Mapping and Candidate Gene Analysis of an All-Stage Stem Rust Resistance Gene in Durum Wheat Landrace PI 94701
Hongyu Li, Kairong Li, Hongna Li, Chen Yang, Geetha Perera, Guiping Wang, Shikai Lyu, Lei Hua, Shams ur Rehman, Yazhou Zhang, Michael Ayliffe, Haitao Yu, Shisheng Chen

TL;DR
This study identifies and maps a stem rust resistance gene in durum wheat, offering a potential tool for improving wheat disease resistance.
Contribution
The study maps a novel all-stage stem rust resistance gene, SrPI94701, in durum wheat and identifies candidate genes.
Findings
SrPI94701 was mapped to chromosome arm 5BL and showed resistance to six Pgt races.
Two NLR genes were upregulated in resistant plants and showed high polymorphism between parental lines.
Flanking markers could not reliably predict SrPI94701 presence in a survey of 104 wheat accessions.
Abstract
Puccinia graminis f. sp. tritici (Pgt), the causal agent of wheat stem rust, poses a significant threat to global wheat production. Genetic resistance offers a cost-effective and sustainable solution. The durum wheat landrace PI 94701 was previously hypothesized to carry two stem rust resistance (Sr) genes, but their chromosomal locations were unknown. In this study, we mapped and characterized an all-stage Sr gene in PI 94701, temporarily designated as SrPI94701. In seedling tests, SrPI94701 was effective against all six Pgt races tested. Using a large segregating population, we mapped SrPI94701 on chromosome arm 5BL within a 0.17-cM region flanked by markers pku69124 and pku69228, corresponding to 1.04 and 2.15 Mb genomic regions in the Svevo and Chinese Spring reference genomes. Within the candidate region, eight genes exhibited differential expression between the Pgt-inoculated…
Genes, proteins, chemicals, diseases, species, mutations and cell lines named across the full text — each resolved to its canonical identifier and authoritative record.
