves lower surface colour is also used by Sundari (2015), and they found three different colours: greenish white, beige and brown. Apart from D. zibethinus, leaf surface colour is also used by Shan et al. (2019) in analysing the taxonomic value of leaf colour traits for Eriobotrya.
Leaf venation
From observation in Table 1 and Fig. 6, all five accessions shared the same characteristics on the leaf venation characteristics. This study found the data obtained is the same as previous research reported by Laraño and Buot Jr. (2010) which also indicated that collected taxonomic data from this research is relevant to be used in identifying and classifying D. zibethinus accessions. Similarity in leaf venation characteristics among accessions might indicate strong genetic relationship within D. zibethinus species that also mentioned by Effendi (2013). Leaf venation is also used in taxonomic study of other species such as Psidium species where this observation provided morphological classification data (Oliveira et al. 2017).
Areolation is referring to the smallest areas that present around leaf tissue that are surrounded by major leaf veins and form a contiguous field over most of the leaf area (Mishra et al. 2010). This study found Jel-4, Jel-15 and Jel-54 have well developed areolation while Jel-34 and Jel-45 areolation is moderately developed. This variation might be useful in further classification of D. zibethinus. Lastly, all accessions observed to have two or more branched of FEVS and looped MUV. These characteristics could be used as supporting data in identification and classification of D. zibethinus accessions.
Genome Size Variation
In this study, intraspecific genome size variations were revealed among all six selected D. zibethinus accessions used in this study. This study found that D. zibethinus genome size ranges between 1.7433 pg to 1.8000 pg. It shows the difference range between accessions is closer from one another and it is expected all selected accessions under study are confirmed comes from the same D. zibethinus species.
D. zibethinus Jel-15 recorded the highest genome size with 1.8000^b^ ± 0.006 pg among D. zibethinus accessions from Jelebu while Jel-54 was the lowest with 1.7433^a^ ± 0.009 pg (Fig. 2). However, D. zibethinus clone D197 Musang King was observed to have the highest genome size mean with 1.8467^c^ ± 0.009 pg in comparing to all five D. zibethinus accessions from Jelebu. Genome size estimation analysis of selected D. zibethinus accession obtained positive result with 0.001 significant level that determine each accession is different from one another.
The crucial of utilising genome size analysis on plant species could be observed from Christia vespertilionis, which Ibrahim et al. (2022) has reviewed various previous research that research on this plant species for its ethnomedicine purposes. In addition, they proved also it has potential to be used in modern medicine as material for anti-cancer activity, anti-malaria activity and anti-inflammatory activity. Taxonomic study of this species via various methods are needed and important to recognise the plant to avoid confusion of plant identification from other Christia species. Midin et al. (2017) has successfully incorporated the application of the genome size estimation of C. vespertilionis species that could be used as additional taxonomic data for the species identification. FCM was also used by Jarret et al. (1995) in characterising genome size variations of Paspalum germplasm. They reported this technique is fast, precise, and sufficient to differentiate individual accessions between Paspalum accessions. These previous studies observed the occurrence of intraspecific among C. vespertilionis and Paspalum species.
Intraspecific Variations
Cytogenetics also refers to the study of chromosomes via microscopy technology leads to the understanding of the genome of each plant. This is reliable technique to be used for analysing intraspecific variations among D. zibethinus accessions from Jelebu as chromosomes consists of genetic materials that least affected to the environment conditions, the study on the chromosome’s behaviour, structure as well as its function could assist in understanding genetic characterisation (Spinner & Ferguson-Smith 2019). Intraspecific variation is referring to the variation of plant within a species. Intraspecific variation study is common to obtain low characteristics variations since the study is focusing on less population than interspecific variation study (Hahn & Maron 2016). Based on a report by Hahn and Maron (2016), it is safe to conclude all D. zibethinus under study could be categorised as intraspecific variations study due to narrow genome size variations.
The occurrence of intraspecific variations is reported by Smarda and Bures (2006), and Cavallini and Natali (1991). Smarda and Bures (2006) reported the occurrence of intraspecific genome size variations between Festuca pallens varieties, and they observed correlation between genome size variations to macroecological, geographical and evolutionary factors. The finding of this study might have influenced from the environment or ecological factor which interesting to be analysed in the future to provide more data for understanding intraspecific variations among D. zibethinus accessions. This could be seen also from the grouping by using Duncan’s analysis, that there is no pattern of distribution accessions between clusters. By referring to Table 1, the b-group that consists of Jel-4, Jel-15 and Jel-34 are collected from three different orchards, and still arranged under same group. However, genome size variations could still be used in genetic characterisation of D. zibethinus accessions and possessed valuable data for determining taxa rank.
The understanding of intraspecific variations among D. zibethinus accessions provides taxonomists with sufficient information for the evolutionary theory and ecological conditions of the accessions. It also served as additional data for analysing the phenotypic differences between accessions. This analysis also supported Roches et al. (2018) statement on observing the ecological importance of variations and its availability for each species. It is clearly shown significant of intraspecific variations data for showing next potential research in the future. Data on intraspecific variations of D. zibethinus accessions can be additional taxonomic data for the identification and classification of accession for the registration of new clone in the future, supporting fruit morphological characteristics and DNA identification database.
CONCLUSION
In conclusion, this study highlights the significant value of leaf architecture and genome size data in taxonomic study of D. zibethinus accession from Jelebu and serve as additional information to avoid confusion for D. zibethinus growers as well as researchers in identifying and classifying D. zibethinus accessions. In addition, genetic diversity was potentially supporting data in determining taxa rank for unidentified D. zibethinus accessions. Thus, the finding of this study could become an important reference for future D. zibethinus breeding programme and fruit improvement study. A detailed taxonomic study incorporated several taxonomic data such as leaves morphological characteristics, leaves anatomical characteristics, genome size, and its fruit morphological characteristics must be done to provide more complete taxonomic data of D. zibethinus accessions from Jelebu.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Ackerfield J Wen JA 2002 Morphometric analysis of Hedera L. (the ivy genus, Araliaceae) and its taxonomic implications Adansonia 242197212
- 2Ardiyani M 2015 A new species of Zingiber (Zingiberaceae) from Enggano Island, Indonesia Reinwardtia 14230731010.14203/reinwardtia.v 14i 2.1676 · doi ↗
- 3Bernama 2018 LPP identifies potential area for durian cultivation https://www.malaymail.com/news/malaysia/2018/01/10/lpp-identifies-potential-area-for-durian-cultivation/1550837 accessed on 8 June 2020
- 4Bulger N 2017 Botanical project: Leaf skeletons (non toxic) https://www.naomiloves.com/blog/2017/11/19/how-to-make-leaf-skeletons-non-toxic accessed on 10 November 2020
- 5Cavallini A Natali L 1991 Intraspecific variation of nuclear DNA content in plant species Caryologia 4419310710.1080/00087114.1991.10797023 · doi ↗
- 6Department of Agriculture Malaysia (DOA)2020 Garis panduan kebangsaan untuk menjalankan ujian kelainan, keseragaman dan kestabilan http://pvpbkkt.doa.gov.my/Test%20Guidelines/Durian.pdf accessed on 14 July 2020
- 7Department of Agriculture Malaysia (DOA)2013 Durian Pulau Pinang Department of Agriculture http://jpn.penang.gov.my/index.php/perkhidmatan/teknologi-tanaman/buah-buahan/64-durian-sp-26113 accessed on 10 November 2020
- 8Effendi LN 2013 Identifikasi morfologi durian Durio zibethinus Sunan dan Brongkol dalam penyusunan basis data keragaman Masters diss Universitas Sebelas Maret Indonesia