Activity in the pontine reticular nuclei scales with handgrip force in humans
Tyler L. Danielson, Layla A. Gould, Jason M. DeFreitas, Rob J. MacLennan, Chelsea Ekstrand, Ron Borowsky, Jonathan P. Farthing, Justin W. Andrushko

TL;DR
This study shows that brain activity in the pontine reticular nuclei increases with handgrip force, suggesting a role in force modulation in humans.
Contribution
The study provides direct evidence that the reticulospinal tract contributes to force modulation in humans.
Findings
PRN activation increased with handgrip force for both contralateral and ipsilateral nuclei.
Linear scaling of PRN activity with force was observed bilaterally at the group level.
Ipsilateral PRN showed significant scaling after correcting for multiple comparisons.
Abstract
The neural pathways that contribute to force production in humans are currently poorly understood, as the relative roles of the corticospinal tract and brainstem pathways, such as the reticulospinal tract (RST), vary substantially across species. Using functional magnetic resonance imaging (fMRI), we aimed to measure activation in the pontine reticular nuclei (PRN) during different submaximal handgrip contractions to determine the potential role of the PRN in force modulation. Thirteen neurologically intact participants (age: 28 ± 6 yr) performed unilateral handgrip contractions at 25%, 50%, 75% of maximum voluntary contraction during brain scans. We quantified the magnitude of PRN activation from the contralateral and ipsilateral sides during each of the three contraction intensities. A repeated-measures ANOVA demonstrated a significant main effect of force (P = 0.012, ηp2 = 0.307) for…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
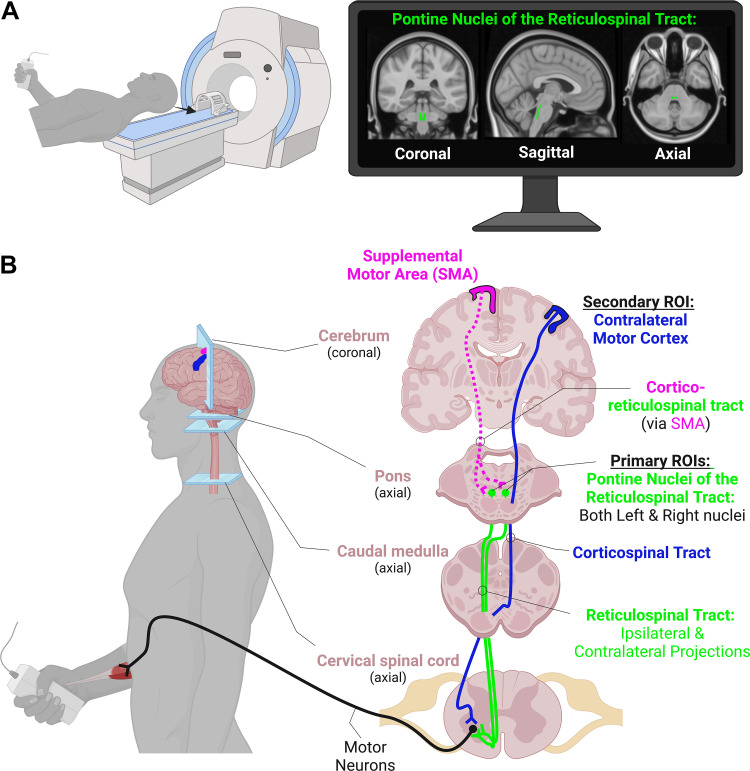 Figure 1
Figure 1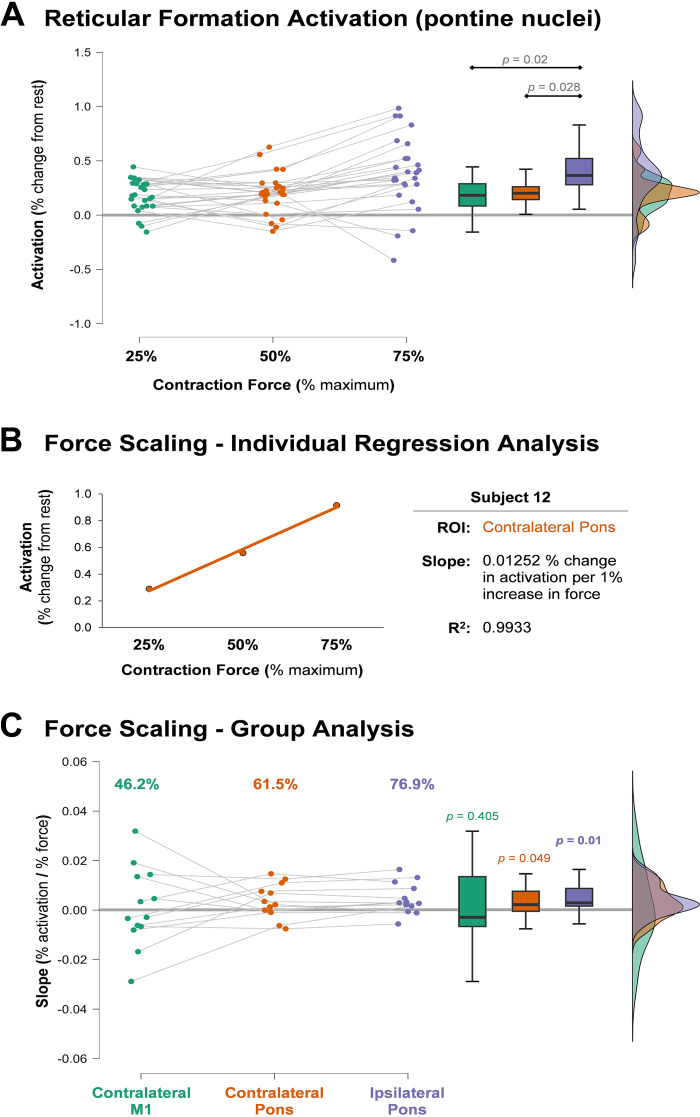 Figure 2
Figure 2- —Canadian Government | Natural Sciences and Engineering Research Council of Canada (NSERC)10.13039/501100000038
- —Canadian Government | Natural Sciences and Engineering Research Council of Canada (NSERC)10.13039/501100000038
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsHistorical and socio-economic studies of Spain and related regions
INTRODUCTION