Click any figure to enlarge with its caption.
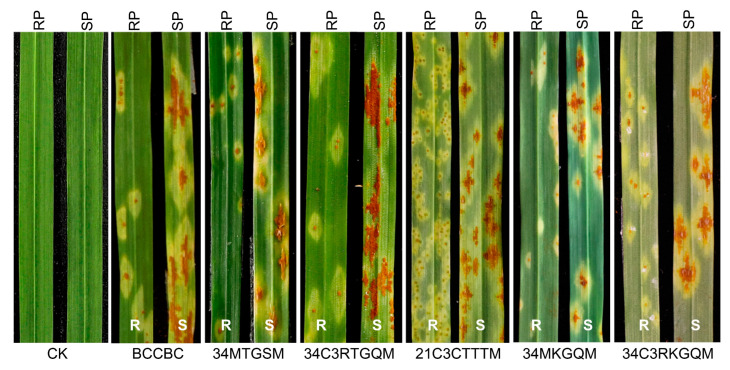 Figure 1
Figure 1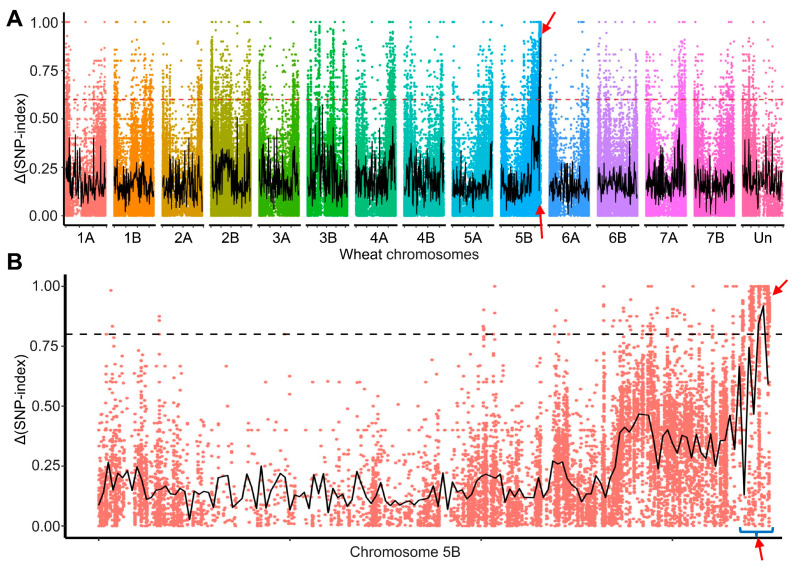 Figure 2
Figure 2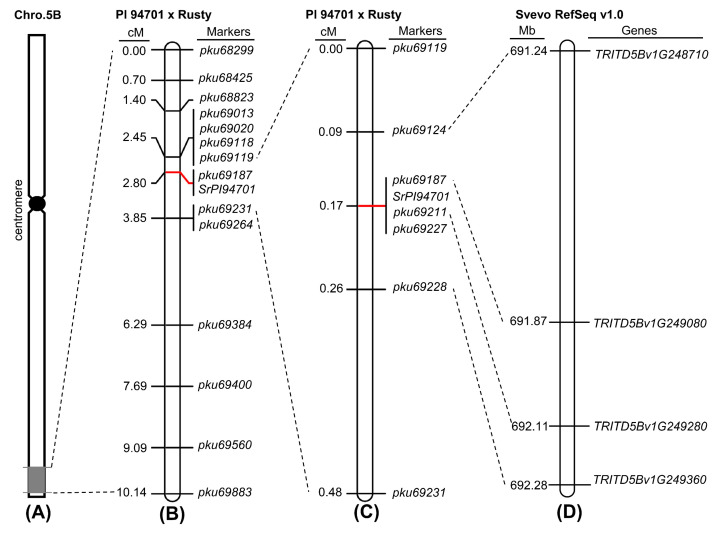 Figure 3
Figure 3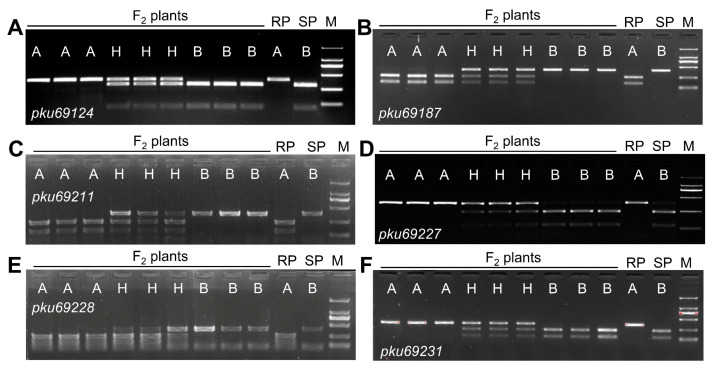 Figure 4
Figure 4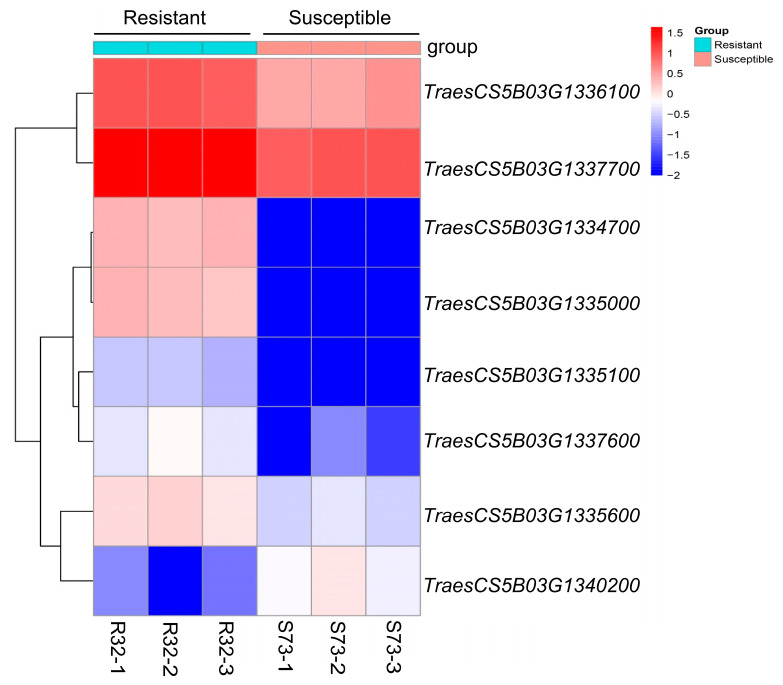 Figure 5
Figure 5- —Provincial Natural Science Foundation of Shandong
- —Key R&D Program of Shandong Province
- —National Key Research and Development Program of China
- —Young Taishan Scholars Program of Shandong Province
Peer Reviews
No public reviews on file for this paper yet. If you reviewed it on a platform where reviews are public (OpenReview, ICLR, NeurIPS, ICML), you can paste yours below so the community can read it here.
Videos
No videos yet. Explain this paper in a talk, walkthrough, or lecture? Add one.
Taxonomy
TopicsWheat and Barley Genetics and Pathology · Bioenergy crop production and management · Plant Disease Resistance and Genetics
1. Introduction
Wheat stem rust, a damaging fungal disease caused by Puccinia graminis f. sp. tritici (Pgt), has historically caused significant yield losses in wheat worldwide. In China, wheat stem rust currently occurs sporadically under natural conditions [1]. Historically, it has been a major problem in wheat production across Africa, Asia, Australia, Europe, and the Americas [2]. Losses are often severe, ranging from 50% to 90%, particularly when there is substantial disease pressure during grain filling. In some instances, individual fields can be destroyed in regions conducive to disease development, making it impossible to grow susceptible cultivars [3,4]. Growing resistant wheat cultivars stands as the most cost-effective and environmentally sustainable method for controlling this disease [5].
