Morphological Characterisation of Three Populations of Heterobranchus longifilis from Nigeria
A. Alih Raphael, Solomon Shola Gabriel, Olufeagba Samuel Olabode, O. Cheikyula Joseph, Abol-Munafi Ambok Bolong, Mhd Ikhwanuddin, Okomoda Victor Tosin

TL;DR
This study examined the morphological differences of Heterobranchus longifilis from three Nigerian regions but found no clear distinction using physical traits alone.
Contribution
The study demonstrates that morphological traits cannot reliably distinguish H. longifilis populations from different ecoregions in Nigeria.
Findings
Univariate analysis showed some significant differences in morphometric parameters.
Multivariate analysis failed to separate the populations into distinct clusters.
Molecular markers are suggested for further population characterization.
Abstract
This study attempted to discriminate the population of Heterobranchus longifilis in Nigeria using their morphological characteristics. Therefore, 60 sexually mature wild samples of H. longifilis (1:1 for the male and female ratio) of relatively similar size (40 cm) were collected from three eco-regions namely, Guinea Savanna (Benue River, Makurdi), Rainforest Savanna (Niger River, Onitsha) and Sahel Savanna (Rima River, Sokoto). They were transported to the hatchery unit of the Fisheries and Aquaculture Department, Joseph Sarwan Tarka University Makurdi where the morphometric data was collected. The data for 39 traditional morphometric measurements and 5 meristic counts obtained from each fish were subjected to univariate and multivariate analysis. While significant differences were observed in some parameters following univariate analysis; it was revealed that the morphometric…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
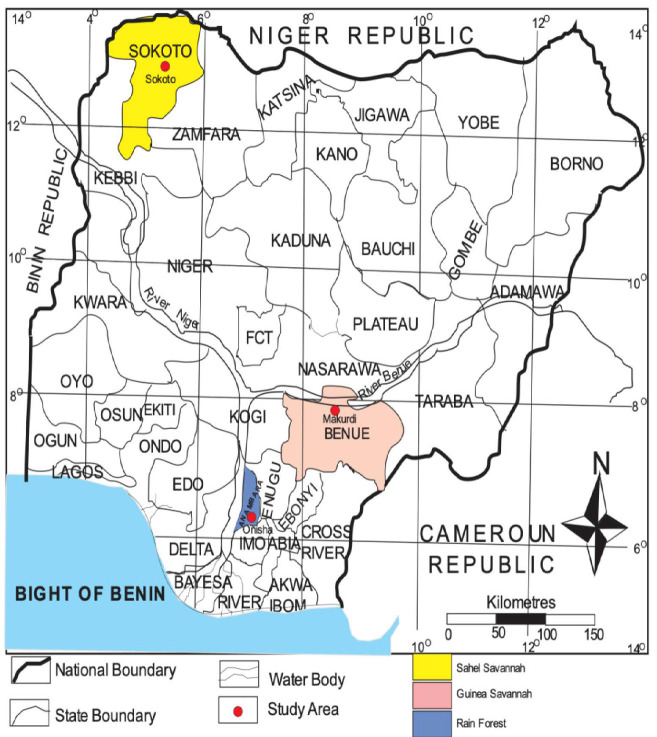 Figure 1
Figure 1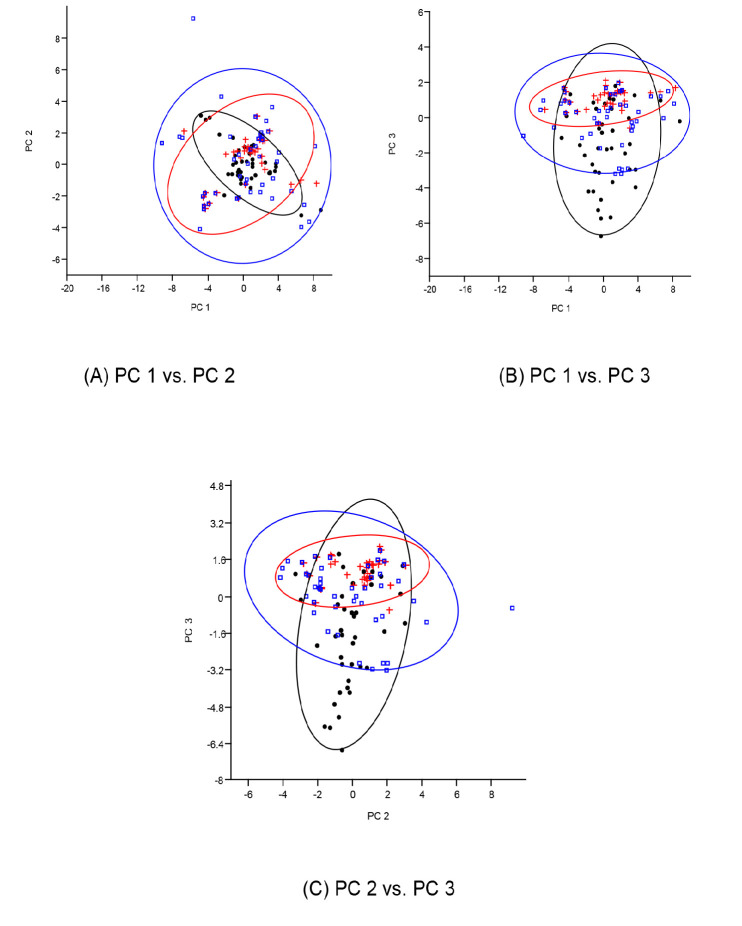 Figure 2
Figure 2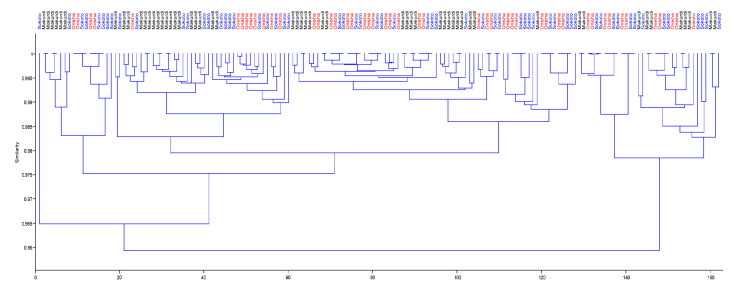 Figure 3
Figure 3 Figure 4
Figure 4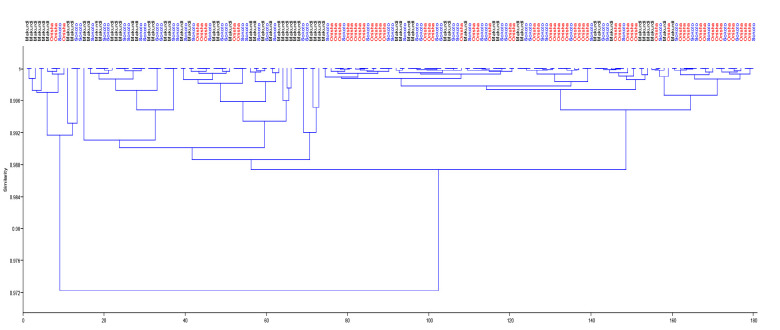 Figure 5
