Antibiotic Resistance Profiles of Escherichia coli and Salmonella spp. Isolated From Dairy Farms and Surroundings in a Rural Area of Western Anatolia, Turkey
Savaş Aslan, Cengiz Demir, Elçin L Kurtoğlu, Mustafa Altındiş

TL;DR
This study examines antibiotic resistance in E. coli and Salmonella from Turkish dairy farms and surrounding areas, highlighting the presence of resistance genes.
Contribution
The study provides new antibiotic resistance profiles and gene detection data for E. coli and Salmonella in rural dairy environments in Turkey.
Findings
E. coli showed highest sensitivity to meropenem and lowest to cefazolin and cefuroxime.
Salmonella isolates were resistant to tetracycline and penicillin/ampicillin.
Only 7.7% of E. coli strains were ESBL-positive, and resistance genes were absent in Salmonella.
Abstract
Background Antibiotic resistance is a significant public health issue worldwide. Antibiotic-resistant zoonotic bacteria such as Escherichia coli (E. coli), Campylobacter, Salmonella, Listeria, Coxiella, and Mycobacterium can be particularly isolated from biofertilizers. Epidemiological studies have shown that cases of foodborne infections and intoxications are significantly related to animal-derived foods. The presence of these species in aquatic environments indicates areas or organisms contaminated with animal or human feces. Especially, the presence of E. coli in aquatic environments has become a serious problem worldwide. Pathogenic strains of E. coli cause waterborne and foodborne diseases. Materials and methods This study included a total of 290 samples collected from five different dairy farms between April and September 2023 which comprised 20 samples of cow manure, 20…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
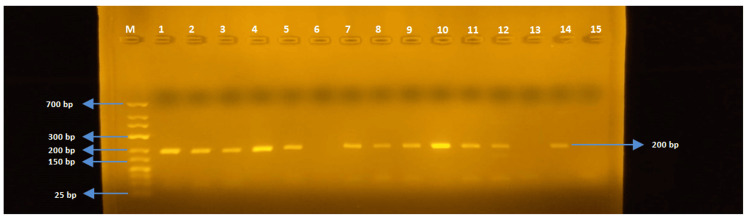 Figure 1
Figure 1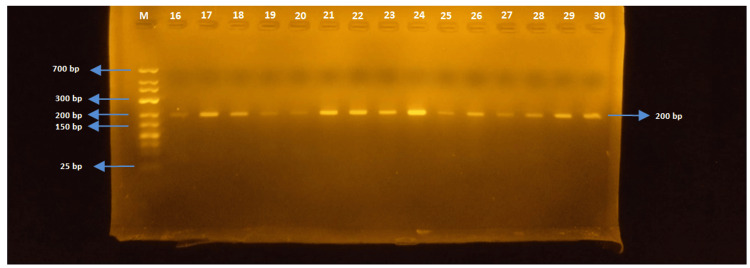 Figure 2
Figure 2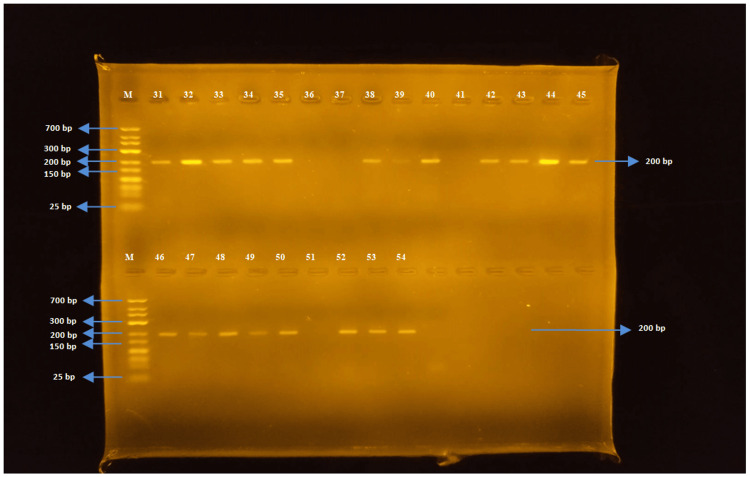 Figure 3
Figure 3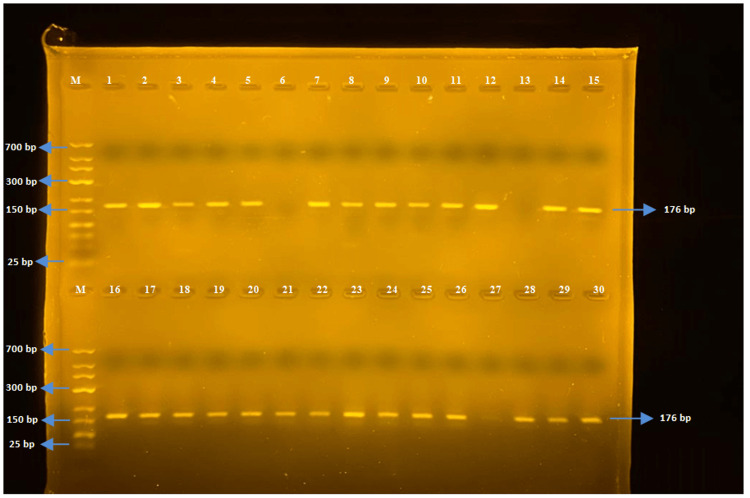 Figure 4
Figure 4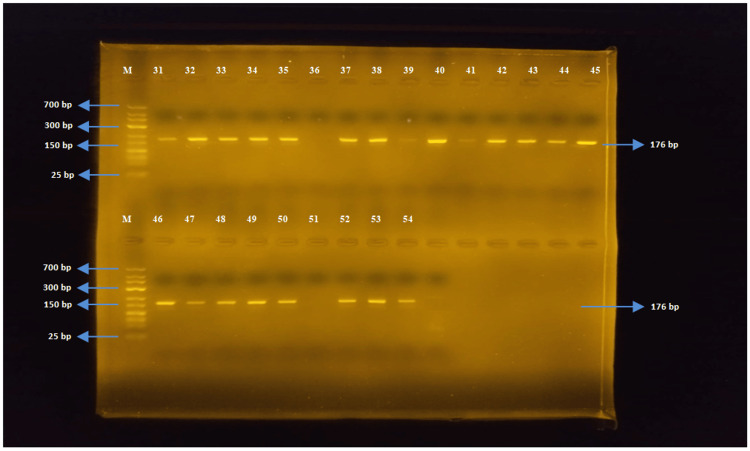 Figure 5
Figure 5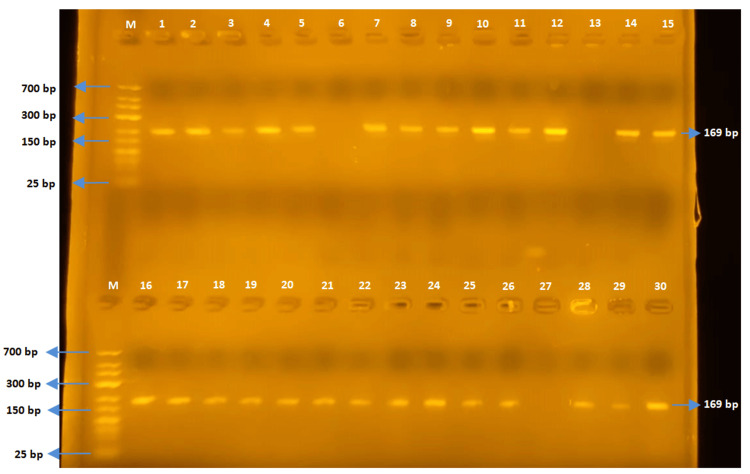 Figure 6
Figure 6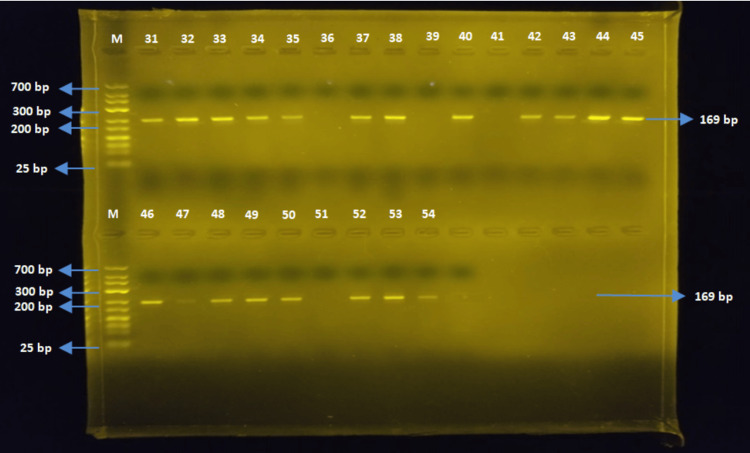 Figure 7
Figure 7| Genes | Forward (F)/reverse (R) | Primers | Base pair (bp) | References |
| ereA | F-Primary | 5’- ACCGCACGTTGATATGTTGA -3’ | 182 bp | Volokhov et al., 2003 [ |
| R- Primary | 5’- CAAATCGCTGTTGACGTGTT -3’ | |||
| tetA | F- Primary | 5’- AATTTCCTGACGGGCTGTTT -3’ | 200 bp | Wu et al., 2024 [ |
| R- Primary | 5’- TGTCCGACAAGTTGCATGAT -3’ | |||