Highly virulent races of Pgt have emerged, overcoming numerous widely deployed resistance genes present in elite wheat cultivars globally. In 1998, a Pgt race, TTKSK (also known as Ug99), was identified in Uganda, exhibiting virulence against the widely deployed Sr genes, Sr31 and Sr38 [6]. Subsequent to the deployment of Sr24 in Kenya [7], further selection for virulence resulted in the identification of races TTKST, TTTSK (Sr36 virulence) [8], and more recently, TTKTT and TTKTK (SrTmp virulence) [9]. To date, a total of 15 variants within the Ug99 race group have been documented across at least 13 countries, highlighting the adaptability and widespread dissemination of Pgt [10,11]. Recently, Ug99 has been detected in Nepal [12], posing a threat to wheat production in South and Eastern Asia. In addition to the Ug99 lineage, several other highly virulent Pgt races have also been identified in stem rust outbreaks, including JRCQC, TKTTF, and TRTTF. The non-Ug99 race JRCQC was identified in Ethiopia [13]. This race exhibits combined virulence against Sr9e and Sr13b, which are commonly deployed in durum wheat [13,14]. Race TKTTF caused severe stem rust epidemics in Ethiopia during 2013–2014, affecting the widely grown Ug99-resistant wheat cultivar “Digalu” [15]. Subsequently, TKTTF was detected in at least ten countries [16,17]. Race TRTTF was observed in various countries, including Iran, Yemen, Ethiopia, and Pakistan [18,19]. This race defeated the resistance conferred by Sr genes Sr36, SrTmp, and Sr1RS^Amigo^, which are effective against the original Ug99 race TTKSK [13]. Additionally, novel Pgt pathotypes caused severe wheat stem rust epidemics and outbreaks in Central Asia [20]. This continuous evolution of the pathogen highlights the ongoing necessity to explore new effective Pgt resistance genes for wheat breeding programs.
To date, more than 60 stem rust resistance genes have been formally cataloged in bread and durum wheats, as well as their wild relatives [21,22]. Among these, tetraploid wheat (Triticum turgidum ssp.) has contributed multiple Sr genes (or alleles), such as Sr2, Sr9d/Sr9e/Sr9g, Sr11, Sr12, Sr13a/Sr13b/Sr13c/Sr13d, Sr14, and Sr17 [4,14,23,24,25]. All of these genes from tetraploid wheat are all-stage resistance (ASR) genes, with the exception of Sr2, which is an adult plant resistance (APR) gene [26]. ASR genes typically confer a high level of resistance at all stages of plant growth and development, but they are often specific to certain pathogen races. In contrast, APR genes tend to provide only partial resistance at later stages of plant development, and a single APR gene is often inadequate for achieving satisfactory levels of plant protection [27]. Due to the large and complex nature of the polyploid wheat genome, only two ASR Sr genes (Sr9 and Sr13) have been successfully cloned so far in durum wheat [14,25]. As durum and bread wheat share common A and B genomes, it is relatively easy to transfer important Sr genes from T. durum into bread wheat, allowing these genes to be utilized in bread wheat breeding programs.
The durum wheat landrace PI 94701, collected in Palestine, was previously postulated to carry two temporarily designated Sr genes (Srdp1 and Srdp2) based on the observed segregation of resistance against Pgt race 11-SS2 [28]. Plants carrying only Srdp1 displayed resistant infection types (ITs = 0; to 1+), and plants carrying only Srdp2 showed moderate resistance (ITs = 1+ to 2−) [28]. Initially, Srdp2 was proposed to be allelic to Sr13, a gene located on chromosome arm 6AL. However, subsequent studies revealed that Srdp2 is distinct from Sr13 based on pathogenic variability investigations and sequencing of polymerase chain reaction (PCR) products of diagnostic markers for Sr13 [14]. Despite these findings, the chromosomal locations of Srdp1 and Srdp2 remain unknown. The primary objectives of the present study were to: (1) assess whether PI 94701 confers resistance to Chinese Pgt races; (2) genetically map the stem rust resistance genes present in PI 94701; and (3) identify the corresponding regions within sequenced wheat genomes and predict potential candidate genes.