Figure 5Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsFish Biology and Ecology Studies
INTRODUCTION
The genus Heterobranchus is one of the most important freshwater fish genera in sub-Saharan Africa (Ataguba et al. 2009). However, on-like the genus Clarias which has been extensively studied at various levels under different aquaculture disciplines around the world, research on the species of the genus Heterobranchus is still localised within the environs of Africa (Alih et al. 2022). Although, several fundamental studies earlier conducted on species such as H. longifilis have led to the improvement of breeding, and aquaculture production under captivity (Baras 1999; Poncin et al. 2002; Ataguba et al. 2009; Olufeagba et al. 2016; Okomoda et al. 2017); there are still research gaps as regards to information about morphological variation between the different population of the fish. Such studies in conjunction with genetic characterisation can be the basis upon which a selective breeding program could be conducted to improve the cultural performance of its progenies (Solomon et al. 2015).
According to Normala et al. (2017), conventional morphological methods continue to have an important role in stock identification despite the development of advanced techniques that can directly examine biochemical or genetic variations. Morphological and biometrical characteristics remain the simplest and most rapid methods used in delineating, discriminating and classifying fish stocks/identifying species (Hockaday et al. 2000; Turan et al. 2004). The study of the variability of morphological characters of fish is important to inform subsequent genetic studies (Olufeagba et al. 2015a, 2015b; Okomoda et al. 2018). The phenotypic plasticity of a population can therefore be used to distinguish between different stocks (O’Reilly & Horn 2004). This is because it is largely not under genetic control alone but influenced by environmental conditions (Chittenden et al. 2010; Arora & Julka 2013).