Within the central nervous system, there are multiple descending pathways that control motor function, and the roles and relative contributions from each pathway may vary substantially across species. In nonmammals, such as reptiles and birds, the main motor pathways descending onto motoneurons originate in the brainstem (1). However, in humans, it is commonly thought that upper limb fine motor control and force modulation is primarily controlled through the corticospinal tract [CST (2, 3)]. An added difficulty in quantifying the role of the CST is how much its size and functionality differs between species. Within humans, the cortico-motoneuronal connections are substantially more developed than in nonhuman primates and other mammals (4), suggesting that the role of CST in humans likely cannot be inferred accurately from animal-model research. Increased development of the CST correlates positively with indices of dexterity (5), giving further evidence for the role of the CST in fine, distal motor production. Due to this specialization of the CST within humans, most research studies investigating the mechanisms of human motor control and muscle force have primarily focused on the CST, with equivocal and heterogeneous findings (6, 7).
Recent evidence, however, now suggests the reticulospinal tract (RST), a brainstem pathway thought to be limited in humans to postural and proximal muscle control, may be more involved in upper limb motor control than previously thought (8–10). In support of this, there has been evidence of distal RST innervation in nonhuman primate models (11) and in humans (8, 9). In addition to its contribution to gross movement control, recent papers have suggested that the RST may also have a role in force modulation (12–14). Some studies have used animal models to study the RST’s role in voluntary force production, but these models require the implantation of electrodes on the RST that is not possible in humans (10, 11, 15–17). For example, Glover and Baker (16) found that the RST activity appears to contribute to contraction force in macaque monkeys. Although, like our study, the evidence was only correlational. Others have examined this control in humans with stroke and spinal cord injury, allowing for models analogous to animal lesion studies (18, 19).
