Progression of herpesvirus infection is inhibited by calcium reporter
Kari Kunnas, Maija Vihinen-Ranta, Simon Leclerc

TL;DR
This study shows that a calcium reporter can inhibit the progression of herpesvirus infection by affecting mitochondrial calcium levels.
Contribution
The study introduces a novel approach using a calcium reporter to interfere with herpesvirus infection progression.
Findings
HSV-1 infection increases mitochondrial calcium and transiently boosts ROS levels.
Cells with a calcium reporter showed reduced viral replication compartments.
The calcium reporter's binding interferes with the infection process.
Abstract
During infection, Herpes simplex virus type 1 (HSV-1) alters the mitochondrial structure and function of the host cell. Live-cell imaging with fluorescent reporters revealed increased mitochondrial calcium and a transient ROS enrichment after HSV-1 infection. Notably, cells co-transfected with a calcium reporter displayed smaller viral replication compartments, while those with a ROS reporter exhibited average growth of viral replication compartments. Our findings suggest that the virus-induced increase in mitochondrial calcium, followed by an increased amount of bound calcium reporter, interferes with the progression of the infection.
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
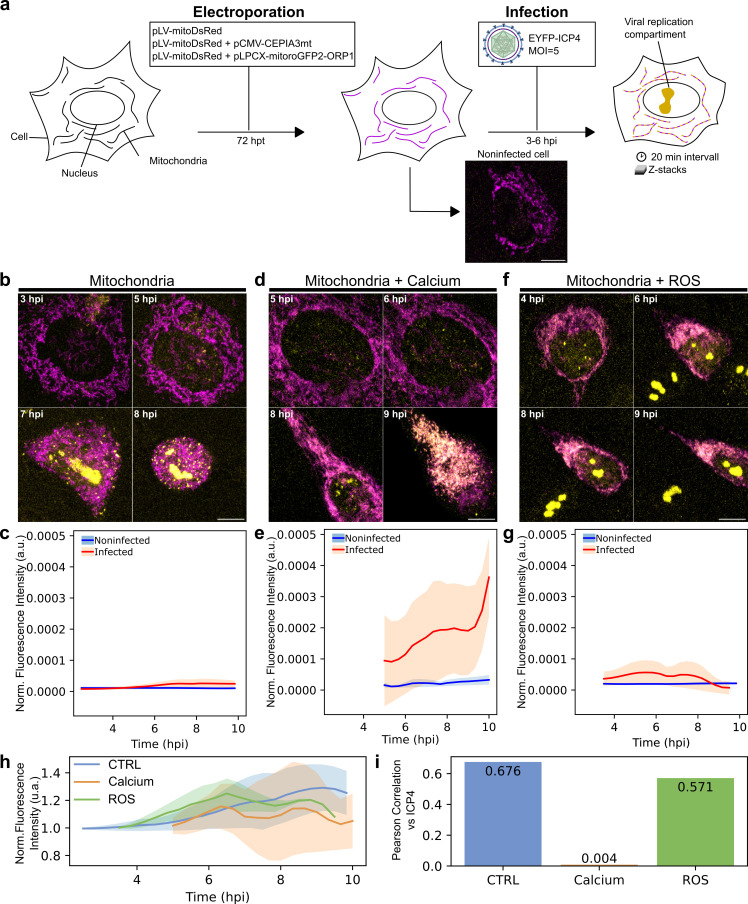 Figure 1
Figure 1|
|
|
|
|
58219 |
pCMV CEPIA3mt |
|
|
44386 |
pLV-mitoDsRed |
|
|
64977 |
pLPCX mito Grx1-roGFP2 |
|
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsHerpesvirus Infections and Treatments · Cytomegalovirus and herpesvirus research · RNA regulation and disease
Description
Mitochondria, the powerhouses of the cell, are essential for ATP production through oxidative phosphorylation. These complex organelles also play a crucial role in the innate immune response (Sorouri, Chang, and Hancks 2022) , particularly through signalling pathways involving calcium (Ca²⁺) (Giorgi, Marchi, and Pinton 2018) and reactive oxygen species (ROS) (Sena and Chandel 2012) . Mitochondrial Ca²⁺ regulates diverse cellular processes. Elevated mitochondrial Ca²⁺ can enhance the Krebs cycle, increasing ATP production to meet cellular energy demands (Traaseth et al. 2004) . Conversely, excessive Ca²⁺ can cause the opening of the mitochondrial permeability transition pore, triggering cell death pathways (Endlicher et al. 2023) . ROS, generated as by-products of the mitochondrial electron transport chain, are signalling molecules affecting gene expression and cell survival (Miller et al. 2019) . However, elevated levels of ROS disrupt cellular homeostasis, causing oxidative stress and damage to biomolecules (Kowalczyk et al. 2021) .