Fish population responses to environmental changes involve the modification of physiological and behavioural characteristics. This consequently affects the morphological and reproductive status as the fish attempts to mitigate the effect of the environmental changes experienced (Turan 1999). This means the characteristics of the different populations are shaped/impacted by the environmental conditions where each population lives. Thus, genetic information alone is not sufficient in choosing the base population for a selective breeding program but an interaction between genetics and environmental factors which are expressed in different morphological changes (Scheiner & Callahan 1993; Hoffman & Merila 1999). Since different habitat experiences different environmental changes, therefore different impacts are experienced by different population structures (Ahmad 2015). These habitat-specific environmental conditions may include such factors as predation pressure, food availability, salinity, temperature, turbidity and water condition (Scapini et al. 1999; Maltagliati et al. 2003; Remerie et al. 2005). The current study was therefore designed to determine the extent of morphological variation of three populations of H. longifilis from three eco-regions in Nigeria. This is considered the first step toward developing a selective breeding program for fish species in captivity (Okomoda et al. 2022).
MATERIALS AND METHODS
The populations of H. longifilis for this study were obtained from three eco-regions of Nigeria (Fig. 1). The first is the Sahel Savanna precisely from Rima River, in Sokoto State located at latitude 13.0059°N, longitude 5.2476°E with an annual mean temperature of 28.30°C and rain ranges between 500 mm and 1,300 mm. H. longifilis samples were also gotten from the Guinea Savanna at the Benue River in Makurdi, Benue State located at latitude 7.7322°N, longitude 8.5391°E. This region has a mean annual temperature of 26.7°C and a mean annual rainfall of 1,077 mm. The place for the fish collection was the Rainforest region at the Niger River, in Onitsha with latitude 6.1329°N and longitude 6.7924°E. It has an annual mean temperature of 27.0°C and a mean annual rainfall of 1,828 mm (https://en.climate-data.org).
A total of 60 fish samples of reproductive age (i.e., 1:1) and relatively similar sizes of about 40 cm were collected from each eco-region for two months (i.e., the collection was done weekly). Hence, a total of 180 fish samples were collected from the three eco-regions. The fish samples were identified using the identification keys adopted by Moses and Olufeagba (2009) following confirmation from the local fishermen before transporting live from the eco-regions to Makurdi in 50 L open black plastic jerry-cans equipped with continuous aeration (using battery-powered motors). Upon reaching the research farm, morphological parameters were determined as shown in the next section.
Morphometric and Meristic Measurements
The morphometric and meristic characterisation in the current study were according to the previous method described by Okomoda et al. (2018). Data for 39 morphometric measurements and 5 meristic counts were taken from each sample of H. longifilis collected from the three eco-regions of Nigeria, using a measuring board, meter rule, and a weighing balance. This includes body weight (BW measured in grams), head length (HL), standard length (SL), total length (TL), dorsal fin length (DFL), adipose fin length (ADFL), head width (HW), eye diameter (ED), inter-orbital distance (IOD), body depth (BD), occipital fontanelle length (OFL), occipital fontanelle width (OFW), vomerine width (VW), dorsal fin height (DFH), predorsal length (PDL), pectoral fin length (PeFL), prepectoral length (PPcL), pelvic fin height (PFH), pelvic fin length (PFL), anal fin ray number (AFRN), anal fin length (AFL), pelvic fin to anal fin (PvFAF), caudal fin ray number (CFRN), snout length (SnL), nasal barbell length (NBL), maxillary barbell length (MxBL), premaxillary length (PmxL), vomerine length (VL), vomerine gap (VG), dorsal fin ray number (DFRN), anal fin height (AFH), caudal fin length (CFL), caudal fin height (CFH), and caudal peduncle depth (CPdD).
Data Analysis for Morphological Parameters
The measurements taken were first standardised to remove the effect of size before analysis was done following the method adopted by Murta et al. (2008) and Jaferian et al. (2010). By doing so, the individuals from each sample collection were normalised into a single arbitrary size, while maintaining individual variation within the sample (Sen et al. 2011). The relation used to achieve this was the allometric formula described by Elliott et al. (1995):
where M = observed character measurement, M**adj = size-adjusted measurement, L**o = standard length of the fish, L**s = overall mean of the TL for all the progenies, and b = estimated for each character from the collected data as the slope of the regression of log M on log L**o, using all fish of all the progenies.