In humans, these studies primarily involve noninvasive stimulation to produce evoked contractions (19–21), whereas animal models have allowed for studies during voluntary movement (11, 15–17, 22). Examining the role of the RST during voluntary force production is important to elucidate how it contributes to the control of human movement. To our knowledge, no study to date has used functional magnetic resonance imaging (fMRI) to corroborate the role of the RST for controlling force in humans despite the benefits of its noninvasive nature. The purpose of this study was to quantify whether the pontine reticular nuclei (PRN), which give rise to the RST, are more active during handgrip contractions at higher compared with lower intensities.
MATERIALS AND METHODS
As a secondary analysis on a data set that has previously been published (23), we aimed to determine the relationship between fMRI blood oxygenation level dependent (BOLD) signal modulation in the pontine reticular formation nuclei and parametric increases in handgrip force during three separate fMRI runs (Fig. 1). Medullary reticular formation nuclei were not included in the analysis due to the medulla not being consistently in the field of view across participants during fMRI scans.
A: handgrip contractions at three different force levels were performed during three separate functional magnetic resonance imaging (fMRI) runs of the cerebrum and brainstem. The monitor in the top right shows our primary region of interest (ROI) in MNI-space, the pontine nuclei of the reticulospinal tract (RST). B: a simplified diagram of the hypothesized pathways involved in voluntary handgrip force production. For comparison, activity levels in the contralateral motor cortex (M1) were also assessed. Not shown, but also potential contributors, are the contralateral supplemental motor area (SMA) and its bilateral projections to the RST on each side, potential connections to the RST from the motor and premotor cortices, and crossed and uncrossed pathways from the ipsilateral M1. Figure was created in biorender.com.
Participants
Thirteen neurologically intact individuals volunteered to participate in this study (11 right-handed, 2 left-handed; sex: 7 females, 6 males; age: 28 ± 6 yr; height: 170.9 ± 9.8 cm; weight: 75.1 ± 16.7 kg). Participants were eligible if they were 18 yr or older and were MRI safe. The presence of intracranial metal clips, cardiac pacemaker, or any factor that would prevent a safe MRI scan disqualified individuals to participate.
Ethical Approval
All participants provided written informed consent before participation. The study adhered to the Declaration of Helsinki and received approval from the University of Saskatchewan Biomedical Research Ethics Board (Ethics No. Bio 01-125).
Experimental Outline
During a single fMRI session, participants engaged in three experimental tasks. These tasks were executed in a random sequence and consisted of submaximal unimanual right isometric handgrip contractions at 25%, 50%, and 75% of their maximum voluntary contraction (MVC). The session began with the acquisition of a structural MRI brain scan, followed by having the participants perform three MVCs. To prevent fatigue, a 1-min rest interval was allotted between each MVC attempt. Based on the MVC results, target force levels were set for the three submaximal experimental tasks. Participants then proceeded to complete the three experimental tasks during separate fMRI runs in random order.
Behavioral Motor Task
Each of the submaximal handgrip conditions were acquired in separate block design fMRI runs. For each run, participants performed 5 sets × 5 repetitions of either a 25%, 50%, or 75% MVC handgrip visuomotor target matching task with an MRI-compatible hand clench dynamometer (Biopac Systems Inc. Aero Camino Goleta, CA). Task blocks involved 1,650 ms [i.e., corresponding to the repetition time (TR) of the T2* fMRI scan] contractions alternating with 1,650 ms of rest (16,500 ms total task block). Each task block was separated by blocks of complete rest (16,500 ms total rest block). During each fMRI run, the participants wore MRI compatible goggles and viewed a computer screen projection, where they were able to view a custom-built LabView (v.8.6) interface. Participants saw clear target lines and go/no-go on screen flashing lights and were cued when to contract or relax. The LabView interface was triggered by the MRI through a fiber optic cable that sent a signal at the beginning of each TR. This signal went through a Siemens fMRI trigger converter and passed to the LabView interface via a BNC connection. This allowed for precise continuous synchronization between the MRI and the experimental paradigm computer throughout the scan (24). Target lines were presented relative to the individual’s peak MVC and force feedback was presented as a vertical force bar that was responsive to each participant’s handgrip contraction (i.e., harder contraction resulted in the bar rising vertically). Participants were visually instructed when to contract and when to relax with a green light that would turn on when the participant was to contract. A second red light was programmed to turn on during rest blocks to indicate a sustained rest. Participants were instructed to relax their nonactive arm throughout each of the fMRI runs.