The balance between Ca²⁺ and ROS within mitochondria is critical in modulating the cellular response to viral infections. However, viral counteractions can manipulate mitochondrial functions, cellular metabolism, and immune responses to facilitate viral replication and spread (Foo et al.,2022). Herpes simplex virus type 1 (HSV-1) is a common human pathogen that can cause various infections, from cold sores to more serious neurological diseases. During infection, HSV-1 reorganizes mitochondrial morphology (Leclerc et al. 2024) and function (Vastag et al. 2011) . Our live-cell studies demonstrate that HSV-1 infection induces a substantial increase in mitochondrial Ca²⁺ and a moderate transient increase of ROS.
To investigate the impact of HSV-1 infection on mitochondrial function in mouse embryonic fibroblast (MEF) cells, we employed live-cell imaging of transiently transfected plasmids expressing fluorescent reporters of mitochondria, Ca²⁺, and ROS. The mitochondria were visualized by fluorescent protein (mitoDsRed) targeted to the mitochondria. Ca²⁺ was detected by a genetically encoded indicator (CMV-CEPIA3mt) and ROS by its indicator (LPCX-mito roGFP-ORP1). Cells were infected 72 hours post-transfection with HSV-1 at a multiplicity of infection (MOI) of 5. Viral replication and localization of a viral replication compartment were verified by the expression of a viral EYFP-ICP4 fusion protein (Fig1. a). The emission spectra of the mitochondrial indicators and EYFP-ICP4 are similar (yellow). However, the nuclear localization of EYFP-ICP4 and the cytoplasmic presence of indicators allowed their separation during analysis.
As previously described (Kobiler et al. 2011; Simpson-Holley et al. 2005; Aho et al. 2017) , our studies confirm the emergence of small, dispersed viral replication compartments at 5 hours post-infection (hpi), followed by their fusion into larger compartments at 7-8 hpi. Later, at 12 hpi, extensive cytopathic effects are observed, including rounded cells with disrupted nuclear membranes, chromatin marginalization, and cell detachment from the coverslip (Fig1. b). This was potentially due to the combined effects of viral infection and extended observation. The viral reporter EYFP-ICP4 expression and the condensation of the mitochondria towards the nuclear envelope followed the progression of infection. In infected cells transfected with mitoDsRed, the viral replication compartment growth and fusion and the change in mitochondria and cell morphology are visible (Fig1. b). The yellow fluorescence of mitochondria remained near zero without mitochondrial function reporter (Fig1. c).
In cells co-transfected with the mitoDsRed and Ca²⁺ reporter, the viral replication compartment started to form at 4 hpi but failed to expand later in infection (Fig1. d). An increased amount of Ca² accompanied the progression of infection with a significant increase between 5 and 10 hpi (Fig1. e). In cells co-transfected with the mitoDsRed and ROS reporter, the viral replication compartment formed was similar to control cells (Fig1. f). Notably, cells started to detach from the coverslip after 6 hpi and the amount of ROS decreased as the infection proceeded (Fig1. g). This suggests that dual transfection of infected cells with a mitoDsRed and ROS reporter led to disturbance of cellular morphology and cell death faster than only dsRed transfected cells. The analysis of the mitochondrial reporters during the progression of infection showed that the intensity of signals of markers fluctuated when the intensity of EYFP-ICP4 and viral replication compartment increased ( Fig. 1 h). In mitoDsRed-labeled control cells, a positive correlation between mitoDsRed and EYFP-ICP4 was observed, reflecting the higher cytoplasmic background resulting from EYFP-ICP4 produced during successful infection. Interestingly, the calcium reporter showed no correlation with EYFP-ICP4, verifying the inhibition of viral replication in these cells. Finally, the ROS reporter displayed a moderate positive correlation with viral protein, suggesting some ROS production even with reduced viral replication ( Fig. 1 i).