2. Results
2.1. Characterization of Stem Rust Resistance in Durum Wheat Accession PI 94701
Durum wheat accessions PI 94701 and Rusty were challenged with six Pgt races, comprising five from China and one from North America (Table S1). PI 94701 displayed high resistance [infection types (ITs) = 1 or 1+] to all tested Pgt races, whereas Rusty exhibited susceptible infection types ranging from 3+ to 4 (Figure 1). The F_1_ plants resulting from the cross between PI 94701 and Rusty exhibited resistance against Pgt race 34MKGQM. A subset of 143 F_2_ plants derived from this cross was evaluated for their reactions to Pgt race 34MKGQM. Within this subset, 105 plants showed resistance (ITs = 1 to 2−), while 38 plants were susceptible (ITs = 3 to 4). Chi-squared analysis of the phenotyping data revealed a segregation ratio of 3:1 consistent with a single dominant gene (χ^2^ = 0.19, p = 0.66), which is henceforth referred to as SrPI94701. Among the 143 F_2:3_ families evaluated with Pgt race 34MKGQM, 32 were homozygous resistant, 73 were segregating, and 38 were homozygous susceptible. This distribution corresponded to the expected 1:2:1 segregation ratio (χ^2^ = 0.57, p = 0.75).
Based on their resistance or susceptibility to Pgt race 34MKGQM, ten homozygous resistant and ten homozygous susceptible F_2:3_ families were selected from the PI 94701 × Rusty cross. These families were further tested against five additional Pgt races: BCCBC, 34MTGSM, 21C3CTTTM, 34C3RTGQM, and 34C3RKGQM, and were grown in five separate growth chambers. The ten families exhibiting resistance to 34MKGQM also demonstrated robust resistance to other Pgt races, while the ten 34MKGQM-susceptible families showed consistent susceptibility (Figure S1). These results suggest that the resistance of PI 94701 to races BCCBC, 34MTGSM, 21C3CTTTM, 34C3RTGQM, and 34C3RKGQM is linked to the SrPI94701 genomic region.
2.2. Genetic Mapping of SrPI94701 on Chromosome Arm 5BL
For the initial mapping, bulked segregant RNA-Seq (BSR-seq) was performed on the F_2:3_ mapping population evaluated with Pgt race 34MKGQM, revealing 32,708 single nucleotide polymorphisms (SNPs) between PI 94701 and Rusty. Based on SNP index analysis, 160 SNPs within the genomic region spanning from 673.5 to 699.9 Mb (Svevo Rel.1.0) on the long arm of chromosome 5B exhibited significant association with the phenotype (Figure 2, Table S2). This result indicates that SrPI94701 was located on chromosome arm 5BL.
To verify the mapping location, we specifically selected SNPs surrounding the region of interest on chromosome arm 5BL and developed 14 5B-genome-specific Cleaved Amplified Polymorphic Sequence (CAPS) markers (Table 1). Subsequently, these markers were used to genotype the 143 F_2:3_ families, resulting in the construction of a genetic linkage map for SrPI94701 (Figure 3A,B). Based on the linkage results, SrPI94701 was found to be completely linked to marker pku69187 and mapped within a 1.4 cM interval flanked by markers pku69119 and pku69231 (Figure 3B).
To precisely determine the position of SrPI94701, an additional 1008 F_4_ plants from six selected segregating F_3_ families were screened for recombinants between markers pku69119 and pku69231. This screening identified ten plants carrying recombination events between these two markers. Using these ten recombinants detected in this screening and one previously identified recombinant in 143 F_3_ families, the genetic distance between markers pku69119 and pku69231 was recalculated to be 0.48 cM (Figure 3C). We evaluated the progeny of these plants carrying informative recombination events with Pgt race 34MKGQM. Using these recombinants and four new PCR markers developed within this genomic region (Table 1), SrPI94701 was further mapped to a 0.17-cM interval flanked by CAPS markers pku69124 and pku69228, and was completely linked to markers pku69187, pku69211, and pku69227 (Figure 3C). PCR amplifications with six CAPS markers are showcased in Figure 4.
2.3. Candidate Genes for SrPI94701 within Tetraploid and Hexaploid Wheat Genomes
The 0.17-cM candidate region delimited by PCR markers pku69124 and pku69228 spans a 1.04 Mb genomic region in the T. durum reference genome of Svevo (691.24–692.28 Mb, Figure 3D) and a 2.15 Mb genomic region in the T. aestivum reference genome of Chinese Spring (CS; 702.56–704.71 Mb). These candidate regions contain 65 annotated genes in Svevo (TRITD5Bv1G248710–TRITD5Bv1G249360; Table S3) and 28 high-confidence genes in CS (TraesCS5B03G1334300–TraesCS5B03G1340200; Table S4). These annotated genes include one typical NLR gene in Svevo and seven in CS, which is particularly noteworthy for this study as NLRs are the predominant class of genes associated with disease resistance in plants [25,32,33,34]. In addition to these NLR genes, we identified one protein kinase in Svevo and six in CS (Tables S3 and S4) as potential candidates due to their known roles in disease resistance across various plant species [35,36,37,38,39].