Handgrip Motor Task Processing
The handgrip force data were processed in Matlab using custom scripts (25). Data were processed with a fourth-order 100 Hz low-pass Butterworth filter and full-wave rectified. The mean from within the full width half maximum of each contraction was then normalized to the mean kg-force of the highest MVC and expressed as a percentage of MVC. The mean of each task block (five contractions) was calculated and used in subsequent analyses.
fMRI Parameters
All scans were acquired in a Siemens 3 T MAGNETOM Skyra MRI (Siemens Healthcare, Erlangen, Germany). At the start of the session, a whole brain T1-weighted (T1w) structural scan was acquired using a high-resolution magnetization prepared rapid acquisition gradient echo (MPRAGE) sequence consisting of 192 T1w echo-planar imaging slices (1-mm slice thickness with no gap), with an in-plane resolution of 1 × 1 mm [field of view = 256 × 256; TR = 1,900 ms; echo time (TE) = 2.08 ms]. Each of the experimental fMRI runs was acquired using T2*-weighted single-shot gradient-echo planar imaging with an interleaved ascending sequence, consisting of 105 volumes (TR = 1,650 ms; TE = 30 ms) of 25 axial slices of 4 mm thickness (1-mm gap) with an in-plane resolution of 2.7 mm × 2.7 mm (field of view = 250) using a flip angle of 90°. The top two coil sets (16 channels) of a 20-channel Siemens head coil (Siemens Healthcare) were used. Runs consisted of a 10-volume alternating block design beginning with five volumes for stabilization (task, rest; 105 volumes total).
fMRI Preprocessing
Functional MRI data processing was carried out using FMRI Expert Analysis Tool (FEAT) Version 6.00, as part of FSL [FMRIB’s Software Library (26)]. Boundary-based registration was used to register the functional image to the high-resolution T1w image. Registration of the functional images to the T1w image was carried out using FMRIB’s Linear Image Registration Tool [FLIRT (27, 28)], and then nonlinearly (rigid + affine + deformable syn) registered to the MNI152_1mm brain template using antsRegistrationSyN and antsApplyTransforms from Advanced Normalization Tools [ANTs (29, 30)]. The following preprocessing was applied: motion correction using Motion Correction FMRIB’s Linear Image Registration Tool [MCFLIRT (28)]; non-brain removal using FNIRT-based brain extraction as implemented in the fsl_anat anatomical processing script (31); spatial smoothing using a Gaussian kernel of FWHM 6 mm; grand-mean intensity normalization of the entire four-dimension (4-D) data set by a single multiplicative factor (32). Next, Independent Component Analysis Automatic Removal of Motion Artifacts (ICA-AROMA) was used remove motion-related noise from each functional run (32). Following ICA-AROMA data clean up, data were high-pass temporal filtered with a 0.01-Hz cutoff. Time-series statistical analyses were carried out using FMRIB’s Improved Linear Model (FILM) with local autocorrelation correction (33). Z (Gaussianised T/F) statistic images were constructed nonparametrically using Gaussian Random Field theory-based maximum height thresholding with a corrected significance threshold of P = 0.05 (34).