| tetB | F- Primary | 5’- TTCATTAGCGGGTCTTGGTC -3’ | 176 bp | Parson et al., 2024 [ |
| R- Primary | 5’- CCACCACCAGCCAATAAAAT -3’ | |||
| SHV | F- Primary | 5’- CTTTCCCATGATGAGCACCT -3’ | 193 bp | Parson et al., 2024 [ |
| R- Primary | 5’- CGCTGTTATCGCTCATGGTA -3’ | |||
| TEM | F- Primary | 5’- TTTGCCTTCCTGTTTTTGCT -3’ | 169 bp | Alawi et al., 2024 [ |
| R- Primary | 5’- ATAATACCGCGCCACATAGC -3’ | |||
| OXA | F- Primary | 5’- GGATAAAACCCCCAAAGGAA -3’ | 169 bp | Parson et al., 2024 [ |
| R- Primary | 5’- AAGCTACTTTCGAGCCATGC -3’ | |||
| CTX-M | F- Primary | 5’- GGTGATGAACGCTTTCCAAT -3’ | 199 bp | Ellem et al., 2011 [ |
| R- Primary | 5’- TCAATTTGTTCATGGCGGTA -3’ |
| Microorganism | Number | % |
| Acinetobacter baumannii | 3 | 1.6 |
| Acinetobacter baylyi | 2 | 1.1 |
| Acinetobacter calcoaceticus | 4 | 2.2 |
| Acinetobacter calcoaceticus | 1 | 0.5 |
| Acinetobacter courvalinii | 3 | 1.6 |
| Acinetobacter dijkshoorniae | 1 | 0.5 |
| Acinetobacter johnsonii | 1 | 0.5 |
| Acinetobacter pittii | 5 | 2.7 |
| Acinetobacter vivianii | 1 | 0.5 |
| Buttiauxella warmboldiae | 1 | 0.5 |
| Citrobacter braakii | 4 | 2.2 |
| Citrobacter freundii | 4 | 2.2 |
| Comamonas kerstersii | 2 | 1.1 |
| Enterobacter bugandensis | 4 | 2.2 |
| Enterobacter cloacae | 29 | 15.8 |
| Enterobacter ludwigii | 1 | 0.5 |
| Enterococcus faecalis | 2 | 1.1 |
| Escherichia coli | 52 | 28.4 |
|
| 1 | 0.5 |
| Klebsiella aerogenes | 8 | 4.4 |
| Klebsiella oxytoca | 4 | 2.2 |
| Klebsiella pneumoniae | 8 | 4.4 |
| Klebsiella variicola | 4 | 2.2 |
| Kosakonia cowanii | 1 | 0.5 |
| Lactobacillus curvatus | 1 | 0.5 |
| Lactobacillus rhamnosus | 1 | 0.5 |
| Lactococcus lactis | 1 | 0.5 |
| Mixta calida | 1 | 0.5 |
| Proteus mirabilis | 2 | 1.1 |
| Proteus vulgaris | 1 | 0.5 |
| Pseudomonas aeruginosa | 3 | 1.6 |
| Pseudomonas cichorii | 1 | 0.5 |
| Pseudomonas koreensis | 2 | 1.1 |
| Pseudomonas monteilii | 1 | 0.5 |
| Pseudomonas otitidis | 1 | 0.5 |
| Pseudomonas plecoglossicida | 1 | 0.5 |
| Pseudomonas protegens | 1 | 0,5 |
| Pseudomonas putida | 6 | 3.3 |
| Pseudomonas putida | 4 | 2.2 |
| Pseudomonas thivervalensis | 1 | 0.5 |
| Raoultella ornithinolytica | 1 | 0.5 |
|
| 2 | 1.1 |
| Serratia liquefaciens | 2 | 1.1 |
| Serratia mercescens | 4 | 2.2 |
| Total | 183 | 100 |
| Antibiotics | Escherichia coli | |
| Sensitive (%) | Resistant (%) | |
| Amikacin | 94.2 | 5.8 |
| Amoxicillin/clavulanic acid | 75 | 25 |
| Ampicillin | 69.2 | 30.8 |
| Cefazolin | 0 | 100 |
| Cefepime | 94.2 | 5.8 |
| Cefoxitin | 94.2 | 5.8 |
| Ceftazidime | 92.3 | 7.7 |