Subsequently, we analyzed the transcript levels of the candidate genes using published RNAseq studies compiled in the wheat ExpVIP database (http://www.wheat-expression.com/; accessed on 8 June 2024). Of the 28 high-confidence genes annotated in the candidate gene region in the CS genome, only 16 high-confidence genes were found to be expressed in wheat leaves (Table S4).
2.4. Identification of Differentially Expressed Genes (DEGs) Within the SrPI94701 Mapping Interval
Transcript levels of the candidate genes were analyzed using RNA-seq data from the F_4_ sister lines S73 and R32, selected from the PI 94701 × Rusty cross. Given the identification of numerous NLR and kinase genes in the colinear regions of CS (Table S4), we focused on analyzing the 28 annotated genes within the candidate interval in CS. Among these candidate genes, eight exhibited differential expression between the Pgt-inoculated resistant and susceptible plants (Figure 5). Seven DEGs showed significant upregulation in R32 plants compared to S73 plants, whereas only one DEG displayed downregulation (Table S5). These upregulated DEGs included four NLR genes: TraesCS5B03G1334700, TraesCS5B03G1335000, TraesCS5B03G1335100, and TraesCS5B03G1335600, making them strong candidate genes.
We used whole-genome resequencing reads from PI 94701 and Rusty to assemble genomic contigs containing the identified NLR DEGs. Upon comparison of these contigs between PI 94701 and Rusty, we observed that these NLRs were either missing or partially deleted in Rusty (Figure S2). Among them, we excluded TraesCS5B03G1335000 and TraesCS5B03G1335600 as candidate genes due to the identical nature (100% match) of their coding and promoter regions between PI 94701 and CS. However, CS exhibited susceptibility to Pgt races 34MTGSM, 34C3RTGQM, 21C3CTTTM, 34MKGQM, and 34C3RKGQM at the seedling stage, suggesting the absence of SrPI94701.
To validate the RNA-seq findings, we measured the expression levels of the candidate genes TraesCS5B03G1334700 and TraesCS5B03G1335100 relative to ACTIN using quantitative real-time PCR (qRT-PCR). The results confirmed a significant upregulation (p < 0.05) of these candidate genes between the two groups (Figure S3), supporting the conclusions from the RNA-seq analysis.
2.5. Validation of SrPI94701-Linked Markers in Uncharacterized Wheat Accessions
To assess the significance of the haplotype defined by the two flanking and three completely linked markers (Figure 3C and Figure 4), we used these markers to evaluate 53 T. dicoccon and 51 T. aestivum accessions. Among the 53 T. dicoccon genotypes, 29 accessions (54.7%) exhibited an identical haplotype to the resistant parent, PI 94701 (Table S6). Nevertheless, certain accessions with the SrPI94701 haplotype, like PI 273980 and PI 197489, were susceptible to Pgt race 34MKGQM (Table S6), indicating that these markers could not accurately predict the presence of SrPI94701 in uncharacterized T. dicoccon genotypes.
Among the 51 tested hexaploid wheat accessions, the SrPI94701 haplotype was detected in 8 accessions (15.7%; Table S6). Similarly, two accessions carrying the SrPI94701 haplotype (Huaimai40 and Luyuan502) showed susceptibility to Pgt race 34MKGQM, suggesting that these markers were ineffective in predicting the presence of SrPI94701 in hexaploid wheat.
3. Discussion
Initially, the durum wheat landrace PI 94701 was reported to carry a single Sr gene conferring resistance to Pgt race 15B [40]. Later, it was proposed that PI 94701 carries two provisionally designated Sr genes (Srdp1 and Srdp2) based on the segregation of resistance against Pgt race 11-SS2 [28]. However, the precise chromosomal locations of Srdp1 and Srdp2 remained unknown. In this study, using Chinese Pgt races, we successfully mapped the stem rust resistance gene SrPI94701 within a 0.17 cM region on the long arm of chromosome 5B. However, the lack of chromosomal locations and genetic maps for Srdp1 and Srdp2 has limited our ability to establish the mapping relationship between SrPI94701 and Srdp1/Srdp2.
The clustering of NLR genes promotes the generation of new variants through duplication, recombination, and conversion events, thereby increasing the diversity of NLR genes [14,41]. Using the reference genomes of tetraploid and hexaploid wheat, we defined the SrPI94701 candidate region as a 1.04 Mb region in T. durum wheat Svevo and a 2.15 Mb region in hexaploid wheat CS, which includes a cluster of typical NLR genes. However, NLR genes identified within the colinear regions of the Svevo and CS reference genomes display copy number variations (Tables S3 and S4). Similar to the SrPI94701 candidate region, reports of copy number and structural variations of NLR genes have been documented for previously isolated disease resistance genes in wheat and its wild relatives [14,25,33,41,42]. Considering that many cloned R genes in plants encode intracellular NLR proteins [32,34], these NLR genes within the mapped region are strong candidates for SrPI94701.