Altogether, our results show that the formation of an enlarged viral replication compartment was inhibited in cells transfected with Ca²⁺ reporter. Our previous studies demonstrated an infection-induced increase in the overall cellular Ca²⁺ levels (Leclerc et al. 2024) . Our results suggest that the increased presence of calcium leads to elevated levels of reporters. While the calcium-binding affinity of the calcium reporter might be weak, its interaction with Ca²⁺, specifically within the mitochondria could interfere with the progression of infection. This hypothesis needs further investigation, such as using mitochondrial Ca²⁺ chelators like Ruthenium Red (Marmolejo-Garza et al. 2023) , which could help elucidate whether manipulating the available mitochondrial Ca²⁺ pool influences HSV-1 replication dynamics. It is also crucial to consider other factors, such as transfection methods or plasmid constructs, that might impact cell viability. Therefore, using different calcium reporter variants (Kanemaru et al. 2020) could help confirm our findings.
Methods
Cells and viruses
Mouse embryonic fibroblast cells (MEF, ATCC CRL-2991) were grown in Dulbecco’s modified Eagle medium (DMEM) supplemented with 10% foetal bovine serum, L-glutamine, and penicillin-streptomycin (Gibco-Invitrogen, Carlsbad, CA) at 37°C in the presence of 5% CO2.
Electroporation
MEF cells were trypsinized, counted, and 5 x 10 ^5^ cells were resuspended in 110 µl of resuspension buffer (Invitrogen). 4 μg of each plasmid was added to the cell and electroporated (Neon transfection system, Invitrogen) using a single pulse of 30 ms at 1350 volts. After electroporation, cells were split into two wells at a 0.7 to 0.3 ratio. The cells were then grown on a square coverslip in 6 well plates for 72 hours.
Viral infection
On the day of infection, cells were infected with the EYFP-ICP4 (vEYFP-ICP4) strain (Everett et al. 2004) at a multiplicity of infection (MOI) of 5. After one hour, the culture medium was replaced with a phenol-free imaging medium (same formulation, Gibco-Invitrogen, Carlsbad, CA) and incubated for 3-5 hours at 37°C with 5% CO2 before imaging.
Microscopy and image acquisition
Cells were imaged on a Leica TCS SP8X Falcon confocal microscope (Leica Microsystems, Mannheim, Germany) with a water immersion objective (HC PL APO CS2, NA: 1.2) and a HyD detector. Images were acquired at 3x zoom with a pixel size of 190 nm. Z-stacks consisted of 16 planes spaced 500 nm apart. To minimize photobleaching and crosstalk, YFP/roGFP/CEPIA (excitation: 498 nm, emission: 508-545 nm) and DsRed (excitation: 560 nm, emission: 580-650 nm) were acquired sequentially. Autofocus (Leica Microsystems, Mannheim, Germany) compensated for Z-drift during the acquisition. Each Z-stack acquisition took 30-40 seconds. 20 cells were imaged at 20-minute intervals over 12 hours.
Image processing and analysis
Following image acquisition, individual cells were isolated from the acquired Z-stacks. This involved cropping the images to eliminate background and signals from neighbouring cells. To segment mitochondria within each isolated cell, we employed the dsRed-Mito7 channel and the Otsu thresholding method. Next, we quantified two parameters for each cell across the entire timelapse: mitochondrial volume and raw green fluorescence intensity. The mitochondrial volume provides information about the overall size of the mitochondria network, while the raw green fluorescence intensity, measured from the mitochondrial function reporters (CEPIA or roGFP-ORP1), reflects the reporter activity within the mitochondria. To account for potential variations in mitochondrial size, the green fluorescence intensity was normalized by dividing it by the corresponding mitochondrial volume for each cell. We further analysed the normalized green fluorescence by calculating average traces for each condition, followed by smoothing the average trace using a Savitzky-Golay filter (Poly-order 3). A similar procedure was applied to obtain average and smoothed traces for the standard deviation.