Region of Interest Signal Change
FSL’s featquery tool was used to extract the BOLD signal percent change from three regions of interest (ROI) in participants’ native-space. For each ROI, ANTs were used to transform ROIs from MNI-space to native-space prior to signal change extraction. The percent signal change was extracted from the left contralateral and right ipsilateral pontine reticular formation nuclei (left: PnO_PnC_l; right: PnO_PnC_r) from the Brainstem navigator toolkit [https://www.nitrc.org/projects/brainstemnavig (35–41)]. The Navigator Toolkit atlas was developed in vivo by acquiring multiple contrasts (T1w, T2w, T2*, diffusion-weighted imaging) at an ultra-high field strength (7 T) with high spatial resolution (1.1 mm isotropic), and distortion matched diffusion and T2w echo planar images. This approach facilitated the accurate delineation of white and gray matter boundaries on a subject-by-subject level after data were registered to a 1-mm stereotaxic space (MNI standard space). Anatomically defined subnuclei were able to be identified in stereotaxic space based on their microstructural diffusion properties when assessed in conjunction with the other image contrasts. This approach avoided misregistration confounds across the anatomical images. Neighboring nuclei and landmarks were used to initially identify structures, and labels were averaged across subjects, thereby making a probabilistic atlas. Finally, this was validated by inter-rater agreement, internal consistency, and nuclei volume (35–41). In addition, the percent signal change was extracted from left contralateral motor cortex (M1) using an ROI mask from the Brainnetome atlas [left M1: A4ul_l (42)]. The extracted values were then used in subsequent statistical analyses.
Statistical Analysis
An omnibus ROI [contralateral pontine reticular nuclei (PRN), ipsilateral PRN] × condition (25%, 50%, 75% MVC) repeated-measures analysis of variance (RM-ANOVA) was run using the magnitude of brain activation from each of the ROI’s (i.e., BOLD signal percent change from rest) to determine whether PRN activation changed with handgrip force modulation. Data were assessed for normality using Shapiro–Wilks in addition to Zskewness and Zkurtosis. ANOVA tests are robust against violations to normality (43, 44) and thus minor violations to normality as determined by Zskewness and Zkurtosis were considered acceptable for running the parametric model. Sphericity was assessed with Mauchley’s test of sphericity and where violated the Greenhouse-Geisser correction was used. Hedges g effect sizes were calculated and reported to adjust for the small sample size. A Bonferroni adjustment was used to correct for multiple comparisons where appropriate post hoc testing was conducted. The α level for significance was set to α = 0.05.
In addition, for each ROI the slope coefficient of brain activation across the three submaximal handgrip force conditions (percent change of BOLD signal activation/percent target force) was calculated on a subject-by-subject basis using linear regression. The slope coefficients were then analyzed using separate one-sample t tests to determine if each ROI scales its magnitude of activation across the varying submaximal force outputs. A positive slope indicates activation in that ROI increases with handgrip force. A negative slope would represent a decrease in activation with an increase in handgrip force and a nonsignificant slope (i.e., not different from zero) would represent that activation did not vary as a function of handgrip force. For this analysis, the α level was set to α = 0.017 (0.05/3 ROIs; contralateral PRN, ipsilateral PRN, contralateral M1) and P values ≤0.017 were considered significant.
RESULTS
Magnitude of BOLD Activation across Contraction Intensities
RM-ANOVA was performed to test the effect of force (25%, 50%, and 75% MVC) and side (ipsilateral and contralateral) on PRN activation. Mauchly’s test of sphericity was violated for Force (P = 0.023) and Force × Side (P = 0.006). There was a significant main effect for force [F(1.337,16.039) = 5.306, P = 0.012, = 0.307], but no main effect for side [F(1,12) = 0.413, P = 0.533, = 0.033]. The interaction effect (Force × Side) was also not significant [F(1.243,14.921) = 0.957, P = 0.364, = 0.074]. A Bonferroni post hoc analysis showed that activation of pontine nuclei at 75% MVC was significantly greater than at 25% MVC [mean difference (MD) = −0.2, SE = 0.068, P = 0.02] as well as at 50% MVC (MD = −0.179, SE = 0.068, P = 0.042). The difference in activation between 25% and 50% MVC was not significant (MD = −0.021, SE = 0.068, P = 1.000; Fig. 2A).