| Ceftriaxone | 92.3 | 7.7 |
| Cefuroxime | 0 | 100 |
| Ciprofloxacin | 80.8 | 19.2 |
| Colistin | 94.2 | 5.8 |
| Ertapenem | 96.2 | 3.8 |
| Gentamicin | 92.3 | 7.7 |
| Meropenem | 100 | 0 |
| Piperacillin/tazobactam | 94.2 | 5.8 |
| Tigecycline | 96.2 | 3.8 |
| Trimethoprim/sulfamethoxazole | 82.7 | 17.3 |
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsPharmaceutical and Antibiotic Environmental Impacts · Salmonella and Campylobacter epidemiology · Listeria monocytogenes in Food Safety
Introduction
Antibiotic resistance continues to be a global public health concern [1]. The impact of antibiotic-resistant bacteria in the etiology of infections continues to rise to concerning levels [2]. It is estimated that approximately 10 million people die each year due to antimicrobial resistance infections [3]. Drug-resistant microorganisms are being reported from all regions of the world, and their adverse effects have significantly increased in recent years [4]. The indiscriminate use of antibiotics and lack of knowledge on this subject are among the most important reasons for the proliferation, selection, and spread of antibiotic-resistant organisms [5]. Many antimicrobial agents are used in animal feed production to control diseases and are often preferred as growth-promoting factors. At the same time, they continuously spread in the human food chain, leading to serious health problems in humans and animals [6]. Cattle on dairy farms are potential sources of contamination with antibiotic-resistant Escherichia coli (E. coli) and Salmonella spp., which are found in cow manure and can contaminate the farm environment and farm products. Moreover, these resistance elements can cause serious human health problems by being transmitted directly to farm workers through contaminated soil, water, and milk [7].
*Escherichia coli *is recognized as a dangerous pathogen in the dairy farm sector worldwide due to the significant economic losses it causes [8]. There are various types of E. coli, most of which are harmless, but a few can cause serious foodborne infections in humans [9]. Farm animals, especially cattle, carry Shiga toxin-producing E. coli (STEC) and enterohemorrhagic *E. coli *(EHEC) asymptomatically. These pathogens are inherently zoonotic and can be transmitted from farms to humans through contaminated milk, meat, water, and direct contact with animals or their environmental equipment [10]. Dairy cows serve as reservoirs for *Salmonella *spp., which can cause human salmonellosis [11]. *Salmonella *spp. can be transmitted from infected cattle and their surroundings through feces. In recent years, the increasing resistance of *Salmonella *serotypes to commonly used antibiotics has significantly increased the cost of treatment in food animal production [12]. Animal manure contains microbial components that make it a potential source of pathogenic microorganisms for both animals and humans. Fresh farm animal manure is produced in many countries and is predominantly used as a biofertilizer in agricultural lands [3]. Various bacterial pathogens, such as E. coli, Campylobacter, Salmonella, Listeria, Coxiella, and Mycobacterium, inherently resistant to antibiotics and zoonotic, have been obtained from manure. These pathogens can enter the food chain in a way that affects consumer health when manure is used as fertilizer for crop, vegetable, and fruit production in agriculture [13].
The emergence of antibiotic-resistant bacteria and their resistance genes has become an increasingly serious problem in existing drugs. There is a lack of sufficient data on the formation of antibiotic-resistant bacteria, particularly in dairy cattle farming systems. The most serious challenge encountered in controlling and treating these pathogens is the increasing drug resistance in *Salmonella *spp. and E. coli strains and the development of multidrug-resistant epidemic types. Therefore, studies on the exact sources of bacterial dissemination and their genetic profiles are very important. In this study, the aim is to characterize antibiotic resistance genes in E. coli and *Salmonella *spp. strains isolated from dairy farms and their surroundings in the Afyonkarahisar province region of Turkey to contribute to the selection of antibiotics for empirical therapy.
This study was supported by the Afyonkarahisar Health Sciences University Scientific Research Projects Coordination Unit (project number: 19.SHMYO.001).
Materials and methods
Sample collection
From each dairy farm, a total of 58 samples were collected, comprising 20 samples of cow manure, 20 samples of milk, three samples of handwashing water from milkers, five soil samples, five water samples, and five vegetable samples (such as spinach, green pepper, and tomato), resulting in a total of 290 samples collected from five different farms. Cow fecal samples were collected immediately after defecation while handwashing samples from dairy farm workers, soil samples, and vegetable samples were collected when sampling vegetables like spinach, green pepper, and tomato.