In addition to the NLR genes, several protein kinases were also identified in the SrPI94701 mapping region (Tables S3 and S4). This gene family is frequently linked to disease resistance in various plant species [43,44]. In wheat, six cloned rust resistance genes, Sr60 [35], Sr62 [45], Sr43 [46], Yr36 [36], Yr15 [39], and Lr9/Lr58 [47], encode proteins with kinase domains. These NLRs and protein kinases have been prioritized for functional characterization to explore their potential as the causal gene for SrPI94701. However, we cannot rule out the possibility of additional NLRs or protein kinases present in PI 94701 that are absent in the reference genomes.
The wheat stem rust resistance genes, Sr49 and Sr56, were previously mapped close to SrPI94701 on the long arm of chromosome 5B [48,49]. Sr56, known for conferring adult plant resistance, was mapped in the Swiss winter wheat cultivar Arina and demonstrated a 12–15% reduction in stem rust severity under field conditions [49]. Sr56 was flanked by SSR markers sun209 and sun215 [49], corresponding to a genomic region ranging from 705,196,548 bp to 705,764,022 bp in the CS RefSeq v2.1 reference genome. These results suggest that SrPI94701 is distinct from the Sr56 gene. The ASR gene Sr49 was mapped in the hexaploid wheat landrace AUS28011 and showed effectiveness against all commercially important Australian Pgt races that were examined, displaying mesothetic resistant (ITs = 2−) infection types [48]. The location of Sr49 was delimited by SSR markers sun479 and sun209, spanning a genetic region from 702,174,220 bp to 705,196,462 bp (CS RefSeq v2.1) which includes our candidate gene region (702.56–704.71 Mb, Table S4). However, there are three differences between SrPI94701 and Sr49. First, SrPI94701 was discovered in durum (tetraploid) wheat, while Sr49 was identified in hexaploid wheat. Second, wheat plants carrying SrPI94701 (ITs = 1 or 1+) showed better resistance compared to those with Sr49 (ITs = 2−). Third, AUS28011 (Sr49) and PI 94701 (SrPI94701) showed different haplotypes when genotyped with the flanking and completely linked markers of SrPI94701. Nonetheless, due to their co-location within overlapping genetic intervals, SrPI94701 and Sr49 may be the same gene.
Previously, no haplotype containing both flanking marker alleles sun479 and sun209 was identified in the screening of 152 Australian and Nordic wheat lines. Nevertheless, six wheat lines carried the Sr49-linked sun479_200bp_ allele, and seven harbored the Sr49-linked sun209_148bp_ allele [48]. The Sr49 locus was also detected in a Genome-Wide Association Study (GWAS) involving a panel of 283 durum wheat lines challenged with various East African stem rust isolates [50]. These two markers (sun479 and sun209) were detected with a minor allele frequency (average of 0.053), indicating the rarity of Sr49 within this germplasm panel [50]. In this study, the genotypes of the flanking and completely linked markers of SrPI94701 revealed that 54.7% of T. dicoccon and 15.7% of T. aestivum accessions showed the SrPI94701-linked resistant haplotype. The notable differences in allele frequencies observed could be attributed to the diversity among the germplasm sources. Alternatively, it is also possible that they represent distinct genes, which explains the obvious differences in allele frequencies detected.
SrPI94701 confers resistance against multiple Pgt races in China, making it a potentially valuable genetic resource in wheat breeding. However, as SrPI94701 provides only an intermediate level of resistance when used alone, it needs to be combined with other Sr genes to provide economically useful levels of resistance. Some potentially valuable Sr genes that could enhance resistance include Sr35 [51], Sr13 [14], Sr22a/Sr22b [52,53], and Sr26 [54]. Despite this limitation, SrPI94701 can provide an additional layer of resistance against various Pgt races and contribute to diversifying the sources of resistance used in wheat breeding programs.
4. Materials and Methods
4.1. Plant Materials and Mapping Populations
The Pgt-resistant Triticum turgidum ssp. durum (2n = 4x = 28, AABB) wheat accession PI 94701 was crossed with the highly susceptible durum wheat genotype Rusty [14,55]. For the initial mapping, we evaluated a population of 143 F_2_ plants derived from the PI 94701 × Rusty cross inoculated with Chinese Pgt race 34MKGQM (isolate 20IAL06). To construct a high-density genetic map, six F_2_ plants (plants 15R, 37R, 75R, 90R, 118R, and 126R) heterozygous for the SrPI94701 region, identified using flanking molecular markers, were selected. These plants generated a secondary population consisting of 1008 F_3_ individuals. These F_3_ plants were genotyped with SrPI94701 flanking markers to identify plants with recombination events within the candidate gene region. The recombinants and their corresponding F_4_ progeny (~25 plants per family) were challenged with Pgt race 34MKGQM. As a source of Sr49, we used the bread wheat landrace AUS28011 (Mahmoudi) from Tunisia, which exhibited good resistance to five Australian Pgt pathotypes [48]. Finally, we used a collection of 53 accessions of T. turgidum ssp. dicoccon and 51 accessions of T. aestivum to determine the value of the SrPI94701-linked markers developed in the present study for marker-assisted selection.