A: the activation in the pontine nuclei of the reticular formation during contractions of varying intensities. The contralateral and ipsilateral nuclei are collapsed together because there was neither a side × force intensity interaction nor a main effect for side as a factor. Pontine activation did show a main effect for force intensity, with post hoc pairwise comparisons shown above the box plots. B: the relationship between % activation and % force from a single region of interest (ROI) from a participant with a linear fit to clearly demonstrate the slope analysis. The relationship was fit with a linear model, and the slope interpreted as the degree to which activation for that region changes with increases in force. C: the slope coefficients from each subject for each ROI. The values above the scatter plot demonstrate the percent of subjects that exhibited a positive slope (i.e., positive scaling in activation with increasing force) for each ROI. The P values above the box plots are from one-sample t tests, showing that the mean of the slopes was significantly different than zero in the pons before correcting for multiple comparisons, but not in motor cortex (M1). Only the ipsilateral pontine reticular nuclei (PRN) significantly scaled after statistical correction (corrected α = 0.017). The thick gray horizontal lines in A and C extend the zero point for easy comparison.
ROI BOLD Activation Scaling with Force
Linear regression analyses were conducted on each ROI for each subject to determine the slope coefficient, which quantifies the rate of increase in activation with increasing contraction intensity (Fig. 2B). Next, using separate one sample t tests of the individual slopes, we showed at the group-level pontine reticular activation was significantly different from zero on both the ipsilateral [t(12) = 2.672, P = 0.01, g = 0.712] and contralateral [t(12) = 1.792, P = 0.049, g = 0.478] sides before correction; however, after a manual Bonferroni correction for multiple comparisons, only the ipsilateral side remained significant. The contralateral M1 was not statistically different from zero [t(12) = 0.245, P = 0.405, g = 0.065], indicating it did not reliably scale with force (Fig. 2C).
DISCUSSION
The present study examined the relationship between unimanual handgrip force production and BOLD signal activation of the contralateral and ipsilateral PRN of the RST, as well as the contralateral M1 for comparison. Our primary finding is that activation within the PRN consistently scaled linearly with increased force production. The traditional understanding is that the RST primarily innervates axial and proximal muscle groups in humans (45). However, over the past 15 yr a small body of literature has begun to challenge this belief (8, 9, 11, 46) with research suggesting the RST may reach distal musculature of the arms and hands and modulate force (12–14). With this task, we have shown, for the first time, PRN BOLD signal activation significantly scales with handgrip force, where a positive relationship between the magnitude of activation and force output are observed. Specifically, we observe significant scaling in the ipsilateral PRN (with a large effect size) after correcting for multiple comparisons. There was also scaling with a moderate effect size in the contralateral PRN. In addition, the RM-ANOVA showed that side (ipsilateral vs. contralateral) had no effect on activation across the force conditions. These findings, paired with the lack of scaling in contralateral M1 provide some support for the notion of the PRN and possibly the RST pathway contribute to high force outputs, rather than the traditional belief that human distal motor control is exclusively governed by contralateral cortical regions (47). Firing rates within populations of M1 cells are highly variable with force modulation (6, 16, 47), which is supported by a lack of M1 BOLD signal scaling with varying handgrip contraction forces (23). Glover and Baker (16) also found that cells in the reticular formation almost exclusively had a positive correlation between firing rate and force. Our data in humans support their conclusion, as we found no robust support for the role of M1 in force modulation in our previous publication (23), and add to it with evidence that force modulation may be driven by the PRN, and possibly the RST.