Sample processing
Two different types of samples, solid (cow feces, soil, and vegetables) and liquid (milk, handwashing water from milkers, and water), were measured in grams and milliliters, respectively. For cow feces and soil samples, 10 g of each sample was taken and homogenized in 90 ml of 0.1% peptone water. Collected vegetable samples were divided into small pieces with a sterile knife and thoroughly mixed to obtain a homogeneous mixture. Twenty-five grams of the mixed diced vegetables were homogenized in 225 ml of 0.1% peptone water. For preparing liquid samples, 10 ml of the sample was mixed with 90 ml of diluent for the initial dilution. Finally, 10-fold serial dilutions were made from all initial dilutions for bacterial enumeration.
Identification of bacterial strains and antibiotic susceptibility testing
Samples were cultured on blood agar, xylose lysine deoxycholate (XLD) agar, and eosin methylene blue (EMB) agar plates. The plates were then incubated at 37°C for 24 hours. Following incubation, colonies identified as Gram-negative were subjected to species-level identification using the matrix-assisted laser desorption/ionization time-of-flight mass spectrometry (MALDI-TOF MS) (Bruker Daltonics, Bruker Microflex LT, Bremen, Germany) and VITEK 2 automated systems (BioMerieux Inc., Durham, NC). Until susceptibility and molecular tests were conducted, strains were suspended in 2 ml sterile Eppendorf tubes containing tryptic soy broth supplemented with 15% glycerol and stored at -80°C. The VITEK 2 automated system was utilized to determine the antibiotic susceptibility of the bacterial strains included in the study. Standard strains of E. coli American Type Culture Collection (ATCC) 25922 and Salmonella typhimurium ATCC 14028 were used for all susceptibility tests.
Bacterial DNA isolation
The DNA extraction was performed using the boiling method. For this purpose, three to five colonies from fresh passages on sheep blood agar were aseptically picked with a sterile loop. These colonies were then suspended in 300 µl of distilled water in labeled sterile Eppendorf tubes, ensuring complete dissolution. Subsequently, the tubes were placed in a water bath filled with water and foam spacers at 95°C for 20 minutes. After incubation, the Eppendorf tubes were centrifuged at 14,000 rpm for five minutes at room temperature. Following centrifugation, 100 µl of the supernatant was used as the template DNA for amplification.
DNA amplification
The DNA samples were amplified using specific primers and A.B.T.™ 2X HS-PCR MasterMix (with BlueDye) (P02-02-01, Turkey) from A.B.T Laboratory Industry, Arnavutköy, Turkey, in the SimpliAmp™ thermal cycler (Thermo Fischer Scientific Inc., Waltham, MA). The thermal cycling conditions were set as follows: initial denaturation at 95°C for five minutes, followed by 40 cycles of denaturation at 95°C for 15 seconds, annealing at 60°C for 30 seconds, extension at 72°C for 30 seconds, and a final extension step at 72°C for five minutes. After amplification, PCR products were visualized using 2% agarose gel electrophoresis. The primer pairs used in resistance gene detection in the obtained isolates ranged between 169 base pair (bp) and 200 bp (Table 1).
Results
In this study, a total of 290 samples collected from five different farms were included. Out of the 290 samples consisting of feces, milk, vegetables, soil, animal irrigation/vegetable irrigation water, and handwashing water, a total of 183 microorganisms were isolated. These 183 microorganisms comprised 44 different species in total. The most frequently isolated microorganisms in our study were E. coli (28.4%), Enterobacter cloacae (15.8%), Klebsiella pneumoniae (4.4%), and Klebsiella aerogenes (4.4%) (Table 2).
Identification revealed that 52 isolates were E. coli, while two were identified as *Salmonella *spp. Antibiotic susceptibility testing was conducted for E. coli strains. Among the 52 E. coli strains, the highest antibiotic susceptibility rate was observed for meropenem, while the lowest susceptibility rate was detected for cefazolin and cefuroxime antibiotics (Table 3). While two of the *Salmonella *spp. (n = 2) isolates were found to be resistant to tetracycline, and one was found to be resistant to penicillin and ampicillin. No resistance to trimethoprim/sulfamethoxazole was detected in either isolate. Extended-spectrum beta-lactamases (ESBLs) were detected in only four (7.7%) *E. coli *strains.
In this study, the molecular analysis of seven different gene regions responsible for antibiotic resistance was conducted. The presence of ereA, tetA, tetB, SHV, TEM, OXA, and CTX-M genes was investigated, and the gel electrophoresis images of the targeted gene regions were evaluated using markers ranging from 25 bp to 700 bp in size.