4.2. Evaluation for Stem Rust Resistance
Stem rust seedling assays for both the parental lines and the segregating populations were conducted at the Peking University Institute of Advanced Agricultural Sciences following previously described procedures [56,57]. Six Chinese and North American Pgt races, namely BCCBC, 34MTGSM, 34C3RTGQM, 21C3CTTTM, 34MKGQM, and 34C3RKGQM [58], were evaluated for virulence against parental lines PI 94701 and Rusty. The avirulence/virulence profiles of these Pgt races are detailed in Supplementary Table S1. Pgt urediniospores, stored at − 80 °C, were subjected to heat shock at 42 °C for 3 min. Subsequently, a urediniospore/talc suspension was applied to seedlings at the three-leaf stage using the shaking-off method [29]. Following inoculation, the plants were kept in a dark dew chamber overnight and then transferred back to growth chamber conditions maintained at 22–24 °C with a 16 h photoperiod. Pgt infection types (ITs) were recorded at 12–14 dpi using the Stakman 0–4 scale, indicating levels of immunity to susceptibility [59]. For plants carrying recombination events within the candidate region, progeny tests were conducted using approximately 25 plants from each F_2:3_ family.
4.3. Bulked Segregant RNA-Seq (BSR-Seq) Analysis
Based on the phenotypic data against Pgt race 34MKGQM, 15 homozygous resistant and 15 homozygous susceptible F_3_ families were selected to constitute the resistant bulk (R-bulk) and susceptible bulk (S-bulk) for RNA extraction. Leaves of equal length were taken from the second leaf of all seedlings in each family. After harvest, each sample was immediately flash-frozen in liquid nitrogen and stored at −80 °C for RNA extraction. Total RNAs were extracted from the resistant parent PI 94701 and R/S-bulks using the Spectrum^TM^ Plant Total RNA Kit (MilliporeSigma, Burlington, MA, USA). RNA sequencing was conducted at Novogene Bioinformatics Technology Co., Ltd. (Beijing, China) with 150-base paired-end reads. The raw sequencing data are available at the National Genomics Data Center (NGDC) under the BioProject accession number PRJCA027291. For the susceptible parent Rusty, we used the whole-genome resequencing data described in Wang et al. (2023a) under the BioProject accession number PRJCA017761.
Raw RNA-seq reads were trimmed using Trimmomatic v. 0.32 [60] to remove low-quality nucleotide sequences. The trimmed reads were then aligned to the reference genome of the durum wheat cultivar Svevo [61] using STAR v2.5.0c [62]. SNPs were identified using the HaplotypeCaller tool from GATK v3.2-2 [63]. SNP-index and Δ(SNP-index) were calculated across the whole genome to identify potential regions of interest using the methods described previously [30,64,65].
4.4. Development of PCR Markers
To accelerate the development of PCR markers, we performed whole-genome resequencing for the resistant parent PI 94701 (accession number PRJCA027291). PCR primers were designed using the Primer3 software (https://bioinfo.ut.ee/primer3-0.4.0/primer3/; accessed on 8 June 2024). The identified SNPs within the mapping interval were used to develop CAPS markers [66]. Enzyme cleavage reactions of the CAPS markers were performed according to standard procedures for the corresponding restriction endonucleases (New England Biolabs, Hitchin, UK). PCR reactions were conducted using a Veriti 96-Well Fast Thermal Cycler (Thermo Fisher Scientific Inc., Waltham, MA, USA). After the PCR amplification, 10 µL of PCR products were digested with the suitable restriction enzyme. Subsequently, 5 µL of digested PCR products were analyzed using agarose gel electrophoresis, and the gels were stained with ethidium bromide.
4.5. qRT-PCR Analysis
From the PI 94701 × Rusty population, a pair of F_4_ sister lines homozygous for the absence (S73) or presence (R32) of SrPI94701 were developed using PCR markers. The susceptibility/resistance responses for S73 and R32 against Pgt race 34MKGQM were evaluated. Leaves from S73 and R32 inoculated with race 34MKGQM were sampled at 6 dpi, with three biological replicates used for each genotype. RNA sequencing was conducted at Novogene Bioinformatics Technology Co., Ltd. (Beijing, China). The raw sequencing data are available under the same BioProject accession number, PRJCA027291. DEGs between the sister lines S73 and R32 were determined using the edgeR software v. 4.2.1, applying significance thresholds of FDR < 0.05, p-value < 0.05, and |log2foldchange| > 1 [67]. DEGs within the SrPI94701 candidate region were identified, and a heatmap was generated using the pheatmap R package (https://cran.ms.unimelb.edu.au/web/packages/pheatmap/pheatmap.pdf; accessed on 8 June 2024). qRT-PCR reactions were performed using an ABI QuantStudio 5 Real-Time PCR System (Applied Biosystems, Foster City, CA, USA). Transcript levels were determined in four biological replicates and quantified as fold-ACTIN levels using the 2^−ΔCT^ method [33,41].