During isometric contractions, the recruitment of motor units tend to occur in an orderly fashion from low-threshold to higher-threshold (48). As force demand increases, larger and higher-threshold motor units are recruited (49–52), and the firing rates of smaller, lower-threshold motor units that were already active increase. These recruitment properties may also affect neuromuscular plasticity, as the repeated activation of the larger motor units with high-intensity resistance training often leads to greater gains in strength compared with lower-intensity training (53). In addition to the impact of high-intensity contractions on motoneuron recruitment, and strength, a recent review by Hortobágyi et al. (54) reported high-intensity resistance training also produces greater motor-related neuroplasticity in the neurologically intact young adult brain. However, mechanistic studies into the specific neural pathways and adaptations to resistance training that affords this motoneuronal recruitment remains incompletely understood. This incomplete mechanistic understanding is due in part to research focusing on the CST, which has resulted in equivocal insight. Here, we add to the growing body of literature that pinpoints the PRN, and as a potentially relevant nuclei for force control with a handgrip task, which, until recently (12–14), was thought to be exclusively controlled by the CST (55). Specifically, we show evidence for intensity-dependent activation of the PRN that is correlated with force output, which suggests this is a task relevant pathway for handgrip force. Furthermore, based on the principles of use-dependent plasticity, the repeated preferential activation of the PRN with high-intensity contractions over the course of a resistance training intervention may lead to the strengthening of the RST pathway’s integrity in a manner that facilitates strength adaptations. Recent findings have led to proposals that the RST may have a role in modulating motor unit discharge rates, which would in turn alter force and rate of force production (14). Škarabot et al. (14) proposed hypothesis about the role of RST in modulating force that is supported by the findings of this study.
Evaluating the RST in applied settings presents its own set of challenges, such as the need for external stimuli paired with auditory perturbations (e.g., transcranial magnetic stimulation or cervical medullary electrical stimulation paired with loud acoustic startle sounds ≥ 110 dB), in addition to a high variance in responses. Here, using fMRI, we show a noninvasive evaluation of the PRN in a controlled environment during voluntary motor behaviors. We argue this is an important contribution to the field investigating the role of the RST, as these data provide support to previous findings without the need for external perturbations. Furthermore, when paired with previous research (12, 13, 15), our findings indicate that the RST may be a functionally relevant pathway contributing to muscle force and, by extension, possibly chronic strength training adaptations.
Limitations and Future Directions
Investigating patterns of brain activation during high-intensity contractions with fMRI is not a trivial task, as head motion commonly occurs during effortful movements, and can result in the loss of data due to motion-related contamination. With that, the investigation of fMRI-related BOLD signal activation during different types of motor tasks may not be possible, and thus these experiments are limited to simpler isolated hand movement patterns that are less likely to produce high amounts of motion.
Though we present evidence for the role of the PRN in gross distal hand movement, future research may consider testing PRN activation and RST excitability during finer distal movements. By focusing on dexterous movements such as flexion, extension, abduction, adduction, and circumduction, RST activation bias toward certain types of movement may be determined. Future research into this area should investigate the contributions of the RST to different tasks and their adaptations to training interventions with varying manipulations. A deeper understanding of how the RST contributes to motor function could lead to better rehabilitative modalities for those with dysfunction of the CST, such as individuals with stroke (8, 12). In addition, we focused only on the pontine nuclei of the RST. The medullary nuclei had to be excluded because the medulla was cut off from the scan for many of the participants in favor of capturing complete cortex. Future work should examine whether the medullary nuclei also play a potential role in force modulation. Finally, future research would benefit from replication of this experiment but with a larger sample size to build confidence in data interpretation and robustness of findings. Two final limitations or considerations of this work, that is inherent with all fMRI research, that should be noted when interpreting the data in relation to single-cell invasive measures of neurophysiology are 1) fMRI is not directly measuring neural activity, but it is modeling the hemodynamic response which is related to the local field potentials at the cellular level (56), and 2) the spatial resolution of fMRI is not capable of measuring single-cell neural activity. Within a typical fMRI voxel, there can be a few million individual neurons, and even more synapses that make up the measured signal (57). These two considerations make it difficult to directly compare these population-level hemodynamic measures with findings from single-cell recordings in invasive studies.