The tetA gene region (200 bp)
The PCR was performed on *E. coli *and *Salmonella *spp strains with tetA-F and tetA-R primers. While the tetA gene was detected in 47 of 52 *E. coli *strains, the tetA gene region could not be detected in any of the *Salmonella *spp. strains (Figures 1-3).
Samples one to 15 for tetA
Samples 16-30 for tetA
Samples 31-54 for tetA
The tetB gene region (176 bp)
The PCR was performed using tetB-F and tetB-R primers for *E. coli *and Salmonella spp. strains. The tetB gene was detected in 50 out of 52 E. coli strains, while none of the *Salmonella *spp. strains detected the tetB gene region (Figures 4, 5).
Samples one to 30 for tetB
Samples 31-54 for tetB
The TEM gene region (169 bp)
The PCR was conducted using TEM-F and TEM-R primers for E. coli and *Salmonella *spp. strains. The TEM gene was detected in 48 out of 52 E. coli strains, while none of the Salmonella spp. strains detected the TEM gene region (Figures 6, 7).
Samples one to 30 for TEM
Samples 31-54 for TEM
Other gene regions
In the PCR study, OXA, CTX-M, SHV, ereA gene regions could not be detected in any of the 52 E. coli and two Salmonella spp. strains.
Discussion
Antimicrobial resistance poses a serious global public health concern [19]. Inappropriate use of antibiotics by humans, factories, and farms, poor hygiene, and the inability to prevent infections in healthcare facilities are considered significant contributors to the emergence and spread of antibiotic-resistant bacteria [20]. Extended-spectrum beta-lactamases are enzymes that confer resistance to most β-lactam antibiotics, including penicillins, cephalosporins, and the monobactam aztreonam. Infections with ESBL-producing organisms have been associated with poor outcomes [21]. Escherichia coli is a notable example of microorganisms that develop antibiotic resistance, particularly multidrug resistance (MDR), and can cause life-threatening infections by producing ESBLs [22]. Escherichia coli is a facultative member of the flora predominant in the gastrointestinal system of both humans and animals [23]. Prolonged exposure of E. coli and Salmonella spp. to antibiotics contributes to the development of antibiotic resistance. Therefore, antibiotic-resistant bacteria, including E. coli and *Salmonella *spp. in animals, can serve as significant reservoirs for human colonization and infection. Studies have shown that antibiotic-resistant bacteria can spread from the environment to humans through direct or indirect contact (e.g., consuming contaminated food and water) [24]. Therefore, evaluating the prevalence of antibiotic-resistant E. coli and Salmonella spp. from different sources is of critical importance for establishing guidelines in veterinary and human health services. Particularly in developed countries, bacterial resistance monitoring programs are regularly conducted, and the results regarding the resistance status of indicator, pathogenic, and zoonotic bacteria to antibiotics are periodically published [25]. As E. coli is a natural member of human and animal intestinal flora, it can frequently be exposed to antibiotics used for various purposes in animals, thereby easily developing various resistance mechanisms. Another characteristic of E. coli is its ability to transfer resistance genes to pathogenic and zoonotic bacteria, thus being considered an indicator bacterium in monitoring antibiotic resistance [26]. Therefore, it is crucial to examine not only pathogenic bacteria but also normal flora members like E. coli isolates in monitoring antibiotic resistance.