Upon transforming the data, only data from 162 individuals of the 180 samples collected were used for the multivariate analysis of the Principal Component Analysis (PCA) using PAST free software. The exclusion of some samples from the analysis was due to incompleteness in the data entry due to human errors. The PAST free software was also used to obtain a sample centroids graph on the biplot, which then allows the determination of the most valuable morphological character that can be used to separate the fish groups into distinct multivariate spaces. Dendrograms with complete linkage and Euclidean distances of the fishes were also determined using the PAST free software and reported accordingly.
RESULTS
The result of the present study based on the univariate analysis of variance (ANOVA) reveals similarities in most of the morphometric parameters except for 13 parameters namely: CPL, PrPel, PFW, PFDT, PSL, PFL, DFPF, LML, VW, SnL, PrML, MBL and NBL (Table 1). The Onitsha population had higher values in 9 of the 13 parameters (P < 0.05) while in most cases, Makurdi and Sokoto were similar and had the least values. Expressing the morphometric parameters as percentages of standard length, however, did not change the trend of the result shown in Table 2. The result for the PCA for the transformed homologous morphometric parameters of the H. longifilis (Table 3) only used the first three principal components (PC), as the eigenvalue was more than 1. This is according to the recommendations earlier made by Kaiser (1961).
For the morphometric parameters, the first principal component (PC1) accounted for 28.49% of the total variance with only negative coefficients. The second principal component (PC2) had a mix of positive and negative coefficients and counted for 9.43% of the total variance in this study. In the same vein of mixed positive and negative coefficients, PC3 accounted for just 8.33% of the total variation. In summary, all three principal components accounted for only 46.25% of the variance observed for the H. longifilis from the ecoregions. The low cumulative variance did not permit the recommendation of influential variables as the different populations could not be separated into unique multivariate spaces as seen in the biplot in Fig. 2. Similarly, the dendrogram of complete linkage and Euclidean distance shown in Fig. 3 also showed multiple overlaps of the samples from the different ecoregions.
The result of the univariate analysis of variance (ANOVA) of the meristic counts revealed significant differences in four of the five parameters (with the exception of PFRC). The Onitsha strain had significantly higher counts compared to the Sokoto population, while the least values were gotten from the Makurdi population (P < 0.05). The PCA for the transformed homologous meristic counts of the H. longifilis samples presented in Table 4 showed a cumulative variance of 76.22% for PC1 (55.51%) and PC2 (20.71%) as they had eigenvalue above unity (1). While the PC1 contained only positive coefficients, the PC2 had mixed positive and negative coefficients. The biplot shown in Fig. 4 further showed that the population could not be separated into unique multivariate spaces using the meristic counts, hence, influential variables would largely not be accurate. Also, the dendrogram of complete linkage and Euclidean distance shown in Fig. 5 showed multiple overlaps of the samples from the different ecoregions using the meristic counts.?
DISCUSSION
The importance of obtaining detailed knowledge of the population structure of commercially exploited fish species cannot be overemphasised as it allows for the efficient management of the fisheries. It is the prerequisite for any genetic improvement programme if it is to be successful (Okomoda et al. 2022). Morphological characteristics such as morphometric parameters and meristic counts are commonly used to identify stocks of fish, populations, and species (Turan et al. 2004). It relies on the detection of subtle differences in shapes independent of size to discriminate populations within a given species. Generally, the variability observed or measured encompasses all aspects of the phenotypic variables (Tave 1993). Our study showed significant differences in 13 of the 39 morphometric parameters measured and in four of the five meristic counts (with the exception of PFRC). The study by Ajado and Edokpayi (2003) had earlier reported significant differences in the number of dorsal rays and the gill raker count in their study of two populations of the Clariid Clarias gariepinus population from Delta and Lagos. Similarly, Agbebi et al. (2009) reported significant differences in some parameters of H. bidorsalis with higher values linked to the population from Gboko compared to those from Onitsha and Jos. The differences in the reports of these studies could be linked to the difference in species, sizes and the number of samples used for the various studies.