Conclusions
In a small cohort of neurologically intact young adults, we demonstrate that activation of the PRN of the RST scales in magnitude with handgrip force. These data provide preliminary insight into the potential role of the PRN and possibly the RST during distal muscular force modulation, a concept most often considered under exclusive control by the CST in humans. These data add to the growing body of literature suggesting a potentially important role of the RST for neuromuscular strength adaptations in humans and lays the foundation for which future research can further investigate RST function to disentangle the neural underpinnings of force modulation and the neural adaptations governing increases in strength.
DATA AVAILABILITY
Data are available upon reasonable request.
GRANTS
This study was funded by a Natural Sciences and Engineering Research Council of Canada (NSERC) Discovery Grant 2016-0529 (to Jonathan P. Farthing), in addition to an NSERC Discovery Grant: 183968-2013-25 (to Ron Borowsky).
DISCLOSURES
No conflicts of interest, financial or otherwise, are declared by the authors.
AUTHOR CONTRIBUTIONS
L.A.G., R.B., J.P.F., and J.W.A. conceived and designed research; L.A.G., C.E., R.B., J.P.F., and J.W.A. performed experiments; T.L.D., C.E., and J.W.A. analyzed data; J.M.D., R.B., J.P.F., and J.W.A. interpreted results of experiments; J.M.D. and J.W.A. prepared figures; T.L.D. and J.W.A. drafted manuscript; J.M.D., R.J.M., R.B., J.P.F., and J.W.A. edited and revised manuscript; T.L.D., L.A.G., J.M.D., R.J.M., C.E., R.B., J.P.F., and J.W.A. approved final version of manuscript.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Kalman M. Evolution of the brain: at the reptile-bird transition. In: Encyclopedia of Neuroscience, edited by Binder MD, Hirokawa N, Windhorst U. Berlin, Heidelberg: Springer Berlin Heidelberg, 2009, p. 1305–1312.
- 2Evarts EV. Activity of pyramidal tract neurons during postural fixation. J Neurophysiol 32: 375–385, 1969. doi:10.1152/JN.1969.32.3.375. 4977837 · doi ↗ · pubmed ↗
- 3Evarts EV. Relation of pyramidal tract activity to force exerted during voluntary movement. J Neurophysiol 31: 14–27, 1968. doi:10.1152/JN.1968.31.1.14. 4966614 · doi ↗ · pubmed ↗
- 4Lemon RN. Descending pathways in motor control. Annu Rev Neurosci 31: 195–218, 2008. doi:10.1146/annurev.neuro.31.060407.125547. 18558853 · doi ↗ · pubmed ↗
- 5Heffner R, Masterton B. Variation in form of the pyramidal tract and its relationship to digital dexterity. Brain Behav Evol 12: 161–200, 1975. doi:10.1159/000124401. 1212616 · doi ↗ · pubmed ↗
- 6Maier MA, Bennett KMB, Hepp-Reymond MC, Lemon RN. Contribution of the monkey corticomotoneuronal system to the control of force in precision grip. J Neurophysiol 69: 772–785, 1993. doi:10.1152/jn.1993.69.3.772. 8463818 · doi ↗ · pubmed ↗
- 7Kidgell DJ, Bonanno DR, Frazer AK, Howatson G, Pearce AJ. Corticospinal responses following strength training: a systematic review and meta-analysis. Eur J Neurosci 46: 2648–2661, 2017. doi:10.1111/EJN.13710. 28921683 · doi ↗ · pubmed ↗
- 8Baker SN, Perez MA. Reticulospinal contributions to gross hand function after human spinal cord injury. J Neurosci 37: 9778–9784, 2017. doi:10.1523/JNEUROSCI.3368-16.2017. 28871033 PMC 5628413 · doi ↗ · pubmed ↗