Bacteria within the *Enterobacterales *family have a wide range of hosts, including plants, insects, animals, and humans. Some species of this family are found in the normal intestinal flora of humans and animals [27]. These bacteria are not only pathogens or members of the intestinal flora but are also abundantly found in almost every moist environment, particularly in soil, water, and household settings [28]. In their study, Naderi et al. (2024) [29] analyzed the prevalence of antibiotic resistance genes, phenotypic resistance, and all percentage rates associated with phylogenetic groups among 51 virulence gene-positive E. coli isolates from 36 healthy and 15 diarrheal calves. This study detected 9.8% tetA, 5.9% tetB, 3.9% TEM, and 3.9% bla SHV genes. In their study, Jia et al. (2022) [30] investigated multiple resistance genes found in ultra-broad-spectrum beta-lactamases (ESBL) and AmpC enzymes. In this study, bla CTX-M, bla TEM, and bla SHV were detected at a rate of 29%, 29%, and 9.5%, respectively. In their study of molecular characterization of E. coli strains by Gautam et al. (2019) [31], it was shown that ESBL and AmpC enzymes are commonly found in E. coli of animal origin, the frequency of beta-lactamase antimicrobial drugs has increased, and bla CTX M and bla TEM types are currently more widely distributed genotypes. In a study by Geser et al. (2012) [32] conducted in Switzerland, they found ESBL-producing *Enterobacterales *isolates in 25.3% of animal fecal samples they collected. When the studies conducted in Turkey were examined, the ESBL-producing E. coli strain was not found in adult cattle in the study conducted by Aksoy et al. [33] and Buyuknal et al. [34]. In the survey conducted in Hatay, the prevalence was found to be 8.3% in adult cattle [35]. In a study conducted on fecal samples collected from the Burdur region, the rate of ESBL-producing E. coli was found to be 15.5% [36]. In our study, ESBL was detected in 4 (7.7%) of the isolated E. coli strains. It is observed that the rate of ESBL-producing strains in our study is consistent with other studies conducted in our country, which are important data in observing resistance development in bacteria. Beta-lactams are a group of antibiotics that inhibit cell wall synthesis at various stages. They constitute more than 50% of the antibiotics consumed worldwide [37]. In our study, the susceptibility rates of beta-lactam antibiotics in isolated E. coli strains were investigated. Resistance against ampicillin was detected at a rate of 30.8%. Many studies have shown that high levels of resistance to beta-lactam antibiotics, which are commonly used in clinical practice, are found in enteric bacteria isolated from various environments [38]. Resistance to this group of antibiotics is generally found in transferable resistance plasmids (R-plasmids), which leads to the rapid spread of resistance among the same or different species of microorganisms [39]. In a study conducted in our country, the blaTEM gene, encoding TEM-type beta-lactamase, was investigated in 96 ampicillin-resistant strains. As a result of the study, two strains were found to be positive for the blaTEM gene [40]. Beta-lactamase enzymes can be chromosomal or plasmid-mediated. Although plasmid-mediated beta-lactamases are commonly found in enteric bacteria, the most common type is reported to be TEM-1 among TEM-type beta-lactamases [41]. In a study by Yıldırım et al. (2018) [42], the TEM gene was investigated, including 44 samples. They identified the TEM gene in 33 (91.66%) of the 36 ESBL-producing E. coli isolates. The higher prevalence of this gene could be attributed to more frequent and over-the-counter use of antimicrobials in our country, as well as differences in antimicrobial prescribing practices and hygiene control measures. In the same study, the SHV gene region was also investigated but was not detected in any strain. While different rates of the TEM gene were found in studies conducted in our country, in our study, neither the TEM nor the SHV gene was detected in any of the 54 strains. Hemeg et al. (2018) [43] examined 120 E. coli isolates and detected the blaTEM ampicillin resistance gene in all of these isolates using PCR, highlighting the need to not only focus on patients in the classification of hospital and community-acquired infections. Van et al. (2008) [44] isolated 38 E. coli strains in their study conducted in Vietnam and used PCR analysis to detect the presence of some important antimicrobial resistance genes. They found the blaTEM gene in 84.2% of these E. coli isolates [44].
Tetracycline antibiotics are among the most widely used antibiotics in animal husbandry and agriculture, in addition to their use in the treatment of human infections. Tetracyclines constitute about 3% of the world's antibiotic production. It has been shown that members of the *Enterobacterales *family have at least 15 different genes that confer resistance to tetracyclines, and these genes may be plasmid, transposon, or chromosomal in origin [45]. In a study by Sevim et al. (2016) [40], 24 tetracycline-resistant isolates were examined for the presence of tetA, tetB, and tetC genes, and it was reported that the tetB gene was widespread among the isolates, with eight isolates containing the tetB gene and two containing the tetA gene. In our study, the tetA gene was detected in 47 (90.4%) of the isolated E. coli strains, and the tetB gene was detected in 50 (96.2%) of them. In a study conducted in our country using PCR with CTX-M universal primers, it was reported that all isolates (44 isolates) examined carried the CTX-M gene. Subsequently, in PCR using CTX-M group 1 primers, it was reported that all isolates carried CTX-M genes belonging to Group 1 [42]. Kürekci et al. (2019) [46] isolated 52 E. coli strains from various markets and butchers in Hatay and identified the CTX-M gene in 31 (62.3%) of them and the TEM gene in 19 (36.5%) of them. They reported that animal-derived foods significantly risk extraintestinal E. coli infections producing ESBLs. Nevertheless, they suggested that analyzing clinically ESBL-producing E. coli isolates together with those isolated from animal-derived foods would help better understand their potential source in Turkey. In our study, PCR was performed using CTX-M-F and CTX-M-R primers for E. coli and *Salmonella *spp. strains. The CTX-M gene region was not detected in any of the E. coli strains or *Salmonella *spp. strains. In a study by Dehdashti et al. (2019) [47], none of the 49 isolated E. coli isolates were found to carry CTX-M antimicrobial resistance genes similar to our study. Pehlivanlar Onen et al. (2015) [48] collected 100 chicken and 100 meat samples from various markets and butchers and isolated beta-lactamase-producing E. coli from 81 chicken samples and seven meat samples. They detected the blaCTX-M gene in 60 of the chicken isolates and 19 of the meat isolates, and the blaTEM gene in 19 of the chicken isolates and two of the meat isolates using PCR, reporting that retail-sold meat, especially chickens, was highly contaminated with ESBL-producing E. coli. Macrolides are used to treat infections because they are reliable and have good efficacy. Macrolide resistance can have significant consequences for public health. In a study by Phuc Nguyen et al. (2009) [49], the ereA gene was not detected in any of the 190 E. coli isolates. In a study by Dehkordi et al. (2014) [50], the ereA gene was detected in nine (18%) of the 50 isolated E. coli isolates. In a study conducted in our country by Keskin (2019) [51], the ereA gene was not detected. The ereA gene region was also not found in our study, which examined 54 strains.