Sample size:
Control (pLV-mitoDsRed only) - Noninfected cells: 11 - Infected cells: 9
Calcium (pLV-mitoDsRed and pCMV CEPIA3mt) - Noninfected cells: 10 - Infected cells: 6
ROS (pLV-mitoDsRed and pLPCX mito roGFP2-ORP1) - Noninfected cells: 6 - Infected cells: 5
Reagents
**: **
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Aho Vesa Myllys Markko Ruokolainen Visa Hakanen Satu MäntyläElina Virtanen Jori Hukkanen Veijo Kühn Thomas Timonen Jussi Mattila Keijo Larabell Carolyn A. Vihinen-Ranta Maija 2017616 Chromatin organization regulates viral egress dynamics Scientific Reports 712045-232210.1038/s 41598-017-03630-y PMC 547383428623258 · doi ↗ · pubmed ↗
- 2Endlicher René Drahota Zdeněk ŠtefkováKateřina ČervinkováZuzana Kučera Otto 2023427 The Mitochondrial Permeability Transition Pore—Current Knowledge of Its Structure, Function, and Regulation, and Optimized Methods for Evaluating Its Functional State Cells 1292073-44091273127310.3390/cells 1209127337174672 PMC 10177258 · doi ↗ · pubmed ↗
- 3Everett Roger D. Sourvinos George Leiper Claire Clements J. Barklie Orr Anne 2004215 Formation of Nuclear Foci of the Herpes Simplex Virus Type 1 Regulatory Protein ICP 4 at Early Times of Infection: Localization, Dynamics, Recruitment of ICP 27, and Evidence for the De Novo Induction of ND 10-Like Complexes Journal of Virology 7840022-538X 1903191710.1128/jvi.78.4.1903-1917.200414747555 PMC 369473 · doi ↗ · pubmed ↗
- 4Giorgi Carlotta Marchi Saverio Pinton Paolo 2018824 The machineries, regulation and cellular functions of mitochondrial calcium Nature Reviews Molecular Cell Biology 19111471-007271373010.1038/s 41580-018-0052-830143745 · doi ↗ · pubmed ↗
- 5Gutscher Marcus Pauleau Anne-Laure Marty Laurent Brach Thorsten Wabnitz Guido H Samstag Yvonne Meyer Andreas J Dick Tobias P 2008511 Real-time imaging of the intracellular glutathione redox potential Nature Methods 561548-709155355910.1038/nmeth.121218469822 · doi ↗ · pubmed ↗
- 6Kanemaru Kazunori Suzuki Junji Taiko Isamu Iino Masamitsu 2020218 Red fluorescent CEPIA indicators for visualization of Ca 2+ dynamics in mitochondria Scientific Reports 1012045-232210.1038/s 41598-020-59707-8PMC 702904132071363 · doi ↗ · pubmed ↗
- 7Kitay Brandon M. Mc Cormack Ryan Wang Yunfang Tsoulfas Pantelis Zhai R. Grace 2013111 Mislocalization of neuronal mitochondria reveals regulation of Wallerian degeneration and NMNAT/WLDS-mediated axon protection independent of axonal mitochondria Human Molecular Genetics 2281460-20831601161410.1093/hmg/ddt 00923314018 PMC 3657477 · doi ↗ · pubmed ↗
- 8Kobiler O. Brodersen P. Taylor M. P. Ludmir E. B. Enquist L. W. 20111230 Herpesvirus Replication Compartments Originate with Single Incoming Viral Genomesm Bio 262161-212910.1128/mbio.00278-11PMC 326906522186611 · doi ↗ · pubmed ↗