4.6. Statistical Analyses
Genetic linkage maps with polymorphic markers and stem rust resistance scores of F_2:3_ families were constructed using the MapChart v2.2 software (https://www.wur.nl/en/show/Mapchart.htm; accessed on 8 June 2024) [31]. The significance of the differences in transcript levels was evaluated using a two-sided, unpaired t-test.
5. Conclusions
In this study, we mapped and characterized SrPI94701, an all-stage stem rust resistance gene in durum wheat landrace PI 94701. The broad efficacy of SrPI94701 in China makes it a desirable target for introgression into Chinese bread wheat cultivars. For recurrent wheat cultivars with different marker haplotypes, the flanking and fully linked markers for SrPI94701 can be used to transfer this gene from durum into modern bread wheat varieties. While SrPI94701 confers only partial resistance to Pgt races, this might be beneficial in breeding programs aiming to combine multiple partial resistance genes and avoid major all-stage resistance genes, a strategy that has been proposed to increase the durability of wheat resistance to stem rust [68].
Using a large mapping population consisting of 2302 recombinant gametes, SrPI94701 was successfully mapped to the telomeric region of chromosome arm 5BL within a 0.17-cM region. The presence of NLR genes in the collinear regions of wheat reference genomes indicates that SrPI94701 may be a typical NLR gene. However, final identification of the SrPI94701 gene will be required to fully test this hypothesis and determine the relationship between SrPI94701 and Sr49. To ascertain if the two identified NLR candidate genes are required for resistance to Pgt, we are currently conducting functional characterization using truncation mutations for each gene.
In summary, the high-resolution genetic map of SrPI94701 and the closely linked PCR markers developed in this study will facilitate map-based cloning of this Sr gene and accelerate its incorporation into modern wheat breeding programs.
The reference list from the paper itself. Each links out to its DOI / PubMed record.
- 1Cao Y. Yao P. Wu Y. Bi Y. Yang J. Discovery and verification of an important inoculum source for Puccinia graminis f. sp. tritici in China Plant Prot.200128294298
- 2Saari E.E. Prescott J. World distribution in relation to economic losses Diseases, Distribution, Epidemiology, and Control Elsevier Amsterdam, The Netherlands 1985259298
- 3Amulaka F. Maling’a J. Pathak R. Cakir M. Mulwa R. Yield evaluation of a wheat line with combined resistance to Russian wheat aphid and stem rust race “Ug 99” in Kenya Am. J. Plant Sci.201341494149910.4236/ajps.2013.47181 · doi ↗
- 4Singh R.P. Hodson D.P. Huerta-Espino J. Jin Y. Bhavani S. Njau P. Herrera-Foessel S. Singh P.K. Singh S. Govindan V. The emergence of Ug 99 races of the stem rust fungus is a threat to world wheat production Annu. Rev. Phytopathol.20114946548110.1146/annurev-phyto-072910-09542321568701 · doi ↗ · pubmed ↗
- 5Singh R.P. Hodson D.P. Jin Y. Lagudah E.S. Ayliffe M.A. Bhavani S. Rouse M.N. Pretorius Z.A. Szabo L.J. Huerta-Espino J. Emergence and spread of new races of wheat stem rust fungus: Continued threat to food security and prospects of genetic control Phytopathology 201510587288410.1094/PHYTO-01-15-0030-FI 26120730 · doi ↗ · pubmed ↗
- 6Pretorius Z.A. Singh R.P. Wagoire W.W. Payne T.S. Detection of virulence to wheat stem rust resistance gene Sr 31 in Puccinia graminis f. sp. tritici in Uganda Plant Dis.20008420310.1094/PDIS.2000.84.2.203B 30841334 · doi ↗ · pubmed ↗
- 7Jin Y. Szabo L.J. Pretorius Z.A. Singh R.P. Ward R. Fetch T. Detection of virulence to resistance gene Sr 24 within race TTKS of Puccinia graminis f. sp tritici Plant Dis.20089292392610.1094/PDIS-92-6-092330769714 · doi ↗ · pubmed ↗
- 8Jin Y. Szabo L.J. Rouse M.N. Fetch T. Pretorius Z.A. Wanyera R. Njau P. Detection of virulence to resistance gene Sr 36 within the TTKS race lineage of Puccinia graminis f. sp tritici Plant Dis.20099336737010.1094/PDIS-93-4-036730764215 · doi ↗ · pubmed ↗