The relatively small sample size included in this study limits the ability of the obtained data to represent the general population. Additionally, our analysis is based on data collected over a specific period. Therefore, in such cases, causal relationships cannot be fully determined. Studies that collect data repeatedly from the same populations over time can provide a better understanding of causal relationships. To this end, using larger and more diverse samples can increase the generalizability of the findings to a broader population.
Conclusions
In conclusion, according to the data obtained in our study, antimicrobial resistance genes that render antimicrobials used to treat diseases ineffective were detected in *E. coli *and Salmonella spp. isolates isolated from various farms and environmental samples investigated in our study. Therefore, studies should be conducted to prevent the emergence of new resistance genes in our country, as developing new drugs and treatment methods for these diseases is expensive and time-consuming. Since various diseases occur due to food contamination with *E. coli *and Salmonella spp., hygiene rules must be followed in the slaughter, storage, and transportation of these foods and on the farms where these foods are obtained. The results of this study have demonstrated that antimicrobial resistance genes are quite common in animal-derived foods consumed by humans using molecular methods and have shown that there is a potential reservoir for the transmission of antimicrobial resistance from animals to humans. This study will contribute to the measures to be taken for public health in our country and will provide a knowledge base for more advanced studies on this issue.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Colonization with antibiotic-resistant E. coli in commensal fecal flora of newborns Int J Curr Microbiol Appl Sci Tule A Hassani U 1623162962017 https://doi.org/10.20546/ijcmas.2017.605.177
- 2Antibiotics and bacterial resistance in the 21st century Perspect Medicin Chem Fair RJ Tor Y 2564620142523227810.4137/PMC.S 14459 PMC 4159373 · doi ↗ · pubmed ↗
- 3Antibiotic-resistant Escherichia coli and Salmonella spp. associated with dairy cattle and farm environment having public health significance Vet World Sobur MA Sabuj AA Sarker R Rahman AM Kabir SM Rahman MT 9849931220193152802210.14202/vetworld.2019.984-993PMC 6702575 · doi ↗ · pubmed ↗
- 4Antimicrobial resistance: a global multifaceted phenomenon Pathog Glob Health Prestinaci F Pezzotti P Pantosti A 30931810920152634325210.1179/2047773215 Y.0000000030 PMC 4768623 · doi ↗ · pubmed ↗
- 5Bacteria antibiotic resistance: New challenges and opportunities for implant-associated orthopedic infections J Orthop Res Li B Webster TJ 22323620182872223110.1002/jor.23656 PMC 5775060 · doi ↗ · pubmed ↗
- 6Benefits and risks of antimicrobial use in food-producing animals Front Microbiol Hao H Cheng G Iqbal Z 288520142497107910.3389/fmicb.2014.00288 PMC 4054498 · doi ↗ · pubmed ↗
- 7Antibiotics in agriculture and the risk to human health: how worried should we be?Evol Appl Chang Q Wang W Regev-Yochay G Lipsitch M Hanage WP 240247820152586138210.1111/eva.12185 PMC 4380918 · doi ↗ · pubmed ↗
- 8Escherichia coli in Europe: an overview Int J Environ Res Public Health Allocati N Masulli M Alexeyev MF Di Ilio C 623562541020132428785010.3390/ijerph 10126235 PMC 3881111 · doi ↗ · pubmed ↗