Those intra-specific morphological differences noticeable in different population structures are usually not directly under the control of the genes but are subjected to environmental modification (Clay 1977). This is because fishes are the most susceptible vertebrate to environmentally induced morphological variations; hence, they demonstrate greater variance within and between populations than any other vertebrate (Solomon et al. 2015; Okomoda et al. 2018). Betiku (2006) suggested that the mechanism of action resulting from these includes quick adaptation and modification of physiological and behavioural states in response to environmental changes, therefore, modifying the fish’s morphology significantly. Hence, the non-discrimination of various populations in our current study into unique multivariate spaces (using the meristic and morphometric parameters) might be due to the low degree of environmental impact as well as the low level of adaptation of the population at the time of the study was conducted. This was demonstrated in our earlier study with different populations of wild and cultured Anabas testudineus in Malaysia (Okomoda et al. 2022). That previous study showed that the cultured population was unique and distinct, while an extensive overlap characterised the wild populations suggesting a similar origin of the stock. This, therefore, strengthens the narrative that the differences in the degree of environmental impact and level of adaptation of the population could have dictated the level of morphological plasticity observed in different fish species.
Due to the overlap of the different populations, this study could not suggest the most influential parameters for the discrimination of the fish stocks. This contrasts with the findings of Solomon et al. (2015) who suggested head length, BDA, and eye diameter as the most influential morphometric parameters for the discrimination of cultured and wild African catfish C. gariepinus in Nigeria. Haddon and Willis (1995) also stated that morphometrics measurements of the head and the body depth were the most important characters in the identification of the population of the Angler fish, Lophius vorernus, Pacific herring, Clupea pallasi and Orange roughy Hoplostethus atlanticus. Aside from the influence of the changes in environmental factors (Turan et al. 2005; Solomon et al. 2015); differences in geographical and ancestral origin (Hossain et al. 2010), phenotypic variation in natural stock sometimes reveals genetic adaptation to selection pressures (Solem et al. 2006). Therefore, the outcome of studies is most likely evidence of the possible combination of genetic and environmental factors as it influences the morphology of the fish (Olufeagba & Yisa 2003). Hence it is therefore important to initiate genetic studies of the fish from these three eco-regions to determine the levels of genetic variations despite the observable morphometric similarities.
CONCLUSION
Although morphological characterisation has been reported in many previous studies to be useful in the discrimination of fish stock, the current study has demonstrated otherwise when considering three Nigerian populations (namely, Makurdi, Onisha, and Sokoto population). While the exact reason for this deviant observation may not be well understood, genetic studies are urgently needed to determine the level of variability among the population should selectively breeding of the fish for improvement be considered. Future studies can also consider the morphometric comparison of wild and cultured H. longifilis as many years of indiscriminate breeding could have led to unplanned backcrossing of hybrids with pure crosses.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Agbebi OT Ajagbe O Makinde L Afolabi O 2009 Production, growth and effect of varying stocking density of Clariobranchus fry Journal of Fisheries International 44737810.3923/jfish.2009.73.78 · doi ↗
- 2Ahmad ABM 2015 Genetic and morphometric variability assessment of wild and captive populations of the Climbing perch, Anabas testudineus towards development of a selective breeding programme Ph D diss Universiti Malaysia Terengganu
- 3Ajado EO Edokpayi CA 2003 Comparative racial study of Clarias gariepinus (Burchell, 1822) from River Niger and Badagry Lagoon, Southwest Nigeria Nigerian Journal of Fisheries 14148
- 4Alih RA Solomon SG Olufeagba SO Cheikyula JO Abol-Munafi AB Okomoda VT 2022 Breeding performance and embryogenic development of three strains of Heterobranchus longifilis in Nigeria Zygote 30112513110.1017/S 096719942100041134176523 · doi ↗ · pubmed ↗
- 5Arora R Julka J 2013 Phenotype and genotype differentiation between two stocks of Tor putitora (Hamilton) population (Pisces: Cyprinidae) from Himachal Pradesh, India International Journal of Plant, Animal and Environmental Sciences 33141
- 6Ataguba GA Annune PA Ogbe FG 2009 Induced breeding and early growth of progeny from crosses between two African clariid fishes, Clarias gariepinus (Burchell) and Heterobranchus longifilis under hatchery conditions Journal of Applied Biosciences 14755760
- 7Baras E 1999 Sibling cannibalism among juvenile vundu under controlled conditions. I. Cannibalistic behaviour, prey selection and prey size selectivity Journal of Fish Biology 5418210510.1006/jfbi.1998.0847 · doi ↗
- 8Betiku OC 2006 Characterization of wild and domesticated Clariid strains using polyacrylamide gel electrophoresis (PAGE)Masters’ diss Department of Animal Sciences, Obafemi Awolowo University, Ile